Phenotypic Variation in Two Wild Radishes: Raphanus pugioniformis and Raphanus raphanistrum in Field and Common Gardens in Palestine
A study was conducted to explore phenotypic variation in two Raphanus species: Raphanus raphanistrum and R. pugioniformis, which are found in the East-Mediterranean region. For each species, a 100m2 area representing their respective habitats was selected. Morphological characterization of the whole plant, leaves, and fruits was carried out in the field. The study analyzed morphological characteristics of R. pugioniformis populations during both flowering and maturity stages. It also compared the maturity stage characteristics. The examined characteristics included plant height, flower to plant ratio, leaf to plant ratio, leaf length, flower color, tip to pod ratio, seed to fruit ratio, fruit parameters (length, width, and weight), and the number of primary and secondary branches. Results revealed that plant height, tip to pod ratio, and fruit length increased from the flowering stage to the maturity stage. However, the number of primary and secondary branches and the ratio of seeds to fruit remained consistent between the flowering and maturity stages. R. raphanistrum exhibited greater plant height, flower to plant ratio, leaf to plant ratio, tip to pod ratio, seed to fruit ratio, fruit length, fruit width, fruit weight, and number of branches. The study examined also the dry weight and nitrogen, phosphorous, and potassium contents in leaves, stems, and seeds. It was found that R. pugioniformis had higher nitrogen and phosphorous contents in the leaves, stem, seeds, and whole plant compared to R. raphanistrum. R. pugioniformis exhibited higher nitrogen and phosphorous contents in the leaves, seeds, and whole plant. Similar trends were observed for potassium content. In summary, the study highlighted the phenotypic variation between R. raphanistrum and R. pugioniformis populations in the East-Mediterranean region. Differences were observed in various morphological characteristics, dry weight, and nutrient contents, providing valuable insights into the contrasting traits and adaptations of these closely related Raphanus species.
Introduction
Wild radish, belonging to the Raphanus genus, is a notable member of the Brassicaceae family, which is one of the four prominent families encompassing weeds and crops. This species has gained recognition as an emerging model for investigating rapid adaptation and weed evolution [1, 2, 3, 4]. Investigating the origins of weedy radish is relevant due to the unique characteristics of the Raphanus genus. This genus comprises only three species, all of which display self- incompatibility. Native populations of the Raphanus genus are exclusively found in the Mediterranean region, making it an important area to study the origin and distribution of these species [5].
R. pugioniformis, an endemic species of the eastern Mediterranean [6], has received limited research attention. The relationships among these species remain largely unresolved [7]. However, a cDNA sequence analysis conducted on one population from each of eight Raphanus taxa (excluding R. pugioniformis) provided compelling evidence supporting the monophyly of the crop cultivars and the native and weedy forms of R. r. raphanistrum [2].
Weedy _R. r. raphanis_trum is frequently found as a troublesome invader in small grain fields and is widely recognized as one of the most significant agricultural weeds globally [7, 8, 9]. Weedy radish has been extensively utilized in ecological and evolutionary research, specifically for investigating plant-insect interactions [10, 11, 12, 13, 14, 15, 16], and possesses valuable genomic resources that render it an ideal system for studying the origins and adaptations of agricultural weeds, enabling researchers to address pertinent questions in this field [17].
Raphanus raphanistrum and R. pugioniformis, two closely related annual plants from the Brassicaceae family, with spatial isolation (allopatry) between them. In Palestine, R. raphanistrum populations primarily thrive on sandy soils along the Mediterranean coastal plain, while R. pugioniformis is endemic to the southeast Mediterranean, occupying diverse habitats across varying environmental conditions [6].
The differences in seed dispersal, with R. raphanistrum dispersing over long distances and R. pugioniformis dispersing over shorter distances, have an impact on their population spatial structure [7].
Consequently, it is anticipated that these species exhibit dissimilarities in their population genetic structure.
Environmental conditions contribute to the diversity observed in the genetic and physical characteristics of plants in the region [18, 19, 20]. To understand the relationship between phenotypic variation and species distribution, it is essential to investigate the potential impact of environmental conditions. This influence of environmental factors on ecotypic differentiation can be expected not only in locally restricted species but also in widely distributed ones, regardless of possible gene flow between populations. This phenomenon has been observed in Sarcopoterium spinosum, a perennial shrub belonging to the Rosaceae family, where environmental factors have been reported to contribute to ecotypic differentiation despite its wide distribution [21]. The distinct dispersal strategies employed by wild radishes in the southeast Mediterranean provide a unique opportunity to examine the influence of environmental conditions on the phenotypic variation of ecologically significant life history traits in annual plants.
The successful adaptive radiation of the Brassicaceae (cabbage family) in exploiting newly available ecological niches can be primarily attributed to whole-genome duplications. This, coupled with the development of novel morphological and physiological traits, has allowed for effective adaptation to diverse environmental conditions. These adaptations are associated with shifts in pollination ecology, defense strategies, dispersal mechanisms, fruit structure and seed traits, germination patterns, and transitions from perennial to annual growth strategies [20, 21]. Despite the importance of the aforementioned traits in adaptive radiation, our understanding of their precise adaptive significance remains limited due to a lack of comprehensive experimental and observational studies. Furthermore, it is crucial to emphasize the examination of trait divergence in homologous organs within closely related species when investigating adaptive characteristics from an evolutionary perspective. By doing so, we can gain deeper insights into the evolutionary mechanisms driving trait variations and their adaptive implications.
