Snake Envenomation: A Component- Based Review on Hemostatic Alterations
Considered by the World Health Organization a neglected public health problem, snakebites occur predominantly in tropical areas of Africa, Asia and Latin America. Approximately 2.7 million people worldwide are victims of snakebites each year, of which between 81,000 and 138,000 later die. Among the survivors, about 400,000 have some permanent disability. The entirety of the mechanism responsible for the venom pathophysiology is not completely understood. However, most of the venom toxins affect human hemostasis, as Bothrops venom components, which destabilize endothelium, affect platelet aggregation, blood clotting and fibrinolysis. Such effects are not only components of the clinical picture of patients who have suffered envenomation, but are also directly associated with the severity of the accident. Thus, it is of great importance to understand the main hemostatic alterations observed in envenomation’s by snakes of the genus Bothrops, which is predominant in South America. Therefore, this review describes the hemostatic changes that occur in Bothrops snakebites, to further improve the understanding of the general pathological mechanisms of snake envenomation’s and the correlation between endothelium dysfunction and coagulation/fibrinolysis systems as a result of the interaction of each class of venom components with human hemostasis. This knowledge is crucial for the development of new effective therapeutic approaches, attenuating the severity of snakebites and reducing amputations and deaths. Besides this, a molecular comprehension of the hemostatic alterations caused by each component of Bothrops venoms may help to identify new molecules and targets for therapeutic applications.
Introduction
Approximately 2.7 million people worldwide are victims of snakebites each year, of which between 81,000 and 138,000 later die. Among the survivors, about 400,000 have some permanent disability [1]. Considered by the World Health Organization (WHO) a neglected public health problem, snakebites occur more frequently in populations that have poor access to health services, as is the case of most populations living in rural environments and on the urban outskirts [2, 3, 4]. Brazil, among other countries, has a commitment with the WHO to reduce the damage caused by snakebites by 50% by 2030 [5].
Snakebite accidents occur predominantly in tropical areas of Africa, Asia and Latin America. In the latter, snakes of the genus Bothrops are the main causes of snakebites, with Brazil being the country with the highest number of cases on the continent, with 28,000 yearly snakebites and 116 deaths resulting from them, mainly caused by Bothrops jararaca. One of the main aspects that lead to the high incidence of snakebites in Brazil is the wide territorial distribution of snakes of the Bothrops genus in the country and the large number of farm workers [6].
Bothrops venom composition may vary interspecific and intra specifically according to age, habitat and ecological niche of the individuals [1, 7, 8]. In general, Bothrops venoms comprise some protein families including the snake venom metalloproteases (SVMPs); snake venom serine proteases (SVSPs); phospholipases A2; type C lectins; bradykinin potentiators; L-amino acid oxidases; glutaminyl-peptide cyclotransferases; hyaluronidases and venom endothelial growth factors [9, 10].
Most of these toxins affect hemostasis, that is, the set of mechanisms that prevent blood from leaking out of the vessels. Bothrops venom toxins can destabilize endothelium, and both induce or inhibit platelet aggregation, blood clotting, and fibrinolysis. Such effects are not only components of the clinical picture of patients who have suffered envenomation, but are also directly associated with the severity of the accident. Thus, it is of great importance to understand the main hemostatic alterations observed in envenomation’s by snakes of the genus Bothrops.
SVSPs
Snake venom serine proteases (SVSPs) are toxins that affect mainly the hemostatic system. They are proteolytic enzymes that hydrolyze specific peptidic bonds and can be classified as endopeptidases or exopeptidases, depending on the cleavage site [11, 12]. The active site structure of this family of enzymes is formed by the catalytic triad, serine, histidine and aspartic acid, and their boundaries along the chain (His57, Asp102 and Ser195), where the stability is granted by hydrogen bounds [13, 14]. SVSPs are analogous to mammalian trypsin, belong to the S1 family of chymotrypsin of the SA clan, have in their structure, in addition to the catalytic triad, 12 cysteine residues (Cys) that form disulfide bonds, and have cleavage specificity after residues of arginine or lysine. They can act on several components of the coagulation cascade, the fibrinolytic system and the kallikrein-kinin system. Some SVSPs also act on the complement system [15]. Bothrops SVSPs are majorly thrombin-like enzymes. These enzymes are similar to human thrombin, but cleave fibrinogen without leading to the formation of stable fibrin network, by not activating FXIII, which is essential for clot formation (Figure 1). In addition, thrombin-like enzymes usually do not activate coagulation FXIII, which is responsible for forming bonds that provide greater stability to the fibrin network. Procoagulant SVSPs are enzymes capable of activating factors II, VII and X of the blood clotting cascade, decreasing clotting time. Anticoagulant SVSPs activate protein C by cleavage of peptide bonds by a thrombomodulin-independent mechanism [16]. The fibrinolytic action of SVSPs is converting plasminogen to plasmin, which quickly degrades preexisting clots [17].
