A Bird Community Survey of Chinese Fir, Masson Pine and Eucalyptus Plantations in South China- A Case Study
Transect surveys were conducted in Chinese fir, Masson pine and eucalyptus plantations near Liuzhou, Guangxi in south China to determine bird species richness and abundance in the different forest types. The numbers of bird species seen or heard during this survey were recorded. Data was also collected from the surveyed forests to determine the canopy profiles of these forests. A total of 529 individuals representing 30 different species were recorded during this study. Based on Shannon’s diversity index, Masson pine forests were significantly more diverse in bird species (2.1590) than Chinese fir (1.7850) or eucalyptus forests (1.6387). The distribution of bird species, or evenness, among all forests was similar as no one single species dominated any one forest. On average, significantly more bird species were observed in Masson pine forests compared to eucalyptus and Chinese fir forests. When bird species were grouped by dietary guild, there were no significant differences between forest types for carnivores, frugivores, and omnivores. However, significantly more granivores and insectivores were observed in Chinese fir and Mason pine forests. Forest canopy profiles revealed that the canopy profile of Chinese fir was more narrow compared to the more elongated profiles of Masson pine and eucalyptus. The fact that more species and individuals were discovered in the Masson pine and eucalyptus forests may be a result of bird preferences for shelter in the upper levels of vegetation because taller structures offer a larger area of refuge. While plantations may not provide the same amount and type of diversity that native forests provide, they still have value in providing a level of diversity that might otherwise be absent in other land use situations.
Introduction
Rapid economic growth in China over the past decade has created a timber deficit to exist in China’s domestic timber supply. Subsequently, China has become the world’s largest importer of logs and wood pulp, and the second largest importer of lumber and woodchips [1]. In response to this timber deficit China continues to implement policy measures and land tenure reforms that will increase the quantity and quality of its forests. China’s
Journal of Ecology & Natural Resources
State Forest Administration (SFA) is attempting to increase its domestic timber supply from the 180 million m3/year recorded in 2010 to a projected 300 million m3/year by 2020 [2]. In efforts to reduce this timber deficit, China has been rapidly expanding it forest area. Approximately 129 million hectares (62%) are primary and naturally regenerated forests, while 79 million hectares (38%) are planted forests [2]. Accordingly, China now has the largest plantation area in the world, principally of fast-growing species, particularly eucalyptus, introduced from Australia. The total area of eucalyptus in China has grown from an estimated area of 670,000 hectares in the mid- 1990s to 3.6 million hectares in 2011, to its present estimation of at least 4 million hectares [3, 4, 5, 6]. These plantations occur primarily in Guangdong, Guangxi, Hainan Island, Yunnan, Sichuan and Fujian provinces with more than 1.4 million hectares in Guangxi alone [7]. The eucalyptus clone, Eucalyptus urophylla × E. grandis, selected by Dongmen Forest Farms in Guangxi [8], is the species most widely planted in south China because of its adaptability and stable rapid growth, producing as much as 30 m3/ha/yr [9]. However the introduction of eucalyptus has been controversial. Other than the disputes arising between corporate eucalyptus plantations and the use of land by farmers, there are ecological concerns regarding conversion of large areas of natural forests and farmland. One of the pro-arguments for the vast areas of planted eucalyptus in south China is that it restores a level of biodiversity on lands that have been in agriculture for extensive periods of time. One has to be careful about claims that reforestation of any degraded land would offset any loss of species following deforestation [10]. While it is known that the conversion of primary forests to intensive agriculture leads to dramatic losses in biodiversity, there is far less certainty about the conservation to secondary and planted forests [11, 12, 13, 14]. Biodiversity in eucalyptus plantations has been widely studied since it is one of the major concerns regarding the management of this forest type [15, 16, 17, 18]. However, depending on forest location and specific management method these studies have found that management of eucalyptus plantations can encourage the regeneration of natural forest biodiversity in the understory and restore degraded land, or it can have an opposite effect by facilitating the recovery of light- demanding grasses and other pioneer weeds, thus suppressing the recruitment of native plants [19]. It is believed that allelopathogenic effects of the eucalyptus litter, which contains a variety of oils and resins from foliage, has an adverse effect on vegetation thus reducing the number and amount of understory species. While it has been suggested that biodiversity