Epigenetic Regulation of Chromatin during Mitosis in Embryos
Recent studies show that mitosis does not involve complete eradication of the past chromatin states but the memory of active and repressed chromatin is maintained to the next generation. This epigenetic regulation is critical for embryonic development and is also required for normal physiological functions. Further, impairment in this gene regulation can result in disease conditions such as cancer. Yet, the mechanisms by which regulatory components remain bound to the compacted mitotic chromosomes is not clearly understood. Here we review the process of gene bookmarking during mitosis in the developing embryo and its impact on the transcriptional dynamics during embryogenesis. The phenomenon of bookmarking is pivotal to normal embryonic development as it ensures that the transcriptional program is faithfully inherited to maintain cell fates. We review the mechanisms which control the accessibility of regulatory regions in the genome during cell division in the developing embryo. These include regulation by transcription factors, chromatin regulators and epigenetic histone marks such as histone tail modifications. For example, the Polycomb group of genes (PcG) regulate long term memory of the repressed chromatin state in the Drosophila embryo. We will also deliberate on the latest tools and techniques available to study the phenomenon of gene bookmarking such as ATAC seq and chromosome conformation capture technique. Finally, we will also discuss some of the unanswered questions and possible future studies in this field.
Introduction
Cell division (mitosis) is one of the underlying fundamental processes which govern embryogenesis. In this article, we review the chromatin landscape and chromatin state during mitosis in the developing embryo. The status of chromatin during mitosis in the embryonic context is not well understood. We discuss the concept of mitotic memory, essentially defined as the memory of active and repressed chromatin states that are maintained between successive mitotic divisions of mother to daughter cells in the embryo. Thus, the regulatory machinery of the mother cell is stably inherited to the progeny. We aim to highlight these epigenetic memory mechanisms which regulate gene expression states through multiple cell generations without alterations in the DNA sequence and in the absence of the initiating signaling cues.
Chromatin and Mitosis
During mitosis, the chromatin undergoes drastic changes and is subjected to spatial constraints due to the apparent nuclear reorganization. At the onset of mitosis, chromatin proteins undergo phosphorylation which results in condensation of the chromatin [1, 2, 3]. The nuclear envelop is broken down, the chromatin is reorganized into compact, rod shaped chromosomes. In-vitro data shows that phosphorylation of condensing is required for the chromatin compaction. The in vivo experiment also suggests some other components are required in the process. Recent study shows role of histone H3 phosphorylation in recruitment of condensing complexes to chromatin. In early mitosis histone H3 phosphorylation by Haspin and Aurora B but the association with chromatin is lost in mid mitosis. The exact role of histone H3 phosphorylation in chromatin condensation is not fully understood [4]. Several cellular mechanisms then ensure the inactivation of many transcription factors and bring about a significant down regulation of transcription by mechanisms like targeted phosphorylation [3, 5].
Following faithful chromosome segregation, nuclear envelop is formed and bulk transcription resumes in the daughter cells. However, the transcriptional properties of the dividing embryonic cells have to be maintained across multiple divisions in order to maintain cell fate identity and lineage commitment. Classical hypothesis suggest that chromosomes separate from the transcriptional machinery and silence gene expression during mitosis. Recent studies show that certain cell type specific transcriptional activities persist throughout mitosis. For example, the transcription factor GATA1 remains associated with the chromosomes at mitosis in the erythroid cells and aids in the transmission of the transcriptional program through the mitotic event [6, 7, 8]. Further, epigenetic factors such as Polycomb group (PcG) and Trithorax group (TrxG) also remain associated with the DNA during all phases of mitosis. The PcG proteins provide epigenetic memory of silent gene expression states whereas the TrxG proteins provide memory for activated gene expression state. The histone methylation pattern by PcG complex is associated with closed configuration of chromatin. In contrast, the TrxG complexes are associated with open chromatin configuration. Both these associations persist through mitosis, further suggesting the bookmarking function of these factors [9, 10].