Batroxobin is a serine protease isolated from B. moojeni that has been used in clinical medicine as defibrinogenating agent. Batroxobin, unlike thrombin, has fibrinogen as its sole substrate and only releases fibrinopeptide A without cleaving fibrinopeptide B. Also, it is not inhibited by and thrombin or heparin cofactor II [18]. These properties allow Batroxobin to be used as a tool for abnormal coagulation detection. Batroxobin has also been used for prevention and treatment of thrombosis, because it lowers circulating fibrinogen levels [19, 20]. Botroxobin also promotes fibrin accretion greater than thrombin, which may explain the thrombosis in B. moojeni envenomation [21].
Thrombocytin and PA-BJ are two SVSPs purified from B. atrox and B. jararaca respectively. Both enzymes are capable of induce platelet aggregation through the G-protein coupled receptors (GPCR), PAR1 and PAR4. PA-BJ acts through the inactivation of the tethered ligand and induces calcium mobilization in fibroblasts transfected with PAR4, desensitizing these cells to the action of thrombin [22]. Similarly to thrombin, Thrombocytin activated factor XIII by limited proteolysis and increased the procoagulant activity of factor VIII [23].
Zaqueo KD, et al. [24, 25] characterized two SVSPs, BbrzSP-32 and BpirSp-39. BbrzSP-32 is the first SVSP isolated from B. brazili, it presents thrombin-like activity and is able to degrade the α-chain of fibrinogen in in vitro models. BbrzSP-32 demonstrated proteolytic activity on gelatin and on substrates for serine proteases and thrombin like enzymes, besides having coagulant activity on plasma [24]. The SVSP BpirSp-39 is purified from the venom of B. pirajai. This enzyme was able to form fibrin networks and also promote platelet aggregation, displaying fibrinogenolytic, fibrinolytic, gelatinolytic and amidolytic activities. BpirSP-39 is apparently able to activate the clotting cascade factor XIII and act on substrates for plasma kallikrein, factor XIa and XIIa, factor Xa and for thrombin like enzymes [25].
There are other SVSPs that have fibrinogenolytic activity, such as Bjussu SP-I and Bothrops protease A (BPA). Bjussu SP-I is purified from the venom of B. jararacussu and shows high proteolytic activity on serine protease and thrombin chromogenic substrates. It also shows procoagulant and kallikrein-like activity [26]. BPA is a potent fibrinogenolytic enzyme purified from B. jararaca venom, but it does not form fibrin, rather, it prevents the formation of fibrin by degrading fibrinogen a- and b-chains [27].
In general, SVSPs act mainly on the coagulation cascade leading to hemostatic imbalance, but their action is not limited to the coagulation, as they promote specific proteolysis on key points of different systems [12]. It has also been demonstrated that some serine proteases have proangiogenic properties [28], raising questions about the importance of the role of these enzymes on the local and systemic effects on snakebite accidents.
SVMP
Snake venom metalloproteinases (SVMPs) are zinc- dependent endoproteases (Zn2+) that has a consensus (HExxHxxGxxHD) sequence and a Met-turn. They tend to be the most abundant toxins in the venom of Bothrops snakes, especially in those species exposed to low annual temperature ranges, where SVMPs often account for more than 30% of the chemical composition of the venom [29]. SVMPs structurally resemble proteins present in mammals, ADAM (disintegrin and metalloproteinase) and ADAMTS (ADAM with thrombospondin motif type 1) [30]. Traditionally, SVMPs are classified into three classes according to the presence of certain domains. Class P-I is the simplest and has the lowest molecular weight, with only the M domain (metalloproteinase contains the catalytic site), class P-II has the M and D (disintegrin) domains and class P-III has the domains M, D and C (rich in cysteine) [31, 32].
The hemorrhagic effect of envenomation is largely caused by SVMPs, mainly those belonging to classes P-II and P-III. Bleeding is due, at least in part, to the loss of integrity of the endothelium, especially in the microvasculature. Although SVMPs can have a cytotoxic effect on endothelial cells, such effects appear to have little causal relationship to hemorrhage [33, 34]. Furthermore, in B. jararaca envenomation, the increase in thrombomodulin levels, the main marker for endothelium injuries, is not correlated with the activity of SVMPs, but mainly with the mass activation of thrombin during envenomation. Thus, a direct destruction of endothelial cells does not seem to be the main mechanism of action of hemorrhagic SVMPs [35].
SVMPs cause loss of vascular integrity mainly by interfering with adhesion between endothelial cells. SVMPs can cleave adherent proteins present in the membrane of endothelial cells, such as cadherins, and, degrade proteins that constitute the basement membrane, such as type IV collagen and laminin, contributing to the disintegration of the basement membrane [36]. This action is similar to the activity of endogenous metalloproteinases that act in the remodeling of protein fibers present in the extracellular matrix.
BnP1 from B. asper, is a class P-I SVMP capable of degrading type IV collagen, which suggests that the degradation of the basement membrane is primarily carried out by the catalytic activity of the M domain. [37]. The hemorrhagic activity of some SVMPs can still occur through other mechanisms, such as the hemorrhagic factor 3 (HF3) present in B. jararaca, which cleaves proteoglycans present in the glycocalyx of endothelium cells, which contributes to their damage [31].
