Biopotency of Gynandropsis Gynandra L., against Aflatoxin B1Induced Rats with Reference to Tumour Marker Enzymes
Gynandropsis gynandra L., has been reported to be useful in treatment of various ailments in the Indian system of medicine. We proposed to investigate the biopotency of G.gynandra extract against aflatoxin B1-induced hepatocellular carcinoma in rats. A significant increase (p<0.001) in the activities of cancer marker enzymes γ-GT, LDH, ALP and 5|-NT in serum, liver and kidney with a significant elevation in the serum transaminases with a concomitant decrease in the liver and kidney transaminases activities in AFB1-induced HCC bearing rats were observed when compared with the normal control rats. Treatment with the drug GGE showed a significant reversal of the activities of all the enzymes both in serum and liver. These results indicate that G. gynandra extract could potentially modulate the cancer marker enzymes and thus could elicit suppressive activity on tumour growth in AFB1-induced male albino rats.
Introduction
Aflatoxin B1 (AFB1), a secondary metabolite produced by the fungal moulds Aspergillus flavus and A.parasiticus are widely encountered as a contaminant of cereal crops and nuts in humid areas of the world. There is a strong correlation between dietary intake levels and cancer incidence in human populations [1]. Hepatocellular carcinoma (HCC) is one of the leading cause of mortality in regions of world where environmental levels of aflatoxin B1 (AFB1) are high and it has provide strong circumstantial evidence that this mycotoxin is a principal etiological factor in human liver cancer in Asia and Africa [2, 3]. AFB1, the most common mycotoxin is a potent Biopotency of Gynandropsis Gynandra L., against Aflatoxin B1- Induced Rats with Reference to Tumour Marker Enzymes mutagen and naturally occurring hepatocarcinogen [4]. Earlier studies on AFB1-induced hepatocarcinogenesis in experimental animals revealed that even single dose of AFB1 could produce hepatocellular carcinoma in rats [5]. Any damage in liver results in increased permeability of hepatocellular enzymes in to the blood stream reflects the loss of cellular integrity or due to cell necrosis [6]. Analysis of these enzymes reflects the mechanisms of cellular damage and subsequent release of proteins, their extra-cellular turnover and also the mechanism of neoplastic processes [7]. Damaged cells lose their internal mileu including the enzymes, which are distributed all over the extra-cellular space. The rise in their activity in serum has been shown to be in good correlation with the J Cancer Oncol
number of transformed cells in cancer condition [8]. Since the marker enzymes have a high specificity for hepatocarcinogenesis, these can be used as a diagnostic tool to confirm HCC and for its therapy.
Nutritional intervention for the treatment of cancer in their natural form is a promising strategy of recent research. Plant flavonoids and polyphenols are reported to reduce the formation of DNA adduct and thereby counteracting the carcinogenic action of AFB1 [9]. Gynandropsis gynandra L., (Capparidaceae) (English: Caravella) is an important medicinal herb consumed as a leafy vegetable in diet. It is found to be distributed in the tropical and sub-tropical parts of the India and all over the world. Earlier reports on phytochemical analysis of the plant reveal that, the plant contains various derivatives of kaempferol and quercetin, β-carotene, tannins, cleomin, hexacosanol and β-sitosterol [10]. The aqueous extract of the whole plant has reported to show antibacterial and antioxidant activities [11, 12]. Alcoholic extract of the whole plant exhibited anticancer activity against human epidermal carcinoma and hepatoma 129 in mouse [13]. Methanolic extract of the leaves has reported to processes a strong anti-inflammatory activity against the adjuvant induced arthritic rats [14]. Previous studies of our laboratory showed 50% hydroalcoholic extract of the plant possesess anti-proliferative activity, preventive against AFB1-induced lipid peroxidation, potentially regulated the altered glucose metabolizing enzymes during AFB1-induced carcinogenesis in rats and chemopreventive effects agains AFB1 in male albino rats [15, 16, 17, 18]. Based on these reports, we proposed to investigate the anticancer effect of G. gynandra (GGE) against AFB1-induced HCC in male albino rats with reference to tumour marker enzymes.