In this study, our aim is to conduct an evolutionary- ecological investigation to unravel the importance of variations in fruit structure and function, as well as the genetic structure of populations, in two allopatric and closely related Raphanus species found in the East-Mediterranean region: Raphanus raphanistrum and R. pugioniformis. We propose the hypothesis that differences in fruit structure and dispersal mechanisms between these two species have evolved as adaptive traits specifically suited to their respective habitats. To test this hypothesis, we have outlined three primary objectives: Explore the ecological significance of fruit variation and dispersal units, investigate the interplay between diaspore dispersal and germination patterns, and analyze the patterns of genetic variation within and among populations of Raphanus. By addressing these objectives, we aim to gain a deeper understanding of how variations in fruit traits, as well as dispersal and genetic patterns, contribute to the adaptive strategies of these Raphanus species in their respective environments.
The study analyzed morphological characteristics of R. pugioniformis populations during both flowering and maturity stages. It also compared the maturity stage characteristics. The examined characteristics included plant height, flower to plant ratio, leaf to plant ratio, leaf length, flower color, tip to pod ratio, seed to fruit ratio, fruit parameters (length, width, and weight), and the number of primary and secondary branches.
Methodology
Mapping of Wild Radish Populations
In 2021, a survey was carried out in the Jenin district to investigate the presence, distribution, and phenotypic characteristics of wild radish from the Raphanus genus. The study area revealed the occurrence of two allopatric species of Raphanus in separate locations. Clustered populations of Raphanus pugioniformis were identified in Faqua (eastern slope, coordinates: 32.4778° N latitude, 35.3987° E longitude), while clustered populations of Raphanus raphanistrum were found in Yabad (western slope: 32.4594° N latitude and 35.0708° E longitude). For the study, populations of Raphanus were carefully selected to represent approximately 100m2 growing areas within the natural habitat of Raphanus raphanistrum and Raphanus pugioniformis. Randomly chosen populations of five for each species (RR and RP) underwent morphological characterization in the field, encompassing the whole plant, leaves, and fruits.
Common Garden Experiment
Phenotypic Evaluation Using Seeds Collected in Nature: The phenotypic evaluation took place within a controlled net house environment during the natural growing season of the species, spanning from November to March 2021. The average day maximum and minimum temperatures during this period were recorded as 20°C and 14°C, respectively. For germination, seeds were placed on moistened Whatman No. 1 filter paper in 9cm Petri dishes within a growth chamber set at a temperature of 25°C and an 8/16 hour day/night photoperiod. After four days, the seedlings were transferred to germination trays and placed in the net house. Two weeks later, the plants were transplanted into 1L pots filled with a mixture comprising 50% peat, 30% tuff, and 20% perlite. The phenotypic evaluation encompassed several traits, including the bolting date, rosette diameter (measured as the distance between the tips of opposite leaves) at the time of bolting, stalk length at the time of flowering, as well as overall plant size and stalk length.
Plant Harvest and Mineral Analysis: At two different stages, namely flowering and maturity, various measurements were taken, including plant height, number of primary and secondary branches, number of flowers (during the flowering stage), and number of pods. For each population, nine whole plants were individually harvested at seed physiological maturity. They were then placed in sealed paper bags and transported to the laboratory for further analysis.
Upon harvest, the dry weight of the plant shoot was determined by drying the samples at a constant temperature of 70°C. The seeds were carefully separated from the pericarps and weighed, taking into account the total weight as well as the thousand seed weight. The dried plant materials were ground to pass through a 1.5 mm sieve, and a sub-sample of 5 g was further processed to a fine powder using a ball mill. Seed samples were ground after being soaked in liquid nitrogen.
For the analysis of phosphorus (P) and potassium (K), the plant samples were prepared through dry ashing. Total nitrogen (N) analysis was conducted using the Kjeldahl method. The spectrophotometric methods were employed to measure the total P and N contents in the digested plant material, while a Flame Photometer was used to determine the total K content in the plant material digest.
Results
For each of the two Raphanus species (RR and RP), five populations were randomly chosen for morphological characterization. The evaluation encompassed the entire plant, leaves, and fruits, and included measurements of plant height, number of primary and secondary branches, and number of flowers during the flowering stage, and number of pods. These measurements were taken at two different stages, namely flowering and maturity.
Morphological Characteristics of R. pugioniformis at Maturity and Flowering Stages
At seed physiological maturity, nine whole plants were individually harvested from each population. The following morphological parameters were measured: plant height, fruit length, tip/pod ratio, number of seeds to fruit ratio, and number of primary and secondary branches. These measurements were conducted on three populations of R. pugioniformis in their maturity stage in Faqua during May 2021, and the results are summarized in Table 1.
The plant height and fruit length of the R. pugioniformis populations ranged from 142.0 to 146.7 cm and from 2.1 to 3.3 mm, respectively. Population 2 exhibited the highest plant height and fruit length, while population 3 had the lowest values. The tip to pod ratio varied from 1.1 for population 3 to 1.5 for population 1. In terms of the ratio of seeds to fruit, populations 1 and 3 had similar values (ratio of 3.3), while population 2 had a higher ratio of 4.0.