SVMPs normally also contribute to the thrombocytopenia seen in envenomation and also inhibit platelet aggregation induced by endogenous agonists. Normally, the inhibition of aggregation caused by SVMPs results from interference in the interaction between agonists and their respective receptors and integrins present in platelets. Contrary to the alterations caused in the endothelium, the platelet hypoaggregation induced by SVMPs has little relation with the catalytic activity of the M domain [38]. Furthermore, it has already been demonstrated that the D domain isolated from some SVMPs, such as jararhagin, is capable of inhibiting collagen- induced platelet aggregation, probably by interfering with α2β1 integrin. Although less studied, the C domain also seems to be of great importance in the hypoaggregating action of SVMPs [39]. In class P-II SVMPs, the RGD motif in the D domain was also identified, which interacts with the αIIB β3/GpIIb/IIIa integrin, inhibiting platelet aggregation. Although RGD may present variations in some P-II class SVMPs and even be substituted in several P-III class SVMPs, in general the D domain (or D-type domain in the case of P-III) maintains its ability to inhibit platelet aggregation [29].
It has also been shown that some SVMPs, such as jararhagin, can cleave the Von Willebrand factor (VWF), which prevents its correct interaction with the GPIb-IX-V receptor, resulting in hypoaggregation [39]. It is worth mentioning that some SVMPs, even with a hypoaggregating effect, are capable of inducing platelet aggregation in vitro. For example, basparin A catalyzes the conversion of prothrombin to thrombin, and the latter, being a powerful platelet agonist, will trigger the occurrence of platelet aggregation [38].
SVMPs may also contribute to the coagulopathy seen in envenomation, particularly by activating FX and/or prothrombin (Figure 1) [40]. The SVMPs that activate FX are all classified as P-III because they are composed of a larger subunit, which corresponds to a P-III class SVMP, and a smaller subunit, which corresponds to a snaclec (C-type lectin-like protein). Prothrombin activating SVMPs can be of all classes and are divided into group A, whose activity is independent of cofactors, and group B, which act only in the presence of calcium. In both groups, SVMP converts prothrombin to meizothrombin, which then undergoes autolysis, giving rise to thrombin [39, 31].
SVMPs of all classes can present fibrinolytic and fibrinogenolytic activity that, associated with the activation of FX and prothrombin, contributes to the rapid consumption of clotting factors during envenomation without the formation of a stable fibrin network, which favors incoagulability of the blood (Figure 1). In addition, some SVMPs, such as jararhagin, are able to activate the fibrinolytic system by reducing the interaction between tPA and PA-1 and reducing the activity of α2-antiplasmin, contributing to the worsening of coagulopathy in envenomation [31].
![Figure 1: Main alterations in the coagulation cascade caused by SVMPs and thrombin-like enzymes. SVMPs activate factor X and prothrombin leading to a large generation of thrombin. However, fibrin generation is relatively low due to the fibrino (geno) lytic activity of other SVMPs and thrombin-like enzymes. Lippi G, et al. [40].](/fulltextimages/8475/fig_1.png)
C-type Lectins
C-type lectins are homodimeric and non-enzymatic proteins that present the carbohydrate recognition domain (CRD), a domain that, in the presence of calcium ions, binds to carbohydrates. In the venom of snakes of the genus Bothrops, the so-called snake C-type lectins-like (snaclecs) are also present, which despite having some homology with the CRD, do not bind to carbohydrates and are not calcium dependent. These proteins are present in greater amounts in the venom than true type C lectins and, unlike these, are heterodimeric, composed of α and β subunits. These toxins have as main biological functions the action of modulating the activity of platelets and are anticoagulants [42, 43].
Among the snaclecs that interfere with platelet aggregation are those that can bind to GPIb, such as the GPIb-binding jararaca protein. GPIb is a glycoprotein that participates in the GPIb-IX-V complex, which contributes to platelet activation by interacting with VWF under high shear conditions. Normally, this interaction is inhibited with the binding of snaclec to GPIb. However, other effects resulting from the action of this type of snaclecs can be diverse, e.g.: in some cases snaclecs form multimers that favor platelet agglutination while other snaclec can induce platelet activation directly via GPIb [44].
There are also snaclecs that bind directly to VWF, as is the case with botrocetin, found in B. jararaca. This snaclec has a negatively charged concave region that binds to the A1 domain of VWF and induces its binding to GPIb, leading to platelet agglutination. There are still other snaclecs that can induce platelet activation by binding to GPVI or α2β1 integrin, glycoproteins that are normally important in platelet activation by interacting with collagen. Recently, the first described snaclec that inhibits epinephrine-induced platelet aggregation was isolated. This snaclec is bateltin, obtained from the venom of B. alternatus [45, 46]. The main pathways of interference of platelet activity by snaclecs are described in Figure 2.