What is Mitotic Memory?
Mitotic bookmarking refers to the phenomenon where certain transcriptional regulators are retained inside the condensed chromatin and directly instruct gene reactivation once mitosis is complete. At the end of mitosis, the pattern and amplitude of the gene expression is re-established [11]. The knockdown of these bookmarking genes during mitosis results in impaired transcription reactivation [2]. According to one model of epigenetic control of gene expression, TFs which undergo histone modifications bind their target genes upon the mitotic exit, and induce gene expression.
Another hypothesis is that certain TFs bind to the mitotic chromosomes, act as bookmarks during mitosis and enable gene expression upon completion of mitosis. During embryogenesis, many developmental regulators have been shown to interact with mitotic chromatin.
How is Mitotic Memory Achieved?
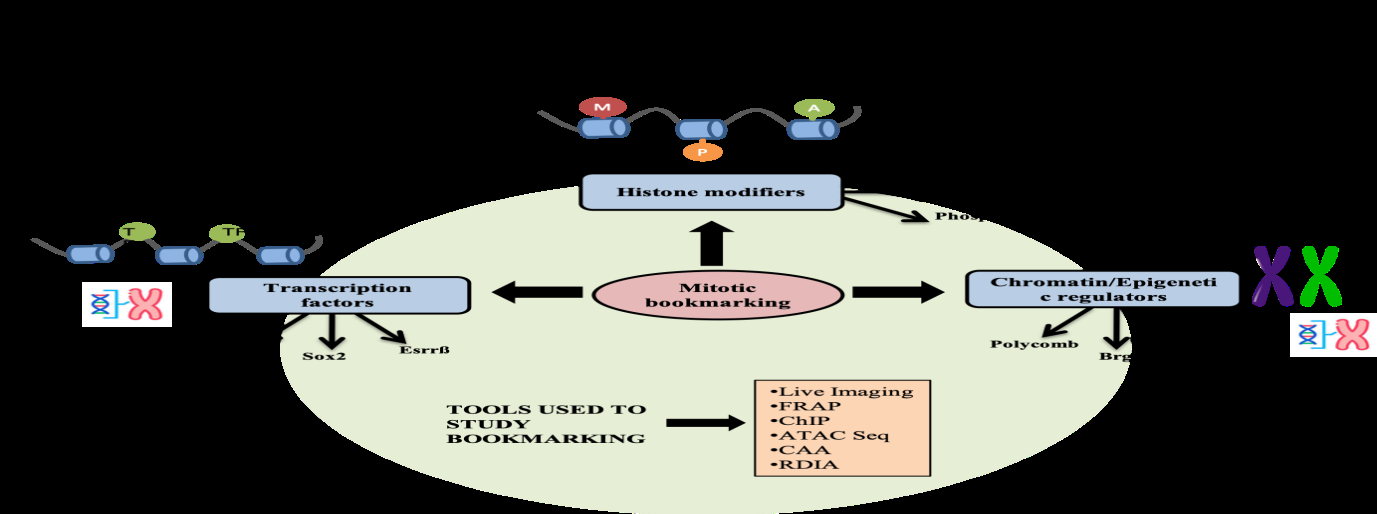
Mitotic memory is primarily achieved by three mechanisms. These include the retention of general and sequence specific transcription factors (TFs), chromatin regulators and epigenetic histone marks such as histone tail modifications. As discussed, a mitotic cell undergoes significant down regulation of overall transcription, primarily mediated by targeted phosphorylation and inactivation of several TFs. The sequence specific TFs like myogenic factor Myf5, Oct4, Sox2 and other chromatin remodellers like Brg1 are hyperphosphorylated, leading to their degradation or inactivation. However, the complete molecular mechanism of phosphorylation mediated inactivation of TFs during embryogenesis is not completely understood [3, 12]. Histone modifications too, function as landmarks to provide memory after the cell divides and also facilitate the interaction of other TFs and regulatory proteins. Some of these modifications include H3-H4 methylation, H3-K79 methylation and H3 acetylation. All these modifications are stably inherited during mitosis [2]. Mitosis is thus, associated with histone phosphorylation and the regulatory regions of the DNA maintain high levels of H3K27 acetylation. The Histone Acetylase Transferase (HAT) p300 remains bound to the mitotic chromatin, indicating its role in maintenance of DNA accessibility [3].