The number of primary branches ranged from 8.7 to
11.7, and the number of secondary branches ranged from 16.3 to 28.0 across the three R. pugioniformis populations. It was observed that the number of primary branches was comparable among the three populations, while significant differences were observed in the number of secondary branches (Table 1).
| Trait | Population 1 | Population 2 | Population 3 |
|---|---|---|---|
| Plant height (cm) | 142.3±2.1 | 146.7±3.1 | 142.0±1.7 |
| Flowers per plant | 0.0±0.0 | 0.0±0.0 | 0.0±0.0 |
| Fruit length (mm) | 2.7±0.3 | 3.3±0.2 | 2.1±0.2 |
| tip/pod ratio | 1.5±0.0 | 1.4±0.3 | 1.1±0.2 |
| Number of seeds/fruit | 3.3±0.6 | 4.0±0.0 | 3.3±1.2 |
| Primary branches | 11.7±1.5 | 8.7±0.6 | 11.0±1.0 |
| Secondary branches | 22.3±2.1 | 28.0±1.7 | 16.3±1.5 |
Table 1: Morphological Characterization (Average ± Standard Deviation) of R. pugioniformis in its Maturity Stage (May 2021) in Fa
Additionally, the morphological characteristics of three populations of R. pugioniformis were examined during its flowering stage in Faqua (March 2021), as presented in Table 2. The following parameters were measured: plant height, flowers per plant, flower color, fruit length, tip/pod ratio, number of seeds to fruit ratio, and number of primary and secondary branches.
The plant height of the R. pugioniformis populations during the flowering stage ranged from 76.7 to 86.7 cm, with a fruit length range of 1.1 to 1.2 mm. Population 3 exhibited the tallest plant height, while population 2 had the lowest values. The fruit length was comparable among the three populations during the flowering stage. When comparing the plant height and fruit length of R. pugioniformis between the flowering and maturity stages, it was observed that the measurements at the maturity stage were significantly higher, approximately twice the values observed during the flowering stage (Tables 1 & 2).
The number of flowers per plant in the R. pugioniformis populations during the flowering stage ranged from 85.0 (population 3) to 106.7 (population 1). As expected, no flowers were observed during the maturity stage of R. pugioniformis (Table 1). The flower color for all three populations of R. pugioniformis was purple. The tip to pod ratio was consistent at 1.0 for all three populations during the flowering stage.
Comparing the ratios between the flowering and maturity stages, it was found that the ratio at the maturity stage (ranging from 1.1 to 1.5) was higher than that during the flowering stage of R. pugioniformis. The ratio of the number of seeds to fruit was similar between populations 1 and 3 (ratio of 3.3), while population 2 had a higher ratio of 4.0. These ratio values during the flowering stage remained consistent during the maturity stage (Tables 1 & 2).
| Population 1 | Population 2 | Population 3 | |
|---|---|---|---|
| Plant height (cm) | 84.7±4.5 | 76.7±2.5 | 86.7±3.2 |
| Flowers per plant | 106.7±7.6 | 97.3±6.7 | 85.0±4.4 |
| Fruit length (mm) | 1.2±0.0 | 1.1±0.2 | 1.2±0.2 |
| Flower color | Purple | Purple | Purple |
| tip/pod ratio | 1.0±0.0 | 1.0±0.0 | 1.0±0.0 |
| Number of seeds/fruit | 3.3±0.6 | 4.0±0.0 | 3.3±1.2 |
| Primary branches | 11.7±1.5 | 8.7±0.6 | 11.0±1.0 |
| Secondary branches | 22.3±2.1 | 28.0±1.7 | 16.3±1.5 |
Table 2: Morphological Characterization (Average ± Standard Deviation) of R. Pugioniformis in its Flowering Stage (March 2021) in
The number of primary and secondary branches for the three populations of R. pugioniformis during the flowering stage ranged from 8.7 to 11.7 and from 16.3 to 28.0, respectively. Comparing the data in Tables 1 and 2, it can be observed that the number of primary and secondary branches remained the same between the flowering and maturity stages (Tables 1 & 2).
Table 3 presents the survey data collected for natural populations of R. pugioniformis during its flowering stage in Faqua (March 2021). The data includes the area of the populations (20 and 30 m2), the dispersion pattern of the populations (clustered), plant density (7 plants/m2), total vegetation cover (ranging from 85% to 95%), slope gradient (15 and 30 cm/m), rockiness or rock cover (20%), and potential pollinators (none observed) in all three populations.
| Population 1 | Population 2 | Population 3 | |
|---|---|---|---|
| Area of the population (m2) | 20 | 30 | 30 |
| Population dispersion | clustered | clustered | clustered |
| Plant density (plant/m2) | 7 | 7 | 7 |
| Total vegetation cover | 90% | 95% | 85% |
| Slope (cm/m) | 30 | 30 | 15 |
| Rockiness (rock cover %) | 20% | 20% | 20% |
| Potential pollinators | No pollinators | No pollinators | No pollinators |
Table 3: Survey, Data Collection for Natural Populations of R. Pugioniformis in its Flowering Stage (March 2021) in Faquoa.
Morphological Characteristics R. pugioniformis and R. raphanistrum in their Maturity Stages
Table 4 presents the measured morphological characteristics of three populations each of R. pugioniformis and R. raphanistrum during their maturity stages in the common garden (May 2021). The parameters assessed include plant height, flowers to plant ratio, leaves to plant ratio, leaf length, flower color, tip to pod ratio, ratio of the number of seeds to number of fruits, fruit parameters (length, width, and weight), and the number of primary and secondary branches.