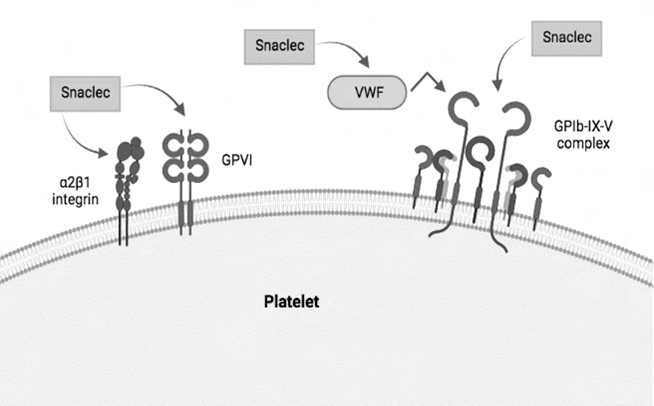
The venom of snakes from the Bothrops genus have snaclecs with anticoagulant activity, which is conferred by the calcium-dependent, high affinity binding of these proteins to the Gla domains of clotting factors. Among the anticoagulant snaclecs are IX/X-bp that binds both factor IX and factor X, IX-bp that bind to factor IX, and X-bp that bind to factor X. In addition, bothrojaracin, identified and purified from B. jararaca venom, is an anticoagulant snaclec that forms a non-covalent complex with thrombin. This binding probably occurs at the thrombin exosite 1 and does not alter the catalytic activity of this enzyme on synthetic peptides [45].
PLA2s
The phospholipases A2 (PLA2s) are lipolytic enzymes that hydrolyze glycerophospholipids at the sn-2 position generating lysophospholipids and fatty acids. PLA2s active site consists of His48, Asp49, Tyr52, Asp99, and a water molecule that plays a role in the catalysis, being connected in the residues His48 and Asp49 through hydrogen bonds [47]. They are calcium-dependent and consist of 125-130 amino acid residues cross-linked and stabilized by seven disulfide bonds [48]. PLA2 is one of the major components in Bothrops venom. A single snake venom PLA2 (svPLA2s) fraction can contain several isoforms of PLA2s, and they may be classified as acidic, basic or neutral, according to their net charge. Although svPLA2s share most of their structure, snake venom PLA2 enzymes differ greatly in their pharmacological properties and biological activities [49].
Plasma phospholipids play a critical role in the formation of coagulation complexes, and PLA2s may be the primary mechanism to the anticoagulant effect of snake venoms, through the hydrolysis of phospholipids surface [50, 51]. Mostly PLA2s act on blood clotting through the enzymatic hydrolysis of procoagulant phospholipids of plasma, or simply by binding to blood coagulation factors and/or phospholipids making them unavailable for the reactions of the coagulation cascade. The PLA2 anticoagulant activity can be either dependent or independent on the presence of phospholipids [52, 53].
In Bothrops genus venom, several acidic phospholipases have been described. BaCol PLA2 that is purified from B. asper venom has moderated anticlotting activity, while Bmoo
PLA2, purified from B. moojeni venom has high anticlotting activity on aPTT using human plasma, as well as antiplatelet activity, inhibiting platelet aggregation induced by collagen or ADP [54, 55]. According to Kini and Evans, the anticoagulant region, that is between amino acid residues 54 to 77, is positively charged in strong anticoagulants facilitating the interaction between these enzymes with negatively charged surfaces and thus, impairing blood clotting [56].
PLA2 plays a central role in platelet activation by hydrolysis of membrane phospholipids in response to a variety of stimuli. Yuan Y, et al. [57] demonstrated that snake venom PLA2s are dependent on high-density lipoproteins and other lipoprotein fractions, possibly to generate lysophosphatidylcholine-enriched HDL particles that modulate platelet function [58]. Based on their platelet activity, svPLA2s enzymes can be classified in three groups: PLA2s that induce platelet aggregation; PLA2s that inhibit platelet aggregation and PLA2S that have biphasic response to platelets [56]. In fact, several Bothrops PLA2s inhibit platelet aggregation [59, 60, 61].
The majority of effects of svPLA2s is exerted by hydrolysis of membrane phospholipids, although different isoforms may display different membrane damaging activity and consequently, different effects on the human hemostasis. In general, there are few reports on the membrane hydrolyzing property of svPLA2s, and they are better characterized according to their myotoxic activity. Therefore, the complete mechanism of membrane damage and toxicity from PLA2s enzymes is not entirely understood [49].
Other Venom Components
Bradykinin potentiating peptides (BPPs) were first identified in the venom of B. jararaca in the 1965 by Ferreira SH, et al. [62]. In general, the term BPP describes an effect and not structure, once there are many related and unrelated peptide sequences that potentialize bradykinin (BK). BPPs are proline-rich peptides that inhibit the angiotensin converting enzyme (ACE) and are the molecular basis for the development of captopril, antihypertensive drug used clinically [63]. The ACE is responsible for converting angiotensin (Ang) I into Ang II, which act as a vasoconstrictor and hypertensive agent. By inhibiting this enzyme, BPPs inhibit formation of Ang II and thus, reduce the blood pressure. ACE also cleaves bradykinin, a hypotensive vasoactive peptide [64]. ACE inhibitor such as BPPs potentiates the effect of BK by inhibiting its degradation. Since there is no generation of Ang II and bradykinin is not degraded, the angiotensin-system effects are decreased and the kallikrein-kinin system is potentiated [63]. Several BPPs were identified in Bothrops venom and more are still being characterized [65, 66].