Mitotic Memory in the Embryo
During embryogenesis, rapid cell division is required for embryonic growth and generation of different cell lineages. Lineage specification is coupled to differentiation and is primarily brought about by transcription factors, which establish cell type specific transcription patterns of developmental genes. As cells exit mitosis, the cell type specific gene expression patterns needs to be faithfully and rapidly restored in the daughter cells. In fact, the acquisition of cell identity is initiated during the cell division process itself. The re-establishment of gene expression is brought about by a combination of epigenetic factors like histone modifications and TFs which are retained on the mitotic chromosomes, termed as mitotic bookmarking. Cell fate commitment during development is determined by regulation of transcription factor and epigenetic modifiers. The epigenetic memory of cell fate decision is achieved by mitotic bookmarking factors and histone modifications [13]. The inheritance of repressed and active chromatin states are result of histone modifications, including acetylation, methylation, phosphorylation, and ubiquitination. Histone methylation such as H3K27me3 and H3K9me3 are hallmarks of repressive chromatin. H3K27me3 and Polycomb repressor complexes control the epigenetic memory of cell fate determination. Whereas trimethylation of H3K4 catalyzed by trithorax-group (TrxG) protein complexes activate repressed genes. Some of the TFs, including GATA1, FOXA1, OCT4, SOX2, ESRRB, KLF4, and the TATA-binding protein and called mitotic bookmarkers because during mitosis they remain associated to the chromatin by nonspecific DNA binding (Figures 1 & 2) [13].
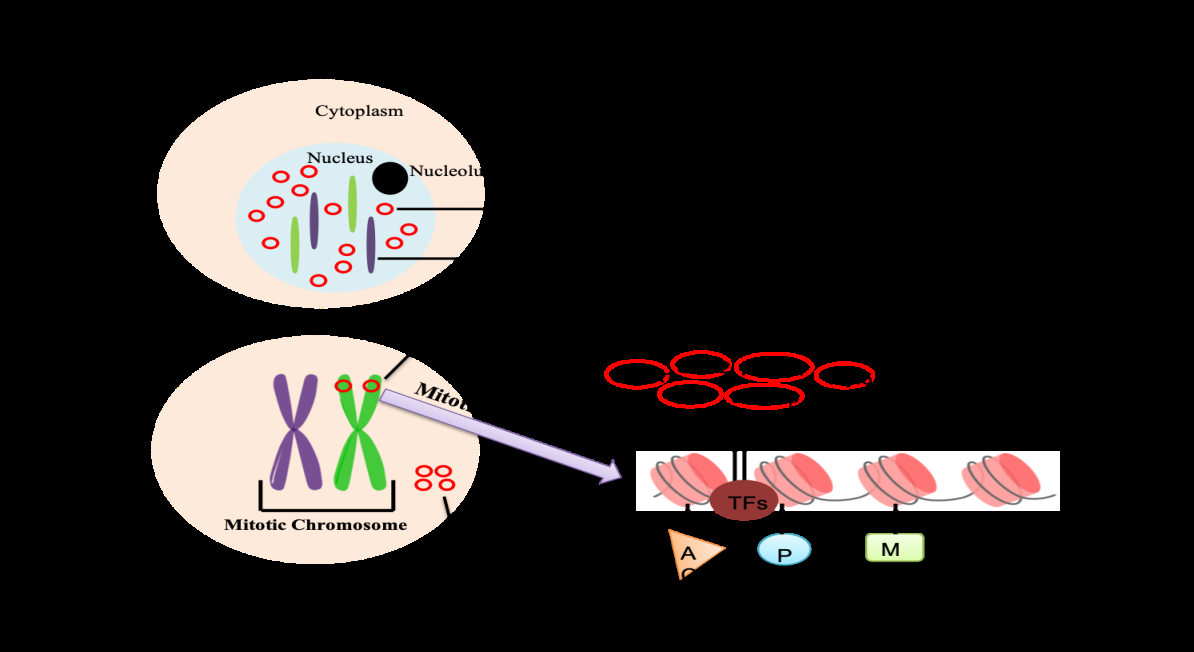
This mitotic memory serves an important function in the developing embryo. Key regulatory proteins involved in proliferation, growth or differentiation are found to remain associated with the target gene loci on the mitotic chromosomes. For example, the RUNX family of proteins which regulate osteogenesis and hematopoeisis, associate with the ribosomal and cell cycle regulatory genes like p21 during mitosis [14, 15, 16]. This association on the mitotic chromosomes results in repression of these regulatory genes during G1 phase [2]. Another myogenic regulatory factor myoblast determination protein (MYOD), localizes to the mitotic chromosomes and associates with the ribosomal RNA genes during cell division of mesenchymal cells. With the onset of the differentiation to form skeletal muscle, MyoD is replaced by myogenin, resulting in repression of the ribosomal RNA genes [14]. Another TF called FOXA1, required for liver development is another bookmarking factor binds to mitotic chromosomes and to about 15% of its interphase targets during mitosis [17, 18]. TF termed as ESRRβ which plays key role in maintenance of pluripotency in embryonic stem cells, binds preferentially to genes associated