Materials and Methods
Chemicals
Aflatoxin B1 (AFB1) was purchased from Sigma Chemicals Co., St. Louis Mo., USA. All other chemicals and reagents were of the highest purity of analytical grade and obtained from the local firms.
Animals
Albino male rats of Wistar strain weighing 80-120g were used for the study. The rats were fed with commercial pelleted rat chow and water ad libitum and maintained under standard laboratory condition with 12h light and dark cycle. All the animal experiments were carried out according to the guidelines of the Institutional Animal Ethics Committee.
Plant Material
The whole plant of G. gynandra was collected during September to November 2005 from Athikkottai, Thanjavur District, India. The plant was identified and authenticated by Dr. M.Jegadesan, Professor of Medicinal Botany, Department of Siddha Medicine, Tamil University, Thanjavur, India, and the voucher specimen (no.263) has been deposited in the department herbarium. Aerial parts of the plant were rinsed in distilled water to remove the impurities, cut in to pieces, dried under shade for a week, coarsely powdered and extracted in 50% alcohol (v/v) using a soxhlet apparatus. The extract was filtered and evaporated to separate the residue and the yield was found to be 27.8% (w/w). The residue thus obtained was stored in an air tight container in a desiccator until further use.
Experimental Design
The rats were divided into four groups of six rats each. Group I rats received a single intraperitoneal dose (0.5ml) of DMSO. Hepatocellular carcinoma (HCC) was induced in group II and III rats by a single i.p., dose of AFB1 in 0.5ml of DMSO (2mg kg-1 body weight) [5]. After eight weeks of AFB1 injection, rats in group III were treated with the GGE (250mg kg-1 body weight) orally twice a day for three weeks by intragastric intubation [16, 17, 18]. Group IV rats served as drug control and received the same dosage of drug as per group III by the same route.
Collection of Samples and Determination of Enzyme Activities
At the end of the experiments, rats were fasted for overnight and sacrificed by cervical decapitation under mild ether anesthesia. The blood sample was collected to separate the serum. The liver and kidney were removed after perfusion with physiological saline, blotted dry, weighed and homogenized in Tris-HCl buffer 0.1 M (pH 7.4) to get a 10% homogenate. The serum and the 10% homogenate of the liver and kidney tissues were used for the determination of the activities of γ-glutamy transpeptidase (γ –GT) by the method of Rosalki & Rao [19], transaminases (AST & ALT) [20], lactate dehydrogenase (LDH) [21], alkaline phosphatase (ALP) by King [22] and 5|-nucleotidase (5|NT) by Luly, et al. [23]. Total protein was estimated by employing the method of Lowry, et al. [24] using BSA as standard.
Histology
Histology Formalin fixed liver tissues were dehydrated using gradient concentrations of ethanol; then washed in xylene and embedded in paraffin wax. Tissue blocks were sectioned at 5-6 μM thickness, deparaffinized and stained with hematoxylin and eosin and analyzed under microscope (Olympus Instruments, Tokyo, Japan).
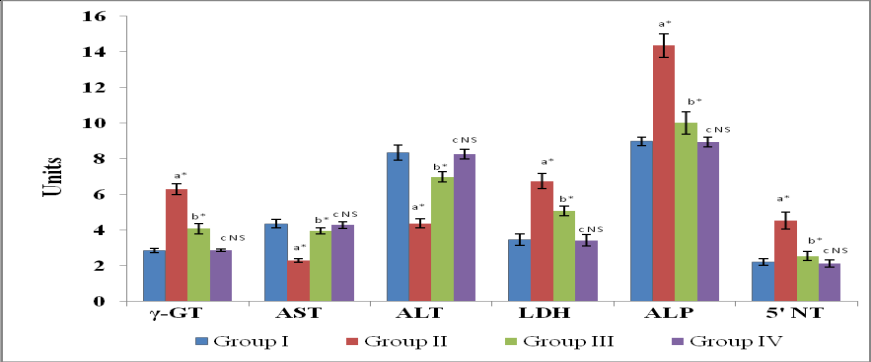
Table 1 shows the activities of tumour marker enzymes in serum of control and experimental rats. Group II AFB1-induced HCC bearing rats showed a significant increase (P<0.001) in the activities of tumour marker enzymes in serum namely, γ-GT, transaminases, LDH, ALP, 5|-NT, when compared with the normal control rats. Administration of the drug GGE significantly decreased (P<0.001) the activities of all the marker enzymes when compared with the HCC-bearing group II rats. Group IV drug control rats did not show any significant variation in the activities of marker enzymes in serum when compared with the group I normal control rats.