Results indicate that R. raphanistrum generally exhibits higher plant height compared to R. pugioniformis (102.0 cm vs. 80.4 cm). The flower to plant ratio and leaves to plant ratio were also higher for R. raphanistrum, as shown in Table 4. Leaf length was found to be 44.0 cm for R. raphanistrum and 37.2 cm for R. pugioniformis. Furthermore, the flowers of the two Raphanus species differed in color, with R. raphanistrum displaying white flowers and R. pugioniformis showcasing purple flowers. The tip to pod ratio for R. raphanistrum was twice that of R. pugioniformis. Similarly, the ratio of the number of seeds to fruits was higher for R. raphanistrum (5.6) compared to R. pugioniformis (4.2). In terms of fruit parameters, R. raphanistrum exhibited longer, wider, and heavier fruits compared to R. pugioniformis, as depicted in Table 4. The number of primary and secondary branches also followed the same trend, with R. raphanistrum displaying higher values compared to R. pugioniformis (12.8 vs. 9.6 for primary branches and 30.6 vs. 24.4 for secondary branches).
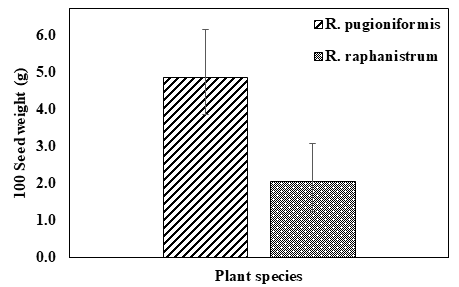
Additionally, the weight of 100 seeds at their maturity stages was measured for both R. pugioniformis and R. raphanistrum. The results demonstrate that R. pugioniformis had a higher seed weight (4.9 g) compared to R. raphanistrum (2.1 g), as illustrated in Figure 1.
| R. pugioniformis | R. raphanistrum | |
|---|---|---|
| Plant height | 80.4±5.8 | 102.0±9.2 |
| Flower/ plant | 96.0±9.3 | 104.0±8.7 |
| Leaves/plant | 129.0±3.1 | 148.4±7.8 |
| Leaf length | 37.2±3.0 | 44.0±3.2 |
| Flower color | purple | white |
| Tip/pod ratio | 1.0±0.0 | 2.0±0.0 |
| Number of seeds /fruit | 4.2±0.4 | 5.6±1.1 |
| Fruit length (mm) | 10.8±1.8 | 14.4±1.7 |
| Fruit width (mm) | 0.5±0.1 | 0.6±0.1 |
| Fruit weight (gm) | 0.7±0.4 | 1.2±0.2 |
| Primary branches | 9.6±1.5 | 12.8±2.6 |
| Secondary branches | 24.4±5.8 | 30.6±4.5 |
Table 4: Morphological Characterization (Average ± Standard Deviation) of R. pugioniformis and R. raphanistrum in its Maturity St
Dry Weight and Mineral Analysis of the Two Raphanus Species
Table 5 provides the measurements of dry weight and nutrient contents (N, P, and K) in different plant parts (leaves, stems, seeds, and whole plants) of R. pugioniformis and R. raphanistrum during their maturity stage in the common garden (May 2021).
The dry weight of R. pugioniformis leaves was approximately double that of R. raphanistrum (11.5 compared to 6.7). Similarly, higher dry weights were observed in R. pugioniformis seeds, while the reverse was true for the dry weight of the stem, where R. raphanistrum had a higher value compared to R. pugioniformis. However, the dry weights of the whole plants were comparable for both R. pugioniformis and R. raphanistrum.
| Dry weight (g) | N content (mg/g) | P content (mg/g) | K content (mg/g) | |
|---|---|---|---|---|
| Leaves | ||||
| R. pugioniformis | 11.5±2.4 | 34.5±9.4 | 6.4±1.3 | 11.7±2.7 |
| R. raphanistrum | 6.7±3.0 | 32.0±7.1 | 5.9±1.3 | 11.9±2.2 |
| Stem | ||||
| R. pugioniformis | 21.6±3.7 | 21.8±5.1 | 3.6±1.9 | 23.5±7.1 |
| R. raphanistrum | 28.3±7.6 | 19.5±4.7 | 2.3±0.5 | 15.4±2.2 |
| Seeds | ||||
| R. pugioniformis | 12.8±4.4 | 42.2±9.9 | 3.2±0.3 | 6.9±0.5 |
| R. raphanistrum | 4.2±2.0 | 34.6±3.9 | 3.6±0.5 | 9.2±5.0 |
| Whole plant | ||||
| R. pugioniformis | 45.9±8.4 | 30.5±5.2 | 4.2±1.0 | 15.8±3.0 |
| R. raphanistrum | 39.2±5.2 | 23.0±5.0 | 3.0±0.4 | 13.7±1.3 |
Table 5: Dry weight and nutrient contents (N, P, and K) in plant parts (leaves, stems, and seeds) of R. pugioniformis and R. raph
Nitrogen content was measured in the various plant parts of both species. R. pugioniformis exhibited higher nitrogen content compared to R. raphanistrum, with values of 34.5, 21.8, 42.2, and 30.5 mg/g for leaves, stem, seeds, and whole plants of R. pugioniformis, respectively. In contrast, R. raphanistrum had nitrogen content values of 32.0, 19.5, 34.6, and 23.0 mg/g for leaves, stem, seeds, and whole plants, respectively.