should be measured at multiple organizational, temporal and spatial levels [20], others have suggested that birds may serve as good indicators of biodiversity as they are susceptible to landscape-level changes in the environment [21, 22, 23, 24, 25]. For example, the forest avifauna community has been decimated in Hong Kong as a result of deforestation, fragmentation, and isolation from larger forest areas and thus reflects the appropriate landscape changes [26]. Regardless, there are concerns regarding the extent that even eucalyptus forests may contribute to biodiversity compared to native forests, or forests planted to native species. Daniels, et al. (1990) found that even though plantations increased bird diversity in the landscape, there was a gradual displacement of bird species composition from what was typical in the natural forests to more of what was found in urban and scrubby habitats, and this was most evident in the eucalyptus forests. Marsden, et al. [27] discovered that the number of bird species in eucalyptus forests was significantly reduced compared to forest reserves. This was the result of the intensive management of these eucalyptus forests that create a void of understory vegetation. Therefore it was the purpose of this paper to present the results of bird surveys conducted in planted forests of eucalyptus (Eucalyptus urophylla x E. grandis), Masson pine (Pinus massoniana) and Chinese fir (Cunninghamia lanceolata) near Liuzhou, Guangxi, in south China. It was desirous to examine how eucalyptus plantations compared to plantations of native species as part of the vast reforestation effort taking place in south China. While this study focuses on a very small area, it could provide some insights into the bird diversity events taking place in planted forests of south China
Methods
Study Area
The study was located in the forests of Guangxi Eco- Engineering Vocational and Technical College located near the city of Liuzhou in the northeast section of Guangxi Province. The forest (approximately N24o29’ latitude, E109o22’ longitude) occupies 1,215 hectares and is comprised mostly of planted stands of Chinese fir Journal of Ecology & Natural Resources
(Cunninghamia lanceolata), Masson pine (Pinus massoniana), and eucalyptus hybrid (Eucalyptus urophylla x E. grandis). A summary of the forest characteristics are displayed in Table 1. This region lies within the southeastern edge of the subtropical monsoon climate areas of China. The mean annual temperature within the region is 21.1oC, with a maximum of 40.3oC in the summer and − 1.5oC in winter. The average annual precipitation ranges from 1200 to 1500 mm with average relative humidity ranging from 80 to 84%. The main geomorphic types are lower mountains and hills, characterized by rolling topography, which are dominated by moderately developed arenaceous shale red soils and strongly developed clay arenaceous shale red soils [28]. A majority of the forests were near urban areas which included highways and roads that created noise and chemical pollution. Dwellings and farms were also present within the forests. Many of the areas contained chicken, duck, or vegetable farms with many seed sources. The forests with duck farms had large bodies of water (either a man-made pond or lake).
In terms of forest understory structure, Chinese fir had the most developed understory of all forests studied, being dominated by herbaceous species with more lichen on trees. The Masson pine forests likewise had understory structure, but it was less dense as compared to the Chinese fir forests. Many of the Masson pine forests had trees that were tapped to collect pine resin. The Masson pine forests also had a greater abundance of mosquitos and insects compared to the other forests. The eucalyptus forests are intensively managed, which includes the use of coppicing to regenerate the forest and the application of fertilizer the first two years. Harvesting occurs every 5 to 6 years, depending on the harvest schedule, and the root stock used for coppicing may be used through at least 4 harvest periods before the stumps are dug up and replanted. In spite of the application of fertilizer, the understory was the least developed of the three forest types, where in some cases it was essentially non-existent.
| Forest attribute | N1 | Mean | Minimum | Maximum | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chinese fir | ||||||||||||||
| Trees per ha | 11 | 1284 | 1080 | 1560 | ||||||||||
| Basal area (m2/ha) | 11 | 28.0 | 19.2 | 35.1 | ||||||||||
| Mean dbh (cm) | 11 | 16.6 | 13.7 | 19.0 | ||||||||||
| Stand age (yrs) | 11 | 29 | 17 | 40 | ||||||||||
| Eucalyptus | ||||||||||||||
| Trees per ha | 6 | 1210 | 1020 | 1360 | ||||||||||
| Basal area (m2/ha) | 6 | 16.4 | 13.0 | 19.1 | ||||||||||
| Mean dbh (cm) | 6 | 13.1 | 12.4 | 13.7 | ||||||||||
| Stand age (yrs) | 6 | 7 | 2 | 21 | ||||||||||
| Masson pine | ||||||||||||||
| Trees per ha | 12 | 1000 | 440 | 1800 | ||||||||||
| Basal area (m2/ha) | 12 | 29.7 | 23.4 | 39.0 | ||||||||||
| Mean dbh (cm) | 12 | 20.8 | 14.6 | 32.1 | ||||||||||
| Stand age (yrs) | 12 | 22 | 12 | 40 |
Table 1: Summary of forest attributes for stands sampled near Luizhou, Gunagxi Province, PR China.