Statistical Analysis
The values were mean + SD for six rats in each group and statistically significant differences between mean values were determined by one-way analysis of variance (ANOVA), followed by the Tukey’s test. For multiple comparison, values of P<0.05 were considered as significant. Statistical package for social studies (SPSS) 7.5 version was used for this analysis.
Results
| Parameters | Group I | Group II | Group III | Group IV |
| $\gamma$-GT‡ | 1.42±0.049 | 3.86±0.23* | 2.09±0.12* | 1.50±0.06NS |
| AST§ | 78.63±3.25 | 148.63±11.84* | 97.38±2.94* | 81.36±2.79NS |
| ALT§ | 97.98±4.68 | 168.37±13.61* | 119.37±4.56* | 100.38±3.53NS |
| LDH§ | 118.35±3.06 | 258.65±3.86* | 145.18±3.66* | 121.36±3.42NS |
| ALP§ | 188.32±9.53 | 246.28±21.21* | 198.35±14.004* | 189.00±10.60NS |
| 5I-NT¥ | 2.81±0.179 | 6.92±0.547* | 3.62±0.184* | 2.79±0.162NS |
Table 1: Marker enzymes in serum of control and experimental rats.
Table 1: Marker enzymes in serum of control and experimental rats. Notes: Values are expressed as mean ±SD for six animals. Comparisons are made between (a) group I and group II, (b) group II and group III, and (c) group IV and group I. Statistical significance of difference is expressed as P *<0.001; NS, difference is not significant. Units: ‡μM of p-nitroaniline liberated min-1 mg protein-1; §μM of pyruvate liberated min-1 mg protein-1; $μM of Phenol liberated min-1 mg protein-1; ¥nM of Inorganic phosphorus liberated min-1 mg protein-1.
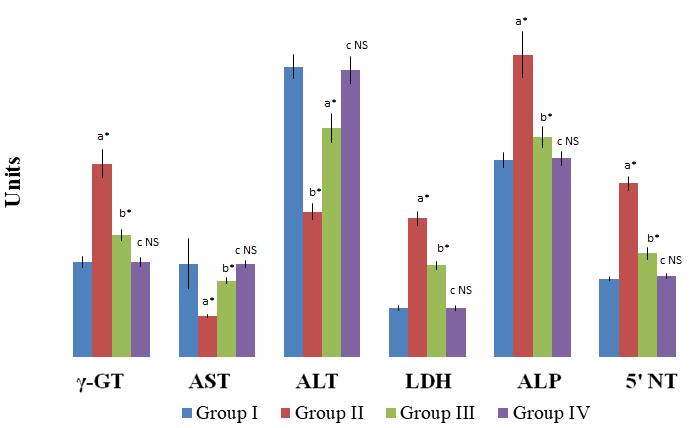
A significant enhancement (P<0.001) in the activities of the tumour marker enzymes namely γ-GT, LDH, ALP and 5|-NT, with a significant decrease (P<0.001) in the activity of transaminases (Figures 1 & 2) were observed in liver and kidney of group II HCC bearing rats. A significant (P<0.001) reversal in the activities of γ-GT, transaminases, LDH, ALP and 5|-NT were observed in group III rats after treatment with the drug GGE. No significant variations were observed in the activities of these enzymes in group IV drug control rats when compared with the group I normal control rats.