with G1 phase, thus aiding to reactivate these genes after mitosis is complete [19, 20, 21, 22]. During hematopoiesis, TF termed as GATA1 which is required to establish lineage specific expression binds to the mitotic chromatin and its target genes are reactivated upon entry into interphase. This results in the faithful transfer of hematopoietic gene expression program to the resultant daughter cells. Thus, the active transcriptional states are inherited which ensure the fidelity of the transcriptional programs during each mitotic division. It also imparts stability, flexibility and or plasticity to the developmental program. For example, the demethylation of H3K4 aids in the reprogramming in Xenopus embryos [23].
Besides embryonic development, mitotic bookmarking plays a key role in onset and progression of many diseases such as leukemia. The chromatin remodelling factor called mixed lineage leukemia protein (MLL) is associated with leukemia. This factor regulates transcription by recruitment of chromatin modifying factors to the target genes and functions as a mitotic bookmarking factor. MLL is retained to the target genes during mitosis, which enables the post- mitotic transcriptional activation of target genes [23, 24]. Another leukemia associated protein called leukemic fusion protein AML1-ETO (RUNX1T1) which bookmarks rRNA genes during mitosis. It also bookmarks genes which regulate myeloid cell differentiation and cell proliferation genes [24, 25, 26].
Tools to Study Gene Bookmarking
The bookmarking factors typically localize to the mitotic chromosomes. The classical fixation methods and immunofluorescence methods tend to disrupt the interaction of TFs with mitotic chromosomes [8, 12]. Thus, visualization by live imaging of fluorescent fusion proteins, expressed from the endogenous loci on the chromosomes is the preferred technique to study mitotic chromosome binding [1, 3, 12]. The dynamic interaction of the bookmarking factors with the chromatin can also be studied using Fluorescence recovery after photobleaching (FRAP) and single particle tracking assays [1, 8, 11]. Further, their site-specific binding to the regulatory elements further reinforces their bookmarking function. However, in cases where live imaging approach is not feasible, ChIP-seq or Assay for Transposase-Accessible Chromatin using sequencing (ATAC seq) are performed to establish bookmarking activity [1, 8, 25]. Chromatin Association Assay (CAA) and RNA DNA Interaction Assay (RDIA) have also been developed to detect DNA interactions with chromatin remodelling proteins like PcG and TrxG during mitosis in Drosophila embryos [25]. Other techniques include motif foot-printing using DNase, pulse labeling of the nascent RNA, chromosome conformation capture techniques, proximal ligation assay and mathematical modelling tools such as Markov chain method [3]. Multiple experimental approaches thus, provide corroborating evidence for bookmarking function.