Snake venom L-amino acid oxidase (sv-LAAO) is a flavoenzyme with molecular mass from 120-150 kDa in dimeric form and 55-66 kDa in monomeric form which stereospecifically catalyzes the oxidative deamination of an L-amino acid to an α-keto acid with concomitant production of ammonia and hydrogen peroxide [67]. These enzymes can either inhibit or activate platelet aggregation. Sv-LAAOs purified from Bothrops, such as BpirLAAO-I, BmooLAAO-I and Balt-LAAO-I induced dose-dependent platelet aggregation [68, 69, 70]. The mechanism for platelet aggregation induced by sv-LAAOs it is not fully understood, although earlier reports suggest that the platelet aggregation is induced by the formation of hydrogen peroxide and subsequent synthesis of thromboxane A2 [71]. The H2O2 produced by LAAOs mimics the effect of oxidases on platelets and at concentrations of 9mM or higher, induces the first phase of platelet aggregation [72]. Even though LAAOs are mainly characterized for their action on platelets, Sakurai Y, et al. [73] described a sv-LAAO that prolonged the activated partial thromboplastin time (aPTT) dose-dependently by inhibiting factor IX and it is fathomable that others sv-LAAOs may also have anticlotting properties.
There are other components, present in low concentrations in Bothrops venoms that play a role in the pathophysiology of snakebite victims. Hyaluronidase is a common enzyme on snake venoms that destroys the glycosidic bonds of connective tissues components, resulting in decreased tissue viscosity and allowing the penetration of the venom [74]. Snake venom vascular endothelial growth factors (svVEGFs) show a potent hypotensive effect and are crucial regulators of blood vessel formation and vascular permeability, also facilitating venom diffusion into tissues [75].
Conclusions
The pharmacological effects of Bothrops venom include cardiotoxicity, neurotoxicity, haemolytic, myotoxicity, haemostatic, haemorrhagic and edema. The coagulopathy seem in Bothrops envenomation is the outcome of a multifocal toxicity of distinct venom components involving the endothelium, platelets and coagulation. In Bothrops envenomation, bleeding is more usual than thrombosis, although this may vary depending on a series of factors including species, age, geographic location and venom components. The major components in Bothrops venom favors the bleeding tendency; Inoculation of SVMPs and SVSPs leads to endothelial dysfunction and activation of the coagulation cascade while also having fibrinolytic/fibrinogenolytic activity, making clot formation unfeasible thus, leading to a consumption coagulopathy. Since other components also act on several hemostatic systems, the bleeding and thrombosis caused by snakebite envenomation should be regarded as two sides of the same pathophysiological process. The entirety of the mechanism responsible for the pathophysiology is not understood, and further understanding of the events that happen following snakebites by toxin components is a good strategic to improve our knowledge of the accident pathophysiology. Besides this, a molecular comprehension of the hemostatic alterations caused by each component on Bothrops venom may help identify new molecules and targets for therapeutic applications.
Conflict of Interest
The authors declare no conflicts of interest.
Funding
We are grateful to the Instituto Butantan, CAPES, FAPESP and CNPq for supporting us in writing this article.
References
-
(2021) Snakebite Envenoming: an interactive data platform to support the 2030 targets. World Health Organization.
-
(2020) SINAN - Notifiable Diseases Information System: accident by venomous animals. Brasilia: Ministry of Health.
-
Kasturiratne A, Wickremasinghe AR, Silva N de, Gunawardena NK, Pathmeswaran A, et al. (2008) The Global Burden of Snakebite: A Literature Analysis and Modelling Based on Regional Estimates of Envenoming and Deaths. PLOS Medicine 5(11): e218.
-
Gutiérrez JM, Warrell DA, Williams DJ, Jensen S, Brown N, et al. (2013) The Need for Full Integration of Snakebite Envenoming within a Global Strategy to Combat the Neglected Tropical Diseases: The Way Forward. PLOS Neglected Tropical Diseases 7(6): e2162.
-
(2021) Visão 2050. CEBDES
-
(2021) Ministry of Health. Health Surveillance Department. Epidemiological Bulletin. Brazil.
-
Cardoso JLC, França FOS, Wen FH, Malaque CMS e Haddad V (2003) Animais peçonhentos no Brasil: biologia, clínica e terapêutica dos acidentes. São Paulo: Sarvier, pp: 72-84.
-
SilvaGM da, Souza DHB de, Waitman KB, Ebram MC, Fessel MR, et al. (2021) Design, synthesis, and evaluation of Bothrops venom serine protease peptidic inhibitors. Journal of Venomous Animals and Toxins Including Tropical Diseases 27.
-
Da Galizio NC, Serino-Silva C, Stuginski DR, Abreu PAE, Sant’Anna SS, et al. (2018) Compositional and functional investigation of individual and pooled venoms from long-term captive and recently wild-caught Bothrops jararaca snakes. J Proteomics 186: 56-70.
-
Nicolau CA, Prorock A, Bao Y, Costa Neves-Ferreira AG da, Valente RH, et al. (2018) Revisiting the Therapeutic Potential of Bothrops jararaca Venom: Screening for Novel Activities Using Connectivity Mapping. Toxins 10(2): 69.
-
Costa J de O, Fonseca KC, Garrote-Filho MS, Cunha CC, de Freitas MV, et al. (2010) Structural and functional comparison of proteolytic enzymes from plant latex and snake venoms. Biochimie 92(12): 1760-1765.
-
Serrano SMT (2013) The long road of research on snake venom serine proteinases. Toxicon 62: 19-26.