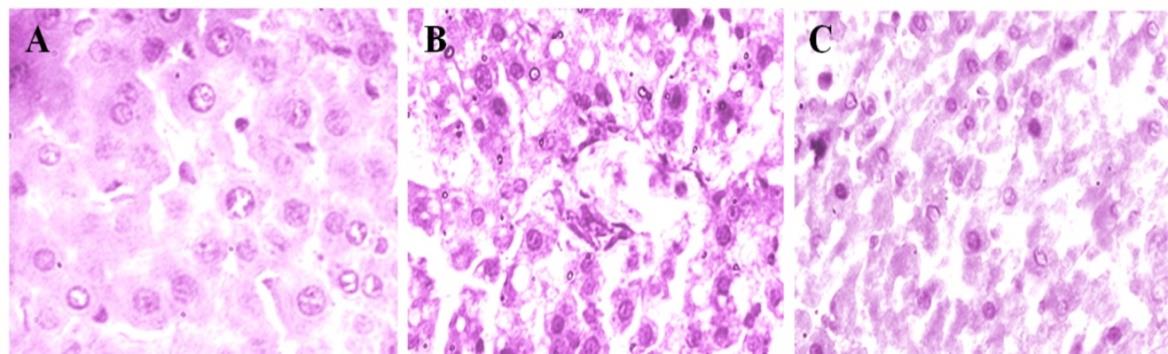
Liver sections from the control group showed normal lobular pattern with central vein and peripheral portal veins (Figure 3A). In contrast, the animals treated with AFB1 showed loss of architecture with early dysplastic cells exhibiting increased Cytoplasm:Nucleus ratio with the marked congestion and carcinomatous changes (Figure 3B). However, treatment with GGE to AFB1 induced rats showed (Figure 3C) almost normal architecture.
Discussion
Studies have demonstrated that disruption of many biochemical properties including marker enzymes of the host cells in aflatoxin B1-mediated cancer condition [25, 26]. Increase in the activities of γ-GT, transaminases, LDH, ALP and 5|-NT were recognized as potential tumour markers in assessing the progression of malignant cells [27]. The conspicuous increase in the activity of γ-GT observed in the AFB1-induced HCC bearing rats in our study is in consistent with the earlier report of Taniguchi et al., with an elevated level of γ-GT during chemical induced hepatocarcinogenesis [28]. The induction of high level of γ-GT is a frequent early event in experimental hepatocarcinogenesis in animals and also in human carcinoma including primary hepatic carcinoma [29]. Oxidative metabolism of extra cellular GSH initiated by high levels of γ-GT in preneoplastic foci may produce damage to cells and that could lead to an increased frequency of genetic alterations and/or tumour promotion effects [30]. These effects would help to move the preneoplastic cells further along the track from preneoplasia to neoplasia [31]. Decreased level of γ-GT observed in the drug treated rats in our study becomes an evident to suggest that, G. gynandra treatment nullify the deleterious effects in AFB1-induced HCC bearing rats by increasing the biliary elimination of toxic metabolites generated during cancer conditions, and thereby decreased the rate of genetic alterations and/or tumor promotion effects [32].
Transaminesemia becomes gradually more pronounced towards the terminus and which indicates the severity or advanced state of liver carcinoma [33]. Kamdem, et al. reported that AFB1-administration results in 65-290% increase in the activity of transaminases [8]. Results of our study are also in agreement with the above report. The elevated levels of serum AST and ALT with a concomitant decrease in the liver and kidney tissues of AFB1-induced HCC bearing rats may be due to the AFB1- induced cellular disintegration and loss of functional integrity of the plasma membrane, cell necrosis indicating organ dysfunction and cellular injury thereby caused increased leakage of the enzymes into the circulation [6, 34]. The reduction in the activity of serum AST and ALT and with an increment in the liver and kidney tissues after drug treatment in the HCC bearing rats observed in our study may be due to the stabilization of plasma membrane and repair of the tissues and thereby decreasing the leakage of the cellular enzymes into circulation. The beneficial effects afforded by G. gynandra against the AFB1-induced destabilization of the plasma membrane during HCC may be due to the presence of flavonoids such as kaempferol and quercetin derivatives. Flavonoids present in medicinal plants were reported to decrease the activity of serum transaminases in toxicated rats by stabilizing the leakage of the enzymes [35]. Increased activity of LDH observed in group II rats in our study may be due to, carcinogens that are known to cause cellular disintegration, mitochondrial damage and anaerobiosis [36, 37, 38]. Increased rate of anaerobic glycolysis during cancer growth leads to accumulation of lactic acid and causes acidic environment [39]. The reduction in the activity of LDH observed after treatment with GGE to AFB1-induced HCC bearing rats could be due to the regulation of glycolysis in cancer cells by the flavonoids and polyphenols [16, 17], which in turn facilitate the regulation of LDH.