Phosphorus content in the leaves, stem, and whole plant of R. pugioniformis was higher than that of R. raphanistrum. However, the trend was reversed for phosphorus content in the seeds. Potassium content ranged from 6.9 to 23.5 mg/g in different parts of R. pugioniformis, with the lowest value observed in the seeds and the highest in the stem. In R. raphanistrum, potassium content ranged from 9.2 (seeds) to 15.4 (stem). These results indicate variations in the nutrient contents of nitrogen, phosphorus, and potassium among different parts of both R. pugioniformis and R. raphanistrum plants, as well as differences between the two species.
Accumulated Nutrients of R. pugioniformis and R. raphanistrum
Table 6 presents the measurements of accumulated nutrients (N, P, and K) in plant parts of R. pugioniformis and R. raphanistrum species. The accumulated nitrogen content in the leaves, seeds, and whole plant of R. pugioniformis was higher compared to R. raphanistrum, whereas the accumulated nitrogen content in the stem showed the opposite trend. Specifically, the nitrogen content was found to be 381.1, 458.2, 560.1, and 1399.5 mg/plant for leaves, stem, seeds, and whole plants of R. pugioniformis, respectively, while for R. raphanistrum, the values were 253.2, 528.8, 141.1, and 882.0 mg/plant.
Similarly, the accumulated phosphorus content in the leaves, stem, seeds, and whole plant of R. pugioniformis (71.9, 76.7, 40.8, and 189.5 mg/plant, respectively) was higher compared to R. raphanistrum (42.3, 65.9, 14.6, and 117.6 mg/plant, respectively). The same trend was observed for accumulated potassium content in the leaves, stem, seeds, and whole plant of R. pugioniformis (131.8, 507.6, 88.5, and 727.9 mg/plant, respectively), which was higher than that of R. raphanistrum (85.2, 424.5, 33.6, and 533.0 mg/plant, respectively).
These results indicate variations in the accumulated contents of nitrogen, phosphorus, and potassium among the different plant parts of both R. pugioniformis and R. raphanistrum. Furthermore, significant differences in accumulated nutrient contents were observed between the two species.
| N accumulated | P accumulated | K accumulated | |
|---|---|---|---|
| Leaves | |||
| R. pugioniformis | 381.1±36.1 | 71.9±10.9 | 131.8±28.2 |
| R. raphanistrum | 253.2±46.0 | 42.3±4.7 | 85.2±12.5 |
| Stem | |||
| R. pugioniformis | 458.2±57.2 | 76.7±35.3 | 507.6±167.3 |
| R. raphanistrum | 528.8±48.8 | 65.9±20.8 | 424.5±57.1 |
| Seeds | |||
| R. pugioniformis | 560.1±270.0 | 40.8±13.9 | 88.5±34.2 |
| R. raphanistrum | 141.1±56.7 | 14.6±5.4 | 33.6±10.4 |
| Whole plant | |||
| R. pugioniformis | 1399.5±323.2 | 189.5±24.2 | 727.9±197.2 |
| R. raphanistrum | 882.0±77.6 | 117.6±17.7 | 533.2±50.6 |
Table 6: Accumulated Nutrients (N, P, and K (Mg/Plant)) in Plant Parts (Leaves, Stems, and Seeds) of R. pugioniformis and R. raph
Nutrient Contents of R. pugioniformis from Faqua and R. raphanistrum from Yabad
Table 7 presents the comparison of nutrient content (N, P, and K) and dry weight in different plant parts (shoot, seeds, and whole plant) of R. pugioniformis collected from Faqua and R. raphanistrum collected from Yabad in their maturity stage (May 2021).
For dry weight, the shoot of R. pugioniformis ranged from 63.4 to 91.0 g, while the seeds ranged from 4.0 to 6.7 g, and the whole plant ranged from 67.4 to 95.1 g. In contrast, the shoot of R. raphanistrum had a dry weight of 40.3 g, the seeds had a dry weight of 5.1 g, and the whole plant had a dry weight of 45.5 g. Generally, R. pugioniformis exhibited higher dry weight values compared to R. raphanistrum.
In terms of nitrogen content, the shoot of R. pugioniformis ranged from 12.3 to 13.4 mg/g, the seeds ranged from 63.4 to 71.8 mg/g, and the whole plant ranged from 14.4 to 17.5 mg/g. In R. raphanistrum, the shoot had a nitrogen content of 82.1 mg/g, the seeds had a nitrogen content of 62.0 mg/g, and the whole plant had a nitrogen content of 80.0 mg/g. The nitrogen content was highest in the seeds of both species.
Regarding phosphorus content, the shoot of R. pugioniformis ranged from 2.6 to 3.7 mg/g, the seeds ranged from 11.6 to 15.4 mg/g, and the whole plant ranged from 3.0 to 4.5 mg/g. In R. raphanistrum, the shoot had a phosphorus content of 12.1 mg/g, the seeds had a phosphorus content of
8.3 mg/g, and the whole plant had a phosphorus content of 11.7 mg/g. Similar to nitrogen, the phosphorus content was highest in the seeds of both species.
For potassium content, the shoot of R. pugioniformis ranged from 15.1 to 21.2 mg/g, the seeds ranged from 19.5 to 27.6 mg/g, and the whole plant ranged from 15.3 to 21.7 mg/g. In R. raphanistrum, the shoot had a potassium content of 27.9 mg/g, the seeds had a potassium content of 24.3 mg/g, and the whole plant had a potassium content of 27.4 mg/g. Similar to nitrogen and phosphorus, the potassium content was highest in the seeds of both species.