1 N = number of forest stands sampled
Data Collection
Forest Data: Forests were sampled with three 0.05 ha circular plots in each forest. On each plot, the dbh was measured and recorded for each tree with the use of a d- tape to the nearest 0.1 cm. In order to quantify canopy structure in these forests, six to eight trees were selected on each sample plot for the purpose of measuring the crown diameter and crown length. The crown diameter of these selected trees was measured by first locating the major crown axis and marking it on the ground with wire flags. The minor axis was located by marking the crown diameter on the ground with wire flags that was 90o to the major axis. The distance between flags was measured and recorded to the nearest 0.1 m. The quadratic mean of these two measured diameters were determined and used Journal of Ecology & Natural Resources
as the average crown diameter of the tree. The total height and height-to-live-crown were measured to the nearest 1.0 m with a clinometer. The crown length was determined by the difference between total tree height and height-to-live-crown. Since bias is typically introduced with the clinometer, 12 trees that represent the range of tree sizes on sample plots were selected and felled to measure the actual total length (height) and length to live crown of the tree. These measured lengths was used to determine the standard error of the clinometer measurements, and applied to heights measured with the clinometer to achieve more accurate height measurements. Based on comparisons of height measurements from the clinometer with the measured lengths of the tree after they had been felled, a margin of measurement error of ± 0.90 meters and ± 1.68 meters was determined at p = 0.05 for total height and height-to-live-crown, respectively. Equations were developed from the measurement data of selected trees collected in all three years. These equations were used to estimate the total height, height-to-live-crown, and crown diameter of all trees on plots based on the measured dbh. A model of the form: Y = a(lnD)b Where Y = either total height, height to live crown, or crown diameter (m), D = dbh (cm.), and ln = natural logarithm, was used to develop prediction equations. The model coefficients were estimated using the Non-Linear Procedure, Gauss-Newton method in SAS software, version 9.2 (SAS Institute Inc., 2008) and are presented in Table 2. Crown diameter was determined by finding the quadratic mean of the major and minor axis measurements. These equations were applied to all remaining trees for which field measurements of height and crown diameter were not recorded.
| Dependent | Equation coefficients | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variableb | N | a | b | RMSE | R2 | F value | Pr > F | ||||||||||||||||
| H t | 131 | 1.9227 | 2.3320 | 2.5020 | 0.9858 | 4490.2 | <0.0001 | ||||||||||||||||
| H lc | 96 | 1.2022 | 2.4266 | 3.5416 | 0.9486 | 867.5 | <0.0001 | ||||||||||||||||
| D cr | 96 | 1.0260 | 1.4077 | 0.7626 | 0.9696 | 1499.4 | <0.0001 |
Table 2: Coefficients and statistics for the height and crown diameter modelsa using dbh (cm) as the independent variable Table 2
Table 2: Coefficients and statistics for the height and crown diameter modelsa using dbh (cm) as the independent variable Table 2: Coefficients and statistics for the height and crown diameter modelsa using dbh (cm) as the independent variable for eucalyptus (Eucalyptus urophylla x E. grandis) in northeast Guangxi Province, P.R. China. a Model: Y = a(lnD)b, where Y = total height, height to live crown, or crown diameter (m), D = dbh (cm) and ln = natural logarithm.
b Dependent variables are defined as: Ht = total height (m), Hlc = height to live crown (m), and Dcr = crown diameter (m) In order to acquire a good quantification of forest canopy volume and structure, a SAS algorithm developed by Avina, et al. (2007) was modified and used. The original algorithm was developed to quantify forest structure for bat habitat evaluation and used 3 meter increments in the separation of forest strata where a 12- plus meter was the last stratum. For our purposes 3 meter increments were too small and we needed quantify strata above 12 meters. Therefore, we broke the strata into 6 meter increments with 24-plus meters occupying the last stratum.