-
Polgár L (2005) The catalytic triad of serine peptidases. Cell Mol Life Sci 62(19–20): 2161-2172.
-
Pradniwat P, Rojnuckarin P (2014) Snake venom thrombin-like enzymes. Toxin Reviews 33(1-2): 16–22.
-
Kini RM (2005) Serine Proteases Affecting Blood Coagulation and Fibrinolysis from Snake Venoms. Pathophysiology of Haemostasis and Thrombosis 34(4- 5): 200-204.
-
Murakami MT, Arni RK (2005) Thrombomodulin- independent Activation of Protein C and Specificity of Hemostatically Active Snake Venom Serine Proteinases. Journal of Biological Chemistry 280(47): 39309-39315.
-
Matsui T, Fujimura Y, Titani K (2000) Snake venom proteases affecting hemostasis and thrombosis. Biochimica Et Biophysica Acta 1477(1-2): 146-156.
-
Braud S, Bon C, Wisner W (2000) Snake venom proteins acting on hemostasis. Biochimie 82(9-10): 851-859.
-
Aronson DL (1976) Comparison of the actions of thrombin and the thrombin-like venom enzymes ancrod and batroxobin. Thrombosis and Haemostasis 36(1): 9-13.
-
Xu G, Liu X, Zhu W, Yin Q, Zhang R, et al. (2007) Feasibility of treating hyperfibrinogenemia with intermittently administered batroxobin in patients with ischemic stroke/transient ischemic attack for secondary prevention. Blood Coagulation & Fibrinolysis: An International Journal in Haemostasis and Thrombosis 18(2): 193-197.
-
Vu TT, Stafford AR, Leslie BA, Kim PY, Fredenburgh JC, et al. (2013) Batroxobin Binds Fibrin with Higher Affinity and Promotes Clot Expansion to a Greater Extent than Thrombin Journal of Biological Chemistry, 288(23): 16862-16871.
-
Santos BF, Serrano SM, Kuliopulos A, Niewiarowski S (2000) Interaction of viper venom serine peptidases with thrombin receptors on human platelets. FEBS Letters 477(3): 199-202.
-
Niewiarowski S, Kirby EP, Brudzynski TM, Stocker K (1979) Thrombocytin, a serine protease from Bothrops atrox venom. 2. Interaction with platelets and plasma- clotting factors. Biochemistry 18(16): 3570-3577.
-
Zaqueo KD, Kayano AM, Domingos TFS, Moura LA, Fuly AL, et al. (2016) BbrzSP-32, the first serine protease isolated from Bothrops brazili venom: Purification and characterization. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 195: 15-25.
-
Zaqueo KD, Kayano AM, Simões-Silva R, Moreira-Dill LS, Fernandes CFC, et al. (2014) Isolation and Biochemical Characterization of a New Thrombin-Like Serine Protease from Bothrops pirajai Snake Venom. BioMed Research International 1-13.
-
Sant’ Ana CD, Ticli FK, Oliveira LL, Giglio JR, Rechia CGV, et al. (2008) BjussuSP-I: A new thrombin-like enzyme isolated from Bothrops jararacussu snake venom. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 151(3): 443-454.
-
Paes Leme AF, Prezoto BC, Yamashiro ET, Bertholim L, Tashima AK, et al. (2008) Bothrops proteaseA, a unique highly glycosylated serine proteinase, is a potent, specific fibrinogenolytic agent. Journal of Thrombosis and Haemostasis 6(8): 1363-1372.
-
Bhat SK, Joshi MB, Ullah A, Masood R, Biligiri SG, et al. (2016) Serine proteinases from Bothrops snake venom activates PI3K/Akt mediated angiogenesis. Toxicon 124: 63-72.
-
Olaoba O, Karina dos Santos P, Selistre-de-Araujo HS, Ferreira de Souza DH (2020) Snake Venom Metalloproteinases (SVMPs): A structure-function update. Toxicon X 7: 100052.
-
Kang TS, Georgieva D, Genov N, Murakami MT, Sinha M, et al. (2011) Enzymatic toxins from snake venom: Structural characterization and mechanism of catalysis. The FEBS Journal 278(23): 4544-4576.
-
Larreche S, Chippaux JP, Chevillard L, Mathe S, Resiere D, et al. (2021) Bleeding and Thrombosis: Insights into Pathophysiology of Bothrops Venom-Related Hemostasis Disorders. International Journal of Molecular Sciences 22(17): 9643.
-
Takeda S, Takeya H, Iwanaga S (2012) Snake venom metalloproteinases: Structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics 1824(1): 164-176.
-
Brenes O, Munoz E, Roldan-Rodriguez R, Diaz C (2010) Cell death induced by Bothrops asper snake venom metalloproteinase on endothelial and other cell lines. Experimental and Molecular Pathology 88(3): 424-432.
-
Bhat SK, Joshi MB, Vasishta S, Jagadale RN, Biligiri SG, et al. (2021) P-I metalloproteinases and L-amino acid oxidases from Bothrops species inhibit angiogenesis. Journal of Venomous Animals and Toxins Including Tropical Diseases, pp: 27.