Overall, the results indicate differences in nutrient content and dry weight among the different plant parts of R. pugioniformis and R. raphanistrum, with generally higher values observed in R. pugioniformis compared to R. raphanistrum.
| Parameter | R. pugioniformis | R. raphanistrum | ||
|---|---|---|---|---|
| Population 1 | Population 2 | Population 3 | Population 1 | |
| Shoot | ||||
| Dry weight (g) | 91±4.1 | 72.5±16.3 | 63.4±17.1 | 40.3±3.5 |
| N content (mg/g) | 13.3±0.9 | 12.3±4.6 | 13.4±6.2 | 82.1±11.3 |
| P content (mg/g) | 2.6±0.2 | 3.7±1.3 | 3.1±1.1 | 12.1±1.5 |
| K content (mg/g) | 15.1±4.1 | 19.0±2.8 | 21.2±4.7 | 27.9±5.4 |
| Seeds | ||||
| Dry weight (g) | 4.1±2.1 | 6.7±2.4 | 4.0±0.4 | 5.1±1.1 |
| N content (mg/g) | 65.0±0.8 | 63.4±7.2 | 71.8±14.0 | 62±21 |
| P content (mg/g) | 11.6±0.7 | 13.6±1.2 | 15.4±3.4 | 8.3±1.0 |
| K content (mg/g) | 19.5±0.7 | 22.0±2.3 | 27.6±8.5 | 24.3±1.9 |
| Whole plant | ||||
| Dry weight (g) | 95.1±5.9 | 79.2±18.5 | 67.4±17.4 | 45.5±4.1 |
| N content (mg/g) | 14.4±5.5 | 17.5±1.1 | 17.2±4.7 | 80.0±11.9 |
| P content (mg/g) | 3.0±0.3 | 4.5±1.3 | 3.9±1.4 | 11.7±1.2 |
| K content (mg/g) | 15.3±3.9 | 19.2±2.5 | 21.7±4.6 | 27.4±5.0 |
Table 7: Nutrient Contents (N, P, And K) in Plant Parts (Shoot, Seeds, and Whole Plant) of R. Pugioniformis Collected from Faqua
Accumulated Nutrient Contents of R. pugioniformis from Faqua and R. raphanistrum from Yabad
Table 8 presents the comparison of accumulated nutrient content (N, P, and K) in different plant parts (shoot, seeds, and whole plant) of R. pugioniformis collected from Faqua and R. raphanistrum collected from Yabad in their maturity stage (May 2021).
For total nitrogen content, the shoot of R. pugioniformis ranged from 903.9 to 1126.0 mg, the seeds ranged from 266.3 to 425.4 mg, and the whole plant ranged from 1379.6 to 1392.3 mg. In R. raphanistrum, the shoot had a total nitrogen content of 3323.6 mg, the seeds had a total nitrogen content of 325.3 mg, and the whole plant had a total nitrogen content of 3648.8 mg. The highest total nitrogen content was observed in the whole plant of both species.
Regarding total phosphorus content, the shoot of R. pugioniformis ranged from 189.0 to 263.9 mg, the seeds ranged from 48.2 to 90.1 mg, and the whole plant ranged from 250.0 to 354.0 mg. In R. raphanistrum, the shoot had a total phosphorus content of 486.5 mg, the seeds had a total phosphorus content of 43.2 mg, and the whole plant had a total phosphorus content of 529.7 mg. The highest total phosphorus content was observed in the whole plant of both species.
For total potassium content, the shoot of R. pugioniformis ranged from 1346.5 to 1387.0 mg, the seeds ranged from 78.5 to 151.0 mg, and the whole plant ranged from 1455.7 to 1520.7 mg. In R. raphanistrum, the shoot had a total potassium content of 1115.9 mg, the seeds had a total potassium content of 125.2 mg, and the whole plant had a total potassium content of 1241.1 mg. The highest total potassium content was observed in the whole plant of R. pugioniformis.
Comparing the accumulated nutrient contents, it is generally higher in the whole plant and shoot of R. raphanistrum compared to R. pugioniformis, while R. pugioniformis showed higher total potassium content in the whole plant compared to R. raphanistrum.
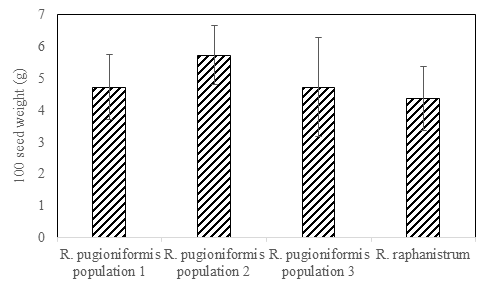
Furthermore, Figure 2 shows the weight of 100 seeds (in grams) of R. pugioniformis and R. raphanistrum collected from Yabad in their maturity stage (May 2021). The weights were 4.7, 5.7, and 4.7 g for the three populations of R. pugioniformis, and 4.4 g for the seeds of R. raphanistrum.