Bird Data
Bird observations were recorded within each of the three forest types along a 1000 m transect during June - July of 2011 and 2012. Great care was given to make sure that the same individual bird was not recorded multiple times. Each transect was walked four different days during the hours of 7:30 and 10:30 local time. The species of bird that was observed or heard while walking this transect was recorded (Table 3). According to Bird Life International, all species observed are listed as least concern, and no species are listed as near threatened, vulnerable, endangered or critically endangered. Observations of the forests were also recorded while traversing transects. Basic descriptions of the forest habitats and the man-made structures that were near them were recorded with the bird observations. The time when the observations were made was documented, and the weather conditions (current temperature; sunny, partly sunny, or cloudy; relative humidity) were measured with a Skymaster Weather Meter and recorded. Of the bird sampling events, 52% occurred when it was cloudy, 20% occurred when it was partly cloudy and 28% occurred when it was sunny. The average air temperature during the sampling events was 26oC, with a range of 24oC
Journal of Ecology & Natural Resources
– 32oC, and the average relative humidity was 80.1%, with a range of 70.6% – 88.9%. Shannon’s diversity and evenness indices were calculated and used to compare bird populations between the different forest types.
| Scientific Name | Common Name | ||||
|---|---|---|---|---|---|
| Abroscopus albogularis | Rufous-faced warbler | ||||
| Acridotheres cristatellus | Crested myna | ||||
| Aegithalos concinnus | Black-throated bushtit | ||||
| Centropus bengalensis | Lesser coucal | ||||
| Centropus sinensis | Greater coucal | ||||
| Copsychus saularis | Oriental magpie-robin | ||||
| Eudynamys scolopaceusa | Western koel | ||||
| Garrulax canorus | Chinese hwamei | ||||
| Garrulax sannio | White-browed laughingthrush | ||||
| Garrulus glandarius | Eurasian jay | ||||
| Lonchura punctulata | Scaly-breasted munia | ||||
| Lonchura striata | White-rumped munia | ||||
| Machlolophus spilonotus | Yellow-cheeked tit | ||||
| Nycticorax nycticorax | Black-crowned night heron | ||||
| Orthotomus sutorius | Common tailorbird | ||||
| Parus major | Great tit | ||||
| Pomatorhinus ruficollis | Streak-breasted scimitar babbler | ||||
| Prinia atrogularis | Hill prinia | ||||
| Prinia crinigera | Striated prinia | ||||
| Prinia flaviventris | Yellow-bellied prinia | ||||
| Prinia inornata | Plain prinia | ||||
| Pycnonotus aurigaster | Sooty-headed bulbul | ||||
| Pycnonotus jocosus | Red-whiskered bulbul | ||||
| Pycnonotus sinensis | Light-vented bulbul | ||||
| Spilopelia chinensis | Eastern spotted dove | ||||
| Spilornis cheela | Crested serpent eagle | ||||
| Spizixos semitorques | Collared finchbill | ||||
| Stachyridopsis ruficeps | Rufous-capped babbler | ||||
| Streptopelia orientalis | Oriental turtle dove | ||||
| Zosterops japonicus | Japanese white eye |
Table 3: The list of bird species recorded by observation of sight or sound in the forest stands sampled in Guangxi province, PR
Results
Bird Observations
A total of 529 individuals representing 30 different species were recorded during this study. Masson pine forests were significantly more diverse in bird species than Chinese fir or eucalyptus forests (Table 4). The bird diversity between Chinese fir and eucalyptus forests were statistically similar. The distribution of bird species, or evenness, among all forests was similar as no one single species dominated any one forest. On average, significantly more bird species were observed in Masson pine forests compared to eucalyptus and Chinese fir forests. All bird species that were recorded had been previously documented in a census conducted in south China, including Guangxi, in 2006, with the exception of the striated prinia, which was found only in the Masson pine forests.
| Lonchura punctulata | Scaly-breasted munia | ||
| Lonchura striata | White-rumped munia | ||
| Machlolophus spilonotus | Yellow-cheeked tit | ||
| Nycticorax nycticorax | Black-crowned night heron | ||
| Orthotomus sutorius | Common tailorbird | ||
| Parus major | Great tit | ||
| Pomatorhinus ruficollis | Streak-breasted scimitar babbler | ||
| Prinia atrogularis | Hill prinia |
Table 4: Comparison of mean values1 for bird species diversity, evenness of species distribution, and the number of species obser
Table 4: Comparison of mean values1 for bird species diversity, evenness of species distribution, and the number of species observed by forest type in Guangxi province, PR China. 1 Means values followed by the same letter are not significantly different within columns at p = 0.05, Duncan’s MRT. When considering the top five species that occur in these forests, Japanese white eye and the common tailorbird occur in all forest types (table 5). Species that are unique to a forest type when considering abundance are: a) spotted dove, light-vented bulbul, and white- rumped munia in Chinese fir, b) Chinese bulbul and streak-breasted scimitar babbler in eucalyptus, c) striated prinia and great tit in Masson pine forests. The greater coucal occurs within the top five species for both eucalyptus and Masson pine forests. With the exception of the striated prinia, these top five species were recorded at 22 – 89% of the sites in the 2006 survey of south China, and five species occurred at 50% or more of the sites. Even though the striated prinia did not appear in the Journal of Ecology & Natural Resources
2006 survey, it was not surprising to find this species as it is widely distributed across southern China, and extends southward into Taiwan, Vietnam, and Laos and westward into Nepal, Bhutan, Afghanistan and Pakistan.