-
Kawano J, Anai K, Sugiki M, Yoshida E, Maruyama M (2002) Vascular endothelial cell injury induced by Bothrops jararaca venom; non-significance of hemorrhagic metalloproteinase. Toxicon 40(11): 1553- 1562.
-
Senise LV, Yamashita KM, Santoro ML (2015) Bothrops jararaca envenomation: Pathogenesis of hemostatic disturbances and intravascular hemolysis. Exp Biol Med (Maywood) 240(11): 1528-1536.
-
Baldo C, Jamora C, Yamanouye N, Zorn TM, Moura-da- Silva AM (2010) Mechanisms of Vascular Damage by Hemorrhagic Snake Venom Metalloproteinases: Tissue Distribution and In Situ Hydrolysis. PLoS Negl Trop Dis 4(6): e727.
-
Loria GD, Rucavado A, Kamiguti AS, Theakston RDG, Fox JW, et al. (2003) Characterization of ‘basparin A,’ a prothrombin-activating metalloproteinase, from the venom of the snake Bothrops asper that inhibits platelet aggregation and induces defibrination and thrombosis. Arch Biochem Biophys 418(1): 13-24.
-
Kini RM, Koh CY (2016) Metalloproteases Affecting Blood Coagulation, Fibrinolysis and Platelet Aggregation from Snake Venoms: Definition and Nomenclature of Interaction Sites. Toxins 8(10): 284.
-
Yamashita KM, Alves AF, Barbaro KC, Santoro, ML (2014) Bothrops jararaca Venom Metalloproteinases Are Essential for Coagulopathy and Increase Plasma Tissue Factor Levels during Envenomation. PLOS Neglected Tropical Diseases 8(5): e2814.
-
Lippi G, Franchini M, Targher G (2011) Arterial thrombus formation in cardiovascular disease. Nat Rev Cardiol 8(9): 502-512.
-
Arlinghaus FT, Eble JA (2012) C-type lectin-like proteins from snake venoms. Toxicon 60(4): 512-519.
-
Lu Q, Navdaev A, Clemetson JM, Clemetso KJ (2005) Snake venom C-type lectins interacting with platelet receptors. Structure–function relationships and effects on haemostasis. Toxicon 45(8): 1089-1098.
-
Clemetson KJ, Lu Q, Clemetson JM (2005) Snake C-Type Lectin-Like Proteins and Platelet Receptors. Pathophysiology of Haemostasis and Thrombosis 34: 150-155.
-
Morita T (2005) Structures and functions of snake venom CLPs (C-type lectin-like proteins) with anticoagulant-, procoagulant-, and platelet-modulating activities. Toxicon 45(8): 1099-1114.
-
Pereira DF da C, Matias Ribeiro MS, de Sousa Simamoto BB, Dias EHV, Dantas NO, et al. (2021) Baltetin: A new C-type lectin-like isolated from Bothrops alternatus snake venom which act as a platelet aggregation inhibiting. Journal of Chromatography B 1173: 122695.
-
Kang TS, Georgieva D, Genov N, Murakami MT et al. (2011) Enzymatic toxins from snake venom: Structural characterization and mechanism of catalysis. The FEBS Journal 278: 4544-4576.
-
Dennis EA (1994) Diversity of group types, regulation, and function of phospholipase A2. The Journal of Biological Chemistry 269(18): 13057-13060.
-
Saikia D, Mukherjee AK (2017) Anticoagulant and Membrane Damaging Properties of Snake Venom Phospholipase A2 Enzymes. _In_: Inagaki H, et al. (Eds.), Snake Venoms, pp: 87-104.
-
Doley R, King GF, Mukherjee AK (2004) Differential hydrolysis of erythrocyte and mitochondrial membrane phospholipids by two phospholipase A2 isoenzymes NK-PLA2-I NK-PLA2-II from the venom of the Indian monocled cobra Naja kaouthia. Archives of Biochemistry and Biophysics 425(1): 1-13.
-
Kini RM (2005) Structure-function relationships and mechanism of anticoagulant phospholipase A2 enzymes from snake venoms. Toxicon: Official Journal of the International Society on Toxinology 45(8): 1147-1161.
-
Stefansson S, Kini RM, Evans HJ (1989) The inhibition of clotting complexes of the extrinsic coagulation cascade by the phospholipase A2 isoenzymes from Naja nigricollis venom. Thrombosis Research 55(4): 481-491.
-
Saikia D, Thakur R, Mukherjee AK (2011) An acidic phospholipase A(2) (RVVA-PLA(2)-I) purified from Daboia russelli venom exerts its anticoagulant activity by enzymatic hydrolysis of plasma phospholipids and by non-enzymatic inhibition of factor Xa in a phospholipids/ Ca(2+) independent manner. Toxicon: Official Journal of the International Society on Toxinology 57(6): 841-850.
-
Posada Arias S, Rey-Suárez P, Pereáñez JA, Acosta C, Rojas M, et, al. (2017) Isolation and Functional Characterization of an Acidic Myotoxic Phospholipase A2 from Colombian Bothrops asper Venom. Toxins 9(11): 342.
-
Silveira LB, Marchi-Salvador DP, Santos-Filho NA, Silva FP, Marcussi S (2013) Isolation and expression of a hypotensive and anti-platelet acidic phospholipase A2 from Bothrops moojeni snake venom. Journal of Pharmaceutical and Biomedical Analysis 73: 35-43.