Conclusion and Future Prospects
The mechanism of epigenetic inheritance through mitosis is crucial to maintain cell identity and pave the way for lineage commitment. However, all genes are not bookmarked during mitosis and the underlying cause of this selective phenomenon is still unknown. To conclude, it is more likely that multiple epigenetic mechanisms regulate a gene which is mitotically bookmarked. Thus, many unanswered questions need to be addressed in the field of mitotic bookmarking. These include the mechanism by which TFs resume their binding and regulation of their target genes post mitosis and how TFs interact with the mitotic chromosomes. Future studies to elucidate the molecular crosstalk between mitotic transcription and structural features present on the chromosomes will provide valuable insights into the epigenetic control to retain cellular identity after mitosis. Technological advances such as high-resolution live imaging and genome wide transcriptome and proteome analysis will aid in answering some of these pertinent questions of gene bookmarking. Lastly, the impact of gene bookmarking to the onset and progression of diseases such as cancer and to develop novel therapeutic strategies or targeted therapy also need to be addressed in the future. The molecular mechanisms uncovering how the cells remember their ‘past’ will prove instrumental to design better reprogramming strategies for gene therapy.
Credit authorship contribution statement
SM, SI, TP contributed equally in manuscript writing and figure preparation. MK developed the concept of the article, prepared the figures and wrote the manuscript.
Declaration of competing interest
The authors have no competing interests.
Acknowledgments
The authors thank the Council of Scientific and Industrial Research (CSIR), Govt. of India and Department of Science and Technology, Govt. of India (DST/INSPIRE/04/2016/001436) for support and funding this research.
References
-
Kadauke S, Blobel GA (2013) Mitotic bookmarking by transcription factors. Epigenetics & Chromatin 6(1): 1-10.
-
Lodhi N, Ji Y, Tulin A (2016) Mitotic bookmarking: maintaining post-mitotic reprogramming of transcription reactivation. Curr Mol Biol Rep 2(1): 10- 16.
-
Festuccia N, Gonzalez I, Owens N, Navarro P (2017) Mitotic bookmarking in development and stem cells. Development 144(20): 3633-3645.
-
van Wely K, Mora G, Vann K, Kutateladze T (2017) Epigenetic countermarks in mitotic chromosome condensation. Nucleus 8(2): 144-149.
-
John S, Workman JL (1998) Bookmarking genes for activation in condensed mitotic chromosomes. Bioessays 20(4): 275-279.
-
Zaidi SK, Young DW, Montecino M, van Wijnen AJ, Stein JL, et al. (2011) Bookmarking the genome: maintenance of epigenetic information. J Biol Chem 286(21): 18355- 18361.
-
Iglesias AR (2013) Pioneering barren land: mitotic bookmarking by transcription factors. Dev Cell 24(4): 342-344.
-
Raccaud M, Suter D (2018) Transcription factor retention on mitotic chromosomes: regulatory mechanisms and impact on cell fate decisions. FEBS Lett 592(6): 878-887.
-
Sarge KD, Park Sarge OK (2005) Gene bookmarking: keeping the pages open. Trends Biochem Sci 30(11): 605-610.
-
Steffen P, Ringrose L (2014) What are memories made of? How Polycomb and Trithorax proteins mediate epigenetic memory. Nat Rev Mol Cell Biol 15(5): 340- 356.