| Chinese fir forest | Eucalyptus forest | Masson pine forest | |
|---|---|---|---|
| Japanese white eye | Japanese white eye | Japanese white eye | |
| Spotted dove | Chinese bulbul | Striated prinia | |
| Common tailorbird | Greater coucal | Great tit | |
| Light-vented bulbul | Common tailorbird | Common tailorbird | |
| White-rumped munia | Streak-breasted scimitar babbler | Greater coucal |
Table 5: The top five bird species1 observed in each forest type based upon the total number of individuals observed by species.
Table 5: The top five bird species1 observed in each forest type based upon the total number of individuals observed by species. The top five species occurrence in each forest type represents 62.3%, 55.6%, and 51.0% of all individuals observed in Chinese fir, Eucalyptus, and Masson pine, respectively. When bird species were grouped by dietary guild, there were no significant differences between forest types for carnivores, frugivores, and omnivores (Figure 1). However, significantly more granivores and insectivores were observed in Chinese fir and Mason pine forests, respectively, compared to the other forest types. The higher abundance of insectivores in the Masson pine forest is consistent with what Zhang et al. [29] found. The higher abundance of mosquitoes in the dark, damp forests of Masson pine was quite evident compared to other forests, which may help to explain the higher number of insectivores. The greater abundance of granivores in the Chinese fir stands may be explained by the fact that even though the seeds of Chinese fir ripen in October – November, the cones, which are attached near the branch tips, will remain on the trees for one or more years. Any remaining seeds in the cones would be highly visible. Frugivores and insectivores were the more common dietary guilds that were observed.
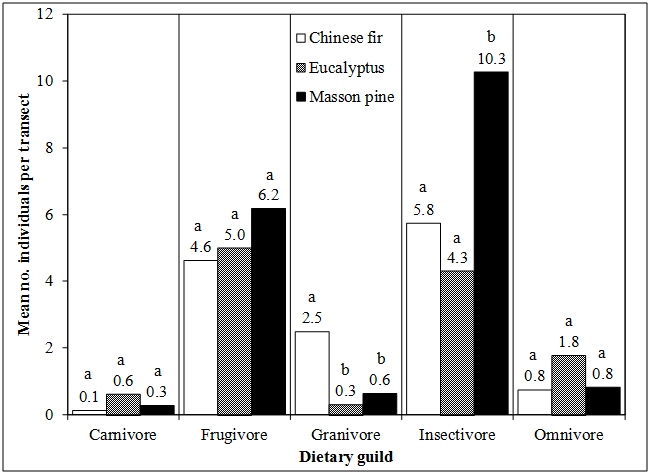
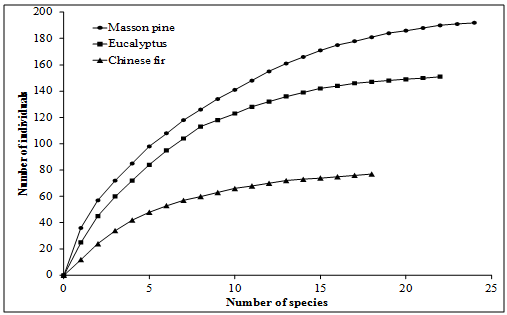
Figure 1: The mean number of individual birds grouped by diet category observed in the sampled Chinese fir, eucalyptus, and Masson pine forests of northeast Guangxi province, P. R. China. Means followed by the same letter are not significantly different between forest types at p = 0.05, Duncan’s MRT. The results of sample-based rarefaction curves (figure 2) illustrate the completeness of the transect inventories and the sufficiency of sampling efforts, due to their rapid approach to an asymptote. Masson pine forests had the highest numbers of individuals and species, reflecting high species richness of bird species. On the other hand, Chinese fir forests had the lowest numbers of individuals and species. Eucalyptus forests displayed more in terms of numbers of species and individuals than Chinese fir but less than Masson pine forests. However, eucalyptus forests were more closely aligned with Masson pine forests in this regard (Figure 2).