| Parameter | R. pugioniformis | R. raphanistrum | ||
|---|---|---|---|---|
| Population 1 | Population 2 | Population 3 | Population 1 | |
| Shoot | ||||
| Total N (mg) | 1126.0±465.2 | 954.2±166.4 | 903.9±599.7 | 3323.6±620.4 |
| Total P (mg) | 234.6±26.5 | 263.9±112.1 | 189.0±49.8 | 486.5±58.5 |
| Total K (mg) | 1387.0±441.6 | 1369.6±354.0 | 1346.5±531.3 | 1115.9±177.4 |
| Seeds | ||||
| Total N (mg) | 266.3±138.0 | 425.4±156.6 | 285.2±35.1 | 325.3±161.2 |
| Total P (mg) | 48.2±27.1 | 90.1±26.8 | 60.9±9.0 | 43.2±13.2 |
| Total K (mg) | 78.5±37.6 | 151.0±65.4 | 109.2±26.1 | 125.2±30.2 |
| Whole plant | ||||
| Total N (mg) | 1392.3±603.1 | 1379.6±277.5 | 1379.6±568.9 | 3648.8±710.1 |
| Total P (mg) | 282.8±40.0 | 354.0±122.1 | 250.0±48.3 | 529.7±70.6 |
| Total K (mg) | 1465.5±469.2 | 1520.7±408.0 | 1455.7±510.7 | 1241.1±206.1 |
Table 8: Accumulated Nutrients (N, P, And K) in Plant Parts (Shoot, Seeds, and Whole Plant) of R. Pugioniformis Collected from Fa
Discussion
Both wild radish species, R. raphanistrum and R. pugioniformis, share similarities in their reproductive phase duration. However, R. raphanistrum demonstrated a significantly higher fruit production per plant compared to R. pugioniformis. Although the fruits of R. pugioniformis were longer than those of R. raphanistrum, the latter contained a greater number of seeds. It is worth noting that while the fruits of R. raphanistrum disperse as single-seeded lightweight units, the fruits of R. pugioniformis remain intact [6, 7]. This key difference in seed dispersal strategies likely contributes to the extensive global distribution of R. raphanistrum. In the case of R. pugioniformis, there is a tradeoff between adaptive traits and fruit size, which provides an ecological advantage by enabling it to thrive in deep-soiled niches near the parent plant. These niches are often characterized by a high abundance of stones and gravel [7].
It is worth noting that the pink petal color in Raphanus species is governed by anthocyanin, and previous studies have shown a common regulatory link between anthocyanin synthesis and trichome formation [22]. This suggests a potential connection between differences in petal color and trichome density in the two species [23]. Moreover, it has been suggested that the presence of pink and yellow flower morphs in wild Raphanus sativus in North America serves as a plant strategy to deter herbivores. Given that trichomes also play a significant role in plant-insect interactions, it is possible that biotic factors, such as herbivory and pollinator preferences, contribute to the observed inter- species phenotypic differentiation in the two wild radishes. Therefore, the measured differences in fitness traits between the species may be influenced by the effects of floral traits on pollinator preferences in the field, as well as the potential tradeoff between trichome formation and reproductive success [24, 25].
Considerable inter-population phenotypic variation was observed during field observations. R. pugioniformis populations exhibited significant differences in nearly all the measured traits. It is worth noting that there is a strong correlation between elevation and temperatures in the region, and elevation was utilized as an approximation for temperature variations during the growth season. However, the impact of soil type could not be assessed due to its classification as a non-continuous numerical variable.
The impact of environmental parameters was generally more pronounced in R. pugioniformis, which supports the hypothesis that differences in seed dispersal distances contribute to inter-population phenotypic variation. In contrast, the distribution of R. raphanistrum in the southeast Mediterranean is more continuous and encompasses habitats that are relatively uniform, with variations primarily attributed to annual precipitation levels [7]. Nevertheless, it should be noted that the distribution range of these species does not encompass a significant aridity gradient, which is known to have an impact on phenological traits in other widely distributed plant species [20, 21]. On the contrary, the growing date (measured as the number of days from germination) in R. pugioniformis showed a positive correlation with annual rainfall and elevation. Furthermore, there were no differences in the total lifespan duration among populations of R. pugioniformis. These results were unexpected, considering the tradeoff between the duration of the reproductive stage and fitness traits, where a shorter flowering period usually leads to lower plant fitness. However, since populations of R. pugioniformis differed in various floral traits such as size and color, it is possible that the measured reproductive output is linked to pollinator preferences at the experimental site.
Conclusion
In conclusion, this study provides evidence for the adaptive phenotypic variation observed in the two closely related Raphanus species. Furthermore, the differences in phenotypic characteristics between the two Raphanus species are adaptive traits suited to their specific habitats. Variations in morphological characteristics, such as plant height, fruit length, tip/pod ratio, number of seeds to fruit ratio, and number of branches (primary and secondary), were observed between the two species collected from different locations in the West Bank at both the maturity and flowering stages.
Moreover, differences in the contents of nitrogen, phosphorous, and potassium were observed among different parts of both R. pugioniformis and R. raphanistrum plants. Additionally, accumulated contents of nitrogen, phosphorous, and potassium also varied between the two species. Specifically, R. raphanistrum exhibited higher potassium, nitrogen, and phosphorous content in the whole plant, seeds, and shoot compared to R. pugioniformis. These findings highlight the distinct nutrient dynamics and adaptations of each species in their respective habitats.
Acknowledgment
The authors are grateful to our donors Saarbrucken University. The authors are also thankful to Nasser Samara and Saleh Ali from Association for Integrated Rural Development (AIRD) who facilitate and supervise field work of this study.
Funding Sources
The study was fully funded by Saarbrucken University, grant number.
Conflict of Interest
The authors declares no conflict of interest.