The rise in the activity of ALP in AFB1-induced HCC bearing rats in our study is well correlated with the report of Mirmomeni et al., with a considerable elevation in ALP during precancerous like lesions [40]. Koss and Greengard [41] have also postulated a striking rise in the serum ALP activity up to 3 to 20 fold in several tissues of rats carrying neoplasm. In addition to the rise in ALP, the 5|-NT was also found to be elevated in AFB1-induced HCC bearing rats in our study. The sequential distribution of 5|-NT should be of value in assessing the relative importance of altered metabolic pathways during hepatocarcinogenesis. Elevation in 5|-NT activity during hepatocarcinogenesis displays an interesting pattern on the plasma membrane, nucleus and in the cytoplasm and has been linked with the changes in the functional and constitutional aspect of plasma membrane [42]. The decreased activities of both ALP and 5|-NT in serum and liver of AFB1-induced HCC bearing rats after treatment with GGE may suggest the recovery from aflatoxin B1 mediated changes in the metabolic, functional and constitutional aspect of plasma membrane, nucleus and cytoplasm. The recovery of the cells against AFB1-induced changes upon treatment of G. gynandra may be due to the flavonoids and polyphenols. Prior reports suggest that plant flavonoids and polyphenol afford protection to cells during carcinogenesis by decreasing the activities of marker enzymes such as LDH, ALP and 5|-NT [43].
In conclusion, the phytoactive constituents such as flavonoids (kaempferol and quercetin derivatives) and polyphenolic tannins reported to present in G. gynandra potentially protects the AFB1-induced tumour promotion by regulating the metabolic, functional and constitutional changes in plasma membrane, nucleus and cytoplasm and recovered cellular integrity. Thus, this study provides substantial evidence for the possible role of G. gynandra against AFB1-induced carcinogenesis through regulating the marker enzymes and regression of HCC in male albino wistar rats.
Conflict of Interest
The authors declare that there are no conflicts of interest.
References
-
Hamid AS, Tesfamariam IG, Zhang Y, Zhang ZG (2013) Aflatoxin B1-induced hepatocellular carcinoma in developing countries: Geographical distribution, mechanism of action and prevention. Oncol Lett 5(4): 1087-1092.
-
Van Rensburg SJ, Cook-Mozaffari P, Van Schalkwyk DJ, Van Der Watt JJ, Vincent TJ, et al. (1985) Hepatocellular carcinoma and dietary aflatoxin in Mozambique and Transkei. Br J Cancer 51(5): 713- 726.
-
Peers F, Bosch X, Kaldor J, Linsell A, Pluijmen M (1987) Aflatoxin exposure, hepatitis B virus infection and liver cancer in Swaziland. Int J Cancer 39(5): 545- 553.
-
Nixon JE, Hendricks JD, Pawloswki NE, Loveland PM, Sinnhuber RO (1981) Carcinogenicity of aflatoxicol in Fischer 344 rats. J Natl Cancer Inst 66(6): 1159-1163.
-
Angsubhakorn S, Get–Ngern P, Miyamoto M, Bhamarapravati N (1990) A single dose-response effect of aflatoxin B1 on rapid liver cancer induction in two strains of rats. Int J Cancer 46(4): 664-668.
-
McIntyre N, Rosalki S (1992) Biochemical Investigation in the management of liver disease. In: Prieto J, Rodes J, Shafritz DA, (Eds.), Hepatobiliary diseases, Berlin, pp: 39-71.