Forest Canopy Profile
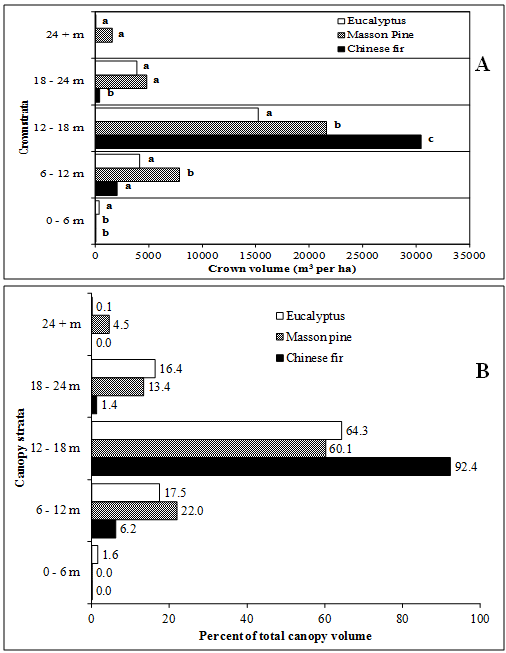
The majority of the canopy profiles of all forest types occurred in the 12 – 18 m stratum (figure 3). The live canopy profile of Chinese fir is rather narrow, occupying only the 6 – 12 m through the 18 – 24 m strata. In fact, 92.4% of the Chinese fir canopy occurred in the 12 – 18 m stratum, compared to 64.3% and 60.1% of the canopy for Masson pine and eucalyptus, respectively. The canopy of eucalyptus occurred in all strata while the canopy of Masson pine occurred in all but the 0 – 6 m stratum. Chinese fir had significantly more crown volume in the 12 – 18 m strata than Masson pine and eucalyptus and significantly less in the 18 – 24 m stratum.
Journal of Ecology & Natural Resources
Discussion and Conclusions
Of the 30 species observed, 22 were identified as either forest dependent or forest generalists, while the other 8 species are considered non-forest dependent. Zhang, et al. [29] found the Japanese white-eye one of three most common birds in Masson pine plantations, as well as primary mixed (pine and broadleaved) and mature broad- leaved forests. This study likewise found that the Japanese white-eye was the most common in the three forests studied here. While Zhang, et al. [29] found the oriental turtle dove to exist in the mature primary forest, the oriental turtle dove in this study was observed to occur in the eucalyptus and Chinese fir forests. Kwok and Corlett [26] found Japanese White-eye, Great Tit and Light-vented Bulbul the three most common species found in natural secondary forests and a Lophostemon confertus plantation in south China. This study found that the Light-vented bulbul was the fourth most common in the Chinese fir plantations and the Great tit was the third most common in the Masson pine plantation. The eucalyptus plantations contained these species but much less common. Zhang, et al. [29] also found that the Masson pine plantation was the least of the three forest types in species richness, comparing it to a primary mixed pine- broadleaved forest and a primary broadleaved forest. In this study, Masson pine plantations had the highest species richness, but this was in comparison to two other types of forests, namely Chinese fir and eucalyptus, both of which were plantations and not primary forests. It has been reported in many areas of south Asia that plantations typically have lower species richness and diversity compared to primary forests [26, 27, 28, 29, 30, 31]. It would appear in the case of this study that while a level of bird species richness existed in these plantations, they were still not as high as one would expect to see in primary forests. The Chinese fir plantation had the lowest diversity as well as the most flattened canopy profile. The Masson pine and eucalyptus forests were taller and had a more elongated canopy profile. The fact that more species and individuals were discovered in the Masson pine and eucalyptus forests may be a result of bird preferences for shelter in the upper levels of vegetation because taller structures offer a larger area of refuge and a better vantage point, potentially reducing the risk of predation [32].
Journal of Ecology & Natural Resources
The loss of habitat (including native forests) is one of the main contributors to global losses in biodiversity [33, 34], and while forest plantations of either native or non-native species may not always be viewed as the best remedial approach, they provide habitat that may otherwise not exist. For example, Loyn, et al. [35] found that the mean abundance of forest and woodland birds was higher in eucalypt plantations than cleared farmland, and marginally lower than in native forests. And in the context of the extensive urban settings in south China, plantations may provide the supplemental habitat structures that are lacking, thus creating a more diverse habitat setting conducive to more bird species. Caula, et al. [36] pointed out that greater bird diversity is seen in urban settings with mixed species woodlands (native and exotic; trees and shrubs; deciduous and coniferous) than in single species woodland stands as more species with different habitat requirements can make use of the greater diversity in floristic composition. Studies and logic would suggest that plantation forests do not provide the conservation value that natural native forests provide [37]. However, they can play a role by providing species richness at a broader landscape scale and connectivity across a landscape filled with forest fragments [38, 39, 40]. From this study we can conclude that eucalyptus plantations in south China are comparable to plantations of native species in regard to bird species abundance and richness. While there may be an overlapping effect due to the fact the different plantations were in close proximity to each other, eucalyptus plantations at least provide connectivity across the landscape and enhance the diversity in areas that may otherwise be void of forests. These plantations can add biodiversity in areas previously in agriculture and in areas where agriculture still exists. It can also increase the diversity of habitats in urban areas, thus making it more amiable to a greater variety of bird species.