References
-
Campbell LG, Snow AA, Ridley CE (2006) Weed evolution after crop gene introgression: Greater survival and fecundity of hybrids in a new environment. Ecology Letters 9: 1198-1209.
-
Campbell LG, Snow AA (2009) Can feral weeds evolve from cultivated radish (_Raphanus_ sativus, _Brassicaceae_)?. American Journal of Botany 96(2): 498-506.
-
Conner JK, Karoly K, Stewart C, Koelling VA, Sahli HF, et al. (2011) Rapid independent trait evolution despite a strong pleiotropic genetic correlation. The American Naturalist 178(4): 429-441.
-
Al-Shehbaz I (2012) A generic and tribal synopsis of the _Brassicaceae_ (Cruciferae). Taxon 61(5): 931-954.
-
Klinger T, Elam DR, Ellstrand NC (1991) Radish as a model system for the study of engineered gene escape rates via crop-weed mating. Conservation Biology 5(4): 531-535.
-
Ziffer-Berger J, Hanin N, Fogel T, Mummenhoff K, Barazani O (2015) Molecular phylogeny indicates polyphyly in _Raphanus_ L. (_Brassicaceae_). Edinburgh Journal of Botany 72(1): 1-11.
-
Ziffer-Berger J, Waitz Y, Behar E, Joseph OB, Bezalel L, et al. (2020) Seed dispersal of wild radishes and its association with within-population spatial distribution. BMC Ecology 20(30): 1-10.
-
Culpepper S (2012) Managing wild radish in wheat. Technical Report, University of Georgia.
-
Holm LG (1997) World Weeds. Natural Histories and Distribution. John Wiley & Sons Inc.
-
Bett KE, Lydiate DJ (2003) Genetic analysis and genome mapping in _Raphanus_. Genome 46(3): 423-430.
-
Conner JK (2002) Genetic mechanisms of floral trait correlations in a natural population. Nature 420(6914): 407-410.
-
Conner JK, Franks R, Stewart C (2003) Expression of additive genetic variances and covariances for wild radish floral traits: Comparison between field and greenhouse environments. Evolution 57(3): 487-495.
-
Lehtilä K, Strauss SY (1999) Effects of foliar herbivory on male and female reproductive traits of wild Radish, _Raphanus_ raphanistrum. Ecology 80(1): 116-124.
-
Malik MS (2009) Biology and ecology of wild radish (_Raphanus_ raphanistrum) Clemson University Tiger Prints, All Dissertations pp: 1-161.
-
Malik (2009) Clemson University.
-
Morgan MT, Conner JK (2001) Using genetic markers to directly estimate male selection gradients. Evolution 55(2): 272-281.
-
Moghe GD, Hufnagel DE, Tang H, Xiao Y, Dworkin I, et al. (2014) Consequences of whole-genome triplication as revealed by comparative genomic analyses of the wild radish _Raphanus raphanistrum_ and three other _Brassicaceae_ species. The Plant Cell 26(5): 1925-1937.
-
Pakniyat H, Powell W, Baird E, Handley LL, Robinson D, et al. (1997) AFLP variation in wild barley (Hordeum spontaneum C. Koch) with reference to salt tolerance and associated ecogeography. Genome 40(3): 332-341.
-
Volis S (2007) Correlated patterns of variation in phenology and seed production in populations of two annual grasses along an aridity gradient. Evolutionary Ecology 21: 381-393.
-
Waitz F, Schnaiter M, Leisner T, Järvinen E (2021) Phips-halo: the airborne particle habit imaging and polar scattering probe – part 3: Single particle phase discrimination and particle size distribution based on angular scattering function. Atmospheric Measurement Techniques 14: 3049-3070.
-
Westberg E, Ohali S, Shevelevich A, Fine P, Barazani O (2013) Environmental effects on molecular and phenotypic variation in populations of Eruca sativa across a steep climatic gradient. Ecol Evol 3(8): 2471- 2484.
-
Gao C, Guo Y, Wang J, Li D, Liu K, et al. (2018) Brassica napus GLABRA3-1 promotes anthocyanin biosynthesis and trichome formation in true leaves when expressed in Arabidopsis thaliana. Plant Biology 20(1): 3-9.
-
Irwin RE, Strauss SY, Storz S, Emerson A, Guibert G (2003) The role of herbivores in the maintenance of a flower color polymorphism in wild radish. Ecology 84(7): 1733-1743.
-
Aronson J, Kigel J, Shmida A, Klein J (1992) Adaptive phenology of desert and Mediterranean populations of annual plants grown with and without water-stress. Oecologia 89: 17-26.
-
Züst T, Joseph B, Shimizu KK, Kliebenstein DJ, Turnbull LA (2011) Using knockout mutants to reveal the growth costs of defensive traits. Proceedings of Biological Sciences 278(1718): 2598-2603.
- Evaluation of Comparative Morphological and Phytochemical Studies on the Seeds Extracts of Cocos nucifera (L.) and Elaeis guineensis Jacq. (Arecaceae)
- Aborted Spores in Argentine Ferns
- Ficus middletonii Chantaras. – A New Distributional Record For Central India
- A Study on the Phytochemical, Antioxidant and Antimicrobial Activities of Ficus glumosa Delile (Moraceae)
- Paclobutrazol (PBZ) and its Metabolic Function in Agriculture: A Review
- Decalepis arayalpathra: Ethnobotany, Scientific Interventions and Prospects