-
Tazi A, Galteau MM, Siest G (1980) γ-Glutamyl transferase of rabbit liver: Kinetic study of phenobarbitol induction and _in vitro_ solubilization by bile salts. Toxicol Appl Pharmacol 55(1): 1-7.
-
Kamdem L, Siest G, Magdalou J (1982) Differential toxicity of aflatoxin B1 in male and female rats, relationship with hepatic drug metabolizing enzymes. Biochem Pharmacol 31(19): 3057-3062.
-
Bhattacharya RK, Firozi PF (1988) Effect of plant flavonoids on microsome catalyzed reactions of aflatoxin B1 leading to activation and DNA adduct formation. Cancer Lett 39(1): 85-91.
-
Sharaf M, EL-Ansari MA, Saleh NAM (1997) Flavonoids of four cloeme and three capparis species. Biochemical Systematics and Ecology 25(2): 161-166.
-
Perumal Samy R, Ignacimuthu S, Raja DP (1999) Preliminary screening of ethnomedicinal plants from India. J Ethnopharmacol 66(2): 235-240.
-
Cook JA, Vanderjagt DJ, Dasgupta A, Mounkaila G, Glew RS, et al. (1998) Use of the Trolox assay to estimate the antioxidant content of seventeen edible wild plants of Niger. Life Sciences 63(2): 106-110.
-
Uniyal MR, Tewari LC (1991) Anti-Cancer Drugs from U.P. Himalaya. Ancient Science of Life 11(1-2): 50-55.
-
Narendhirakanna RT, Kandhaswamy M, Subramanian S (2005) Anti-inflammatory ctivity of _Cleome_ _gynandra_ L. on hematological and cellular constituents against adjuvant-induced arthritic rats. J Med Food 8(1): 93-99.
-
Surendrakumar P, Sadique J (1987) The biochemical mode of action of _Gynandropsis gynandra_ in inflammation. Fitoterapia L 8: 379-385.
-
Sivanesan D, Hazeena Begum V (2007) Preventive role of _Gynandropsis gynandra_ L., against aflatoxin B1 induced lipid peroxidation and antioxidant defense mechanism in rat. Indian J Expt Biol 43: 299-305.
-
Sivanesan D, Hazeena Begum V (2007) Modulatory effect of _Gynandropsis gynandra_ L., on glucose metabolizing enzymes in aflatoxin B1- induced hepatocellular carcinoma in rats. Indian J Biochem Biophys 44(6): 477-480.
-
Sivanesan D, Malarvili T, Hazeena Begum V (2016) Chemopreventive action of _Gynandropsis gynandra_ L., agent against Aflatoxin-B1 in rats. Chemical Science Transactions 5(3): 803-808.
-
Rosalki SB, Rau D (1972) Serum γ-glutamyl transpeptidase activity in alcoholism. Clin Chim Acta 39: 41-47.
-
King J (1965) The transferases alanine and aspartate transaminases. In: Van D, (Ed.), Practical and Clinical Enzymology, London, pp: 121-128.
-
King J (1965) The dehydrogenases or oxido reductase-lactate dehydrogenase. In: Van D, (Ed.), Practical and Clinical Enzymology, London, pp: 83-93.
-
King J (1965) The hydrolysases-acid and alkaline phosphatase. In: Van D, (Ed.), Practical and Clinical Enzymology, London, pp: 191-208.
-
Luly P, Barnabei O, Tria E (1972) Hormonal control _in_ _vitro_ of plasma membrane bound Na+, K+-ATPase of rat liver. Biochim Biophys Acta 282(1): 447-452.
-
Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin-phenol reagent. J Biol Chem 193(1): 265-275.
-
Venkita Subramanian TA, Saraswathy S (1985) Biological effects of aflatoxin, Biol Membr 8: 184-191.
-
Hyder MA, Hasan M, Mohieldein AH (2013) Comparative Levels of ALT, AST, ALP and GGT in Liver associated Diseases. Euro J Exp Biol 3: 280-284.