Acknowledgements
The author would like to thank Margaret Smyth, Rachel Palmer, Anne Mauro, Rachael Crane, and Professors Yuhua Tao and Li-feng Tan for their assistance in the data collection and entry. The author would like to especially thank Profesor Li-feng Tan for her assistance with the bird identification. This project was funded by the China Research Abroad Program, School of Environment and Natural Resources at The Ohio State University.
References
-
RISI (2015) China Timber Supply Outlook. A Special Market Analysis Study. Resource Information Systems Inc. Bedford, Massachusetts.
-
FAO (2015) Global Forest Resources Assessment. Food and Agriculture Organization of the United Nations, Rome.
-
Midgley SJ, Pinyopusarerk K (1996) The role of eucalypts in local development in the emerging economies of China, Vietnam and Thailand. In: Eldridge, K.G., Crowe, M.P. and Old, K.M. (eds.) Environmental Management: the role of Eucalyptus and other fast growing species. Proc. of the Joint Australian/Japanese Workshop held in Australia. 23–27 October, 1995. CSIRO Forestry and Forest Products, Canberra 4-10.
-
Turnbull JW (2007) Development of sustainable forestry plantations in China: a review. ACIAR Impact Assessment Series Report No. 45. Australian Centre for International Agricultural Research, Canberra.
-
ITTO (2010) Report from China. ITTO Tropical Timber Market Report 15: 1-15.
-
Chen SX, Chen XF (2013) Technical Problems and Thinking on Eucalypt Plantation Management in China. Eucalypt Science & Technology 30: 52-59.
-
Liley B (2014) Focus on China: Guigang - Anatomy of a Hardwood Revolution. Presentation to: 'Forest Investment & Market Outlook' Conference, 14-15 April, 2014, Melbourne Australia.
-
Qi SX (2007) Applied Eucalypt cultivation in China. Beijing: China Forestry Publishing House.
-
Wu S, Xu J, Li G, Du Z, Lu Z, et al. (2011) Genotypic variation in wood properties and growth traits of Eucalyptus hybrid clones in southern China. New Forests 42(1): 35-50.
-
Gardner TA, Hernández MIM, Barlow J, Peres CA (2008) Understanding the biodiversity consequences of habitat change: the value of secondary and plantation forests for neotropical dung beetles. Journal of Applied Ecology 45(3): 883- 893. Journal of Ecology & Natural Resources
-
Dunn RR (2004) Managing the tropical landscape: a comparison of the effects of logging and forest conversion to agriculture on ants, birds, and Lepidoptera. Forest Ecology and Management 191: 215-224.
-
Wright SJ, Muller-Landau HC (2006) The Uncertain Future of Tropical Forest Species. Biotropica 38(4): 443-445.
-
Gardner TA, Barlow J, Parry LW, Peres CA (2007) Predicting the Uncertain Future of Tropical Forest Species in a Data Vacuum. Biotropica 39(1): 25-30.
-
Laurance WF (2007) Forest destruction in tropical Asia. Current Science 93(11): 1544-1550.
-
Parrotta JA (1995) Influence of overstory composition on understory colonization by native species in plantations on a degraded tropical site. Journal of Vegetation Science 6(5): 627-636.
-
Michelsen A, Lisanework N, Friis I, Holst N (1996) Comparisons of Under storey Vegetation and Soil Fertility in Plantations and Adjacent Natural Forests in the Ethiopian Highlands. Journal of Applied Ecology 33(3): 627-642.
-
Loumeto JJ, Huttel C (1997) Understory vegetation in fast growing tree plantations on savanna soils in Congo. For Ecol Manag 99: 65-81.
-
Chen QB (2001) A review of researches on biodiversity in Eucalyptus plantations. Chin J Trop Crops 4: 82-90.
-
Wen Y, Ye D, Chen F, Liu S, Liang H (2010) The changes of understory plant diversity in continuous cropping system of Eucalyptus plantations, South China. J For Res 15(4): 252-258.
-
Noss RF (1990) Indicators for Monitoring Biodiversity: A Hierarchical Approach. Conservation Biology 4(4): 355-364.
-
Wilcove DS (1985) Nest Predation in Forest Tracts and the Decline of Migratory Songbirds. Ecology 66(4): 1211-1214
-
Gregory RD, van Strien A, Vorisek P, Gmelig Meyling AW, Noble DG, et al (2005) Developing indicators for European birds. Phil. Trans. R. Soc. B, Biological Sciences 360(1454): 269-288.
-
Blair RB (1999) Birds and butterflies along an urban gradient: surrogate taxa for assessing biodiversity? Ecological Applications 9(1): 164-170.
-
Gregory RD, Noble D, Field R, Marchant J, Raven M, Gibbons DW (2003) Using birds as indicators of biodiversity. Ornis Hung 12-13: 11-24.
-
Garson J, Aggarwal A, Sarkar S (2002) Birds as surrogates for biodiversity: an analysis of a data set from southern Québec; J. Biosci 27(4): 347-360.
-
Kwok HK, Corlett RT (2000) The bird communities of a natural secondary forest and a Lophostemon confertus plantation in Hong Kong South China. Forest Ecology and Management 130(1-3): 227– 234.
-
Marsden SJ, Whiffin M, Galetti M (2001) Bird diversity and abundance in forest fragments and Eucalyptus plantations around an Atlantic forest reserve, Brazil. Biodiversity & Conservation 10(5): 737-751.
-
Zheng H, Ouyang Z, Xu W, Wang X, Miao H, et al (2008) Variation of carbon storage by different reforestation types in the hilly red soil region of southern China. For Ecol Manage 255: 1113-1121.
-
Zhang Q, Han R, Zou F (2011) Effects of artificial afforestation and successional stage on a lowland forest bird community in southern China. Forest Ecology and Management 261(11): 1738-1749.
-
Thiollay JM (1995) The role of traditional agroforests in the conservation of rain forest bird diversity in Sumatra. Conservation Biology 9(2): 335-353.
-
Castelletta M, Thiollay JM, Sodhi NS (2005) The effects of extreme forest fragmentation on the bird community of Singapore Island. Biological Conservation 121: 135-155.
-
Yuan B, Lu C (2016) Effects of urbanization on bird diversity: A case study in Yizhou, Guangxi Province, China. Asia Life Sciences 25(1): 79-96.
-
Houghton RA (1994) The worldwide extent of land- use change. Bioscience 44(5): 305-313.
-
Myers M, Mittermeir RA, Mittermeir CG, Da Fonseca GAB, Kent J (2000) Biodiversity hotspots for Journal of Ecology & Natural Resources conservation priorities. Nature 403(6772): 853- 858.
-
Loyn RH, McNabb EG, Macak P, Noble P (2007) Eucalypt plantations as habitat for birds on previously cleared farmland in south-eastern Australia. Biological Conservation 137: 533-548.
-
Barlow J, Mestrec LAM, Gardnera TA, Peresa CA (2007) The value of primary, secondary and plantation forests for Amazonian birds. Biological Conservation. 136: 212-231.
-
Hughes JB, Daily GC, Ehrlich PR (2002) Conservation of tropical forest birds in countryside habitats. Ecology Letters 5(1): 121-129.
-
Fischer J, Lindenmayer DB, Manning AD (2006) Biodiversity, ecosystem function, and resilience: ten guiding principles for commodity production landscapes. Frontiers in Ecology and the Environment 4(1): 80-86.
-
Caula S, de Villalobos AE, Marty P (2014) Seasonal dynamics of bird communities in urban forests of a Mediterranean city (Montpellier, Southern France). Urban Ecosystems 17(1): 11-26.
-
Ranjit Daniels RJ, Joshi NV, Gadg M (1990) Changes in the Bird Fauna of Uttara Kannada, India, in Relation to Changes in Land Use over the Past Century. Biological Conservation 52(1): 37-48.
- Lessons to Learn: Trees are More than the Lungs of the World
- Community Forestry Enterprises as a Model for Sustainable Forest Development: The Case Of The "Baja Tarahumara" in Chihuahua, Mexico
- Ecological and Socio-Economic Impacts of Chromolaena odorata and Mesosphaerum suaveolens, Two Invasive Alien Species in Central and Southern Benin, West Africa
- Epigenetic Sustainability: Modeling the Human Factor as a Natural Resource through Science 4.0 and the NR3C1 Biological Pilot
- Growth-at-Risk: A Framework for Assessing Economic Vulnerability
- The Rural Territory as a Socioecological System for the Management of Public Policy for Sustainable Rural Development