Nuances of Erlich Ascites Carcinoma Cells Growth and how the External Electrostatic Field Affect Ascites Tumor Growth
In the present work, the Erlich Ascites Carcinoma (EAC) cells in vitro growth kinetics, and its correlation with surveillance of EAC-infected mice is reported. EAC cells as well as EAC-inoculated mice have been exposed to the external Electrostatic Field (ESF). The kinetics of EAC cells growth in vitro was characterized with classical periods: lag-phase-0-3 days; exponential phase-3-12 days; stationary phase- 12-16 days; decline phase-16-24 days of cultivation. However, for the first time, during 24 days cultivation, drops of cell concentration had been detected: at 7th, 14th, and 21st days. The observed phenomenon had not been described earlier for in vitro conditions, but resembles the in vivo observation shown in literature. Analysis of correlation between in vitro growth of cells and in vivo surveillance of mice revealed their inverse character at 7-15 days. Hence, the correlation between EAC cells growth in vitro and in vivo was shown. This may be a substantial contribution to the knowledge of tumor development. Above it, EAC infected mice exposed to the ESF shown diminished surveillance at the days of exposure and enhanced surveillance after stopping the ESF action. It has been shown that ESF cause an EAC cell damage as well as host leucocyte cell is. But unlike to leucocytes, EAC cells rapidly accelerate growing after the end of the exposure. So, finally ESF favors tumor growing with simultaneous inhibition of host immune defense.
Introduction
The knowledge of details of malignant cell development has a critical importance for cancer treatment. Hence, the finding of some correlation between tumor development in vivo, and tumor cells growth in vitro would be highly applicable. This correlation would give the possibility of in vitro knowledge extrapolation to in vivo cases, which might be helpful for chemotherapy design [1]. Mice Ehrlich Ascites Carcinoma (EAC) has been used as a malignancy model for the study of tumor development.
During series of previous in vitro investigations of EAC
Image Article
cell culture we had observed an unexpected phenomenon of decreasing of cell concentration at definite days of cell cultivation without media refreshing [2]. The observed phenomenon had circular character at longtime cultivation. There is a little information in literature which can be collated with our observation. Particularly, Zamai AS, et al. [3] described the decrease of EAC tumor on 12thday of cell transplantation into mice. Author explained it as a consequence of enhanced apoptosis.
Some data regarding the biological effects of external Electrostatic Field (ESF) allow to hypothesize that such a physical factor may be able to promote the induction and progression of cancer. In particular, several authors investigated the influence of the external ESF on several components of the pro-/antioxidant system of the organism. As reported previously [4], the external ESF (E=500V/m)
leads to the increase of intracellular reactive oxygen species (ROS) concentration. Nowadays, there are a lot of data evidencing the ROS-induced development of different chronic diseases, including cancer [5]. By interaction with lipids, proteins, and nucleic acids ROS can cause damage within different tissues [6, 7, 8]. Changes in DNA structure are of particular importance in cancer etiology. Indeed, ROS-induced structural changes of DNA generally include modifications of guanine, which brings to the G→T trans versions [9], thus promoting malignancy induction and progression, if such modifications take place in critical genes, such as oncogens or tumor supression genes [10].
Mutagenic and genotoxic activity of ESF, which was reported in several papers, can lead to cancer development [11, 12, 13]. Genotoxic activity of ESF of 200kV/m tension has been shown using the DNA-comet test on leucocytes of periferal blood of rats which was reflected on the cellular content of the blood [14, 15, 16]. Some studies report the mutagenic activity of ESF of higher tension and in particular conformational changes in DNA are now known to be caused by ESF exposure [17].