-
Ray PK, Singh KP, Suddin R, Prasad AK (1991) Immunological responses to aflatoxins and other chemical carcinogens. J Toxicol Toxin Rev 10(1): 63- 85.
-
Stark AA, Zeiger E, Pagano DA (1993) Glutathione metabolism of gamma-glutamyltranspeptidase leads to lipid peroxidation: characterization of the system and relevance to hepatocarcinogenesis. Carcinogenesis 14(2): 183-189.
-
Taniguchi N, Tsukada Y, Hirai MH (1974) Effect of hepatocarcinogenic azodyes on glutathione and related enzymes in rat liver. Gann 65(5): 381-387.
-
Bannasch P (1986) Preneoplastic lesions as end points in carcinogenecity testing I Hepatic preneoplasia. Carcinogenesis 7(5): 698-695.
-
Fiala S, Mohindru A, Kettering WG, Fiala EA, Morris HP (1973) Glutathione and γ - GT in rat liver during chemical carcinogenesis. J Natl Cancer Inst 57(3): 591-598.
-
Kensler PTW, Egner A, Davidson NE, Roebuck BD, Pikul A, et al. (1986) Modulation of aflatoxin 7N- guanine formation and hepatic tumuorigeneis in rats fed ethoxyquin: role of induction of glutathione S- transferase Cancer Res 46(8): 3924-3931.
-
Stedt SWR, Walder OS (1974) The diagnostic significance of a high ASAT, ALAT ratio in patients with very high serum aminotransferase levels. Acta Med Scand 195(3): 227-229.
-
Drotman RB, Lawhorn GT (1978) Serum enzymes as indicators of chemical induced liver damage. Drug Chem Toxicol 1(2): 163-171.
-
Sanz MJ, Ferrandiz MI, Cejudo M, Terencio MC, Gil B, et al. (1994) Influence of a series of natural flavonoids on free radicals generating systems and oxidative stress Xenobiotica 24(7): 689-699.
-
Chuang SE, Chen AL, Lin JK, Kuo ML (2000) Inhibition by curcumin of diethylnitrosamine-induced hepatic hyperplasia, inflammation, cellular gene products and cell-cycle-related proteins in rats. Food Chem Toxicol 38(11): 991-995.
-
Wang CJ, Wang JM, Lin CY, Chu CY, Chou FP, et al. (2000) Protective effect of _Hibiscus anthocyanins_ against tert-butyl hydroperoxide-induced hepatic toxicity in rats. Food Chem Toxicol 38(5): 411-416.
-
Silva VM, Chen C, Hennig GE, Whiteley HE, Manauto JE (2001) Changes in susceptibility to acetaminophe- induced liver injury by the organic anion indocyanine green. Food Chem Toxicol 39(3): 271-278.
-
Narasimhan S, Schachter H, Rajalakshmi S (1988) Expression of N-acetylglucosaminyltransferase III in hepatic nodules during rat liver carcinogenesis promoted by orotic acid. J Biol Chem 263(3): 1273- 1281.
-
Mirmomeni MH, Suzangar M, Wise A, Messripour M, Emami H (1979) Biochemical studies during aflatoxin B1 induced liver damage in rats fed different levels of dietary protein. Int J Cancer 24(4): 471-476.
-
Koss B, Greengard O (1982) Effect of neoplasms on the content and activity of alkaline phosphatase and gamma-glutamyl transpeptidase in uninvolved host tissues. Cancer Res 42(6): 2146-2158.
-
Duve De C (1963) Ciba foundation symposium on lysosomes, in: Avs de Reuck, Cameron, MP, (Edn.), JA Churchill, London.
- Cancer Diagnosis from RNA Sequence of Blood Cells by Using AI
- Field Cancerization in Oral Cavity, Case Report and Review of Literature. Oncologic Program Salud Integral Hospital, Managua, Nicaragua
- Identification of B Lymphocytes in Cancer Patient’s Blood
- A Case Report of a Breast Cancer Patient Developing Pneumonitis as a Result of Abemaciclib Therapy
- Immune Checkpoint Therapeutics for Today’s Fight and Beyond
- The Amalgamated Sophomore-Gonadoblastoma