-
Palozola K, Lerner J, Zaret K (2019) A changing paradigm of transcriptional memory propagation through mitosis. Nat Rev Mol Cell Biol 20(1): 55-64.
-
Teves SS, An L, Hansen AS, Xie L, Darzacq X, et al. (2016) A dynamic mode of mitotic bookmarking by transcription factors. Elife 5: 22280.
-
Elsherbiny A, Dobreva G (2021) Epigenetic memory of cell fate commitment. Current opinion in cell biology 69: 80-87.
-
Ziadi S, Young W, Montecino M, Lian J, Wijnen A, et al. (2010) Mitotic bookmarking of genes: a novel dimension to epigenetic control. Nat Rev Genet 11(8): 583-589.
-
Zaidi SK, Van Wijnen AJ, Lian JB, Stein JL, Stein GS (2013) Targeting deregulated epigenetic control in cancer. J Cell Physiol 228(11): 2103-2108.
-
Rose JT, Moskovitz E, Boyd JR, Gordon JA, Bouffard NA, et al. (2020) Inhibition of the RUNX1-CBFβ transcription factor complex compromises mammary epithelial cell identity: a phenotype potentially stabilized by mitotic gene bookmarking. Oncotarget, 11(26): 2512-2530.
-
Caravaca JM, Donahue G, Becker JS, He X, Vinson C, et al. (2013) Bookmarking by specific and nonspecific binding of FoxA1 pioneer factor to mitotic chromosomes. Genes Dev 27(3): 251-260.
-
Zaret K (2014) Genome reactivation after the silence in mitosis: recapitulating mechanisms of development?. Dev Cell 29(2): 132-134.
-
Festuccia N, Dubois A, Pournin SV, Tejeda EG, Mouren A, et al. (2016) Mitotic binding of Esrrb marks key regulatory regions of the pluripotency network. Nat Cell Biol 18(11): 1139-1148.
-
Hsiung CC, Blobel GA (2016) A new bookmark of the mitotic genome in embryonic stem cells. Nat Cell Biol 18(11): 1124-1125.
-
Festuccia N, Owens N, Papadopoulou T, Gonzalez I, Tachtsidi A, et al. (2019) Transcription factor activity and nucleosome organization in mitosis. Genome Res 29(2): 250-260.
-
Coux RX, Owens N, Navarro P (2020) Chromatin accessibility and transcription factor binding through the perspective of mitosis. Transcription 11(5): 236-240.
-
Zaidi SK, Nickerson JA, Imbalzano AN, Lian JB, Stein JL, et al. (2018) Mitotic Gene Bookmarking: An Epigenetic Program to Maintain Normal and Cancer Phenotypes. Mol Cancer Res 16(11): 1617-1624.
-
Zaidi SK, Lian JB, van Wijnen A, Stein JL, Stein GS (2017) Mitotic Gene Bookmarking: An Epigenetic Mechanism for Coordination of Lineage Commitment, Cell Identity and Cell Growth. Adv Exp Med Biol 962: 95-102.
-
Black K, Petruk S, Fenstermaker T, Hodgson J, Caplan J, et al. (2016) Chromatin proteins and RNA are associated with DNA during all phases of mitosis. Cell Discovery 2: 16038.
-
Bellec M, Radulescu O, Lagha M (2018) Remembering the past: Mitotic bookmarking in a developing embryo. Curr Opin Syst Biol 11: 41-49.
- Research Progress of Induced Pluripotent Stem Cells and Their Clinical Application Prospects
- Nishan Al-Kamal is the Starting Point of A Feminist Scientist
- Current Concepts and Future Perspectives of Stem Cell Therapy in Peripheral Arterial Disease
- Stem Cell and Oxidative Stress-Inflammation Cycle
- Adipose Derived Mesenchymal Stem Cells Origin, Characteristics and Promises
- Mitochondria Targeted Antioxidants can Improve In Vitro Embryo Production in Buffalo