-
Kini RM, Evans HJ (1987) Structure-function relationships of phospholipases. The anticoagulant region of phospholipases A2. J Biol Chem 262(30): 14402-14407.
-
Yuan Y, Jackson SP, Newnham HH, Mitchell CA, Salem HH, et al. (1995) An essential role for lysophosphatidylcholine in the inhibition of platelet aggregation by secretory phospholipase A2. Blood 86(11): 4166–4174.
-
Curcic S, Holzer M, Pasterk L, Knuplez E, Eichmann TO, et al. (2017) Secretory phospholipase A2 modified HDL rapidly and potently suppresses platelet activation. Sci Rep 7(1): 8030.
-
Andrião-Escarso SH, Soares AM, Fontes MR, Fuly AL, Corrêa FM, et al. (2002) Structural and functional characterization of an acidic platelet aggregation inhibitor and hypotensive phospholipase A2 from Bothrops jararacussu snake venom. Biochem Pharmacol 64(4): 723-732.
-
Teixeira SS, Silveira LB, da Silva FM, Marchi-Salvador DP, Silva FP, et al. (2011) Molecular characterization of an acidic phospholipase A(2) from Bothrops pirajai snake venom: Synthetic C-terminal peptide identifies its antiplatelet region. Arch Toxicol 85(10): 1219-1233.
-
Santos-Filho NA, Silveira LB, Oliveira CZ, Bernardes CP, Menaldo DL, et al. (2008) A new acidic myotoxic, anti- platelet and prostaglandin I2 inductor phospholipase A2 isolated from Bothrops moojeni snake venom. Toxicon 52(8): 908-917.
-
Ferreira SH (1965) A bradykinin-potentiating factor (bpf) present in the venom of _Bothrops_ jararaca. Br J Pharmacol Chemother 24(1): 163-169.
-
Sciani JM, Pimenta DC (2017) The modular nature of bradykinin-potentiating peptides isolated from snake venoms. Journal of Venomous Animals and Toxins Including Tropical Diseases 23(1): 45.
-
Dendorfer A, Wolfrum S, Dominiak P (1999) Pharmacology and cardiovascular implications of the kinin-kallikrein system. Jpn J Pharmacol 79(4): 403-426.
-
Ianzer D, Konno K, Marques-Porto R, Vieira Portaro FC, Stöcklin R, et al. (2004) Identification of five new bradykinin potentiating peptides (BPPs) from Bothrops jararaca crude venom by using electrospray ionization tandem mass spectrometry after a two-step liquid chromatography. Peptides 25(7): 1085-1092.
-
Da Silva Caldeira CA, Diniz-Sousa R, Pimenta DC, Dos Santos APA, Teles CBG, et al. (2021) Antimicrobial peptidomes of Bothrops atrox and Bothrops jararacussu snake venoms. Amino Acids 53(10): 1635-1648.
-
Ullah A (2020) Structure Function Studies and Mechanism of Action of Snake Venom L-Amino Acid Oxidases. Front Pharmacol 11: 110.
-
Izidoro LF, Ribeiro MC, Souza GR, Sant’Ana CD, Hamaguchi A, et al. (2006) Biochemical and functional characterization of an l-amino acid oxidase isolated from _Bothrops_ pirajai snake venom. Bio org Med Chem 14(20): 7034-7043.
-
Stábeli RG, Sant’Ana CD, Ribeiro PH, Costa TR, Ticli FK, et al. (2007) Cytotoxic l-amino acid oxidase from _Bothrops_ moojeni: Biochemical and functional characterization. Int J Biol Macromol 41(2): 132-1340.
-
Stábeli RG, Marcussi S, Carlos GB, Pietro RC, Selistre- de-Araújo HS, et al. (2004) Platelet aggregation and antibacterial effects of an l-amino acid oxidase purified from _Bothrops_ alternatus snake venom. Bioorg Med Chem 12(11): 2881-2886.
-
Du XY, Clemetson KJ (2002) Snake venom l-amino acid oxidases. Toxicon 40(6): 659-665.
-
Praticò D, Iuliano L, Ghiselli A, Alessandri C, Violi F, et al. (1991) Hydrogen Peroxide as Trigger of Platelet Aggregation. Haemostasis 21(3): 169-174.
-
Sakurai Y, Shima M, Matsumoto T, Takatsuka H, Nishiya K, et al. (2003) Anticoagulant activity of M-LAO, l-amino acid oxidase purified from Agkistrodon halys blomhoffii, through selective inhibition of factor IX. Biochim Biophys Acta 1649(1): 51-57.
-
Bala E, Hazarika R, Singh P, Yasir M, Shrivastava R, et al. (2018) A biological overview of Hyaluronidase: A venom enzyme and its inhibition with plants materials. Materials Today: Proceedings 5(2 Part 1): 6406-6412.
-
Yamazaki Y, Matsunaga Y, Tokunaga Y, Obayashi S, Saito M, et al. (2009) Snake Venom Vascular Endothelial Growth Factors (VEGF-Fs) Exclusively Vary Their Structures and Functions among Species. J Biol Chem 284(15): 9885- 9891_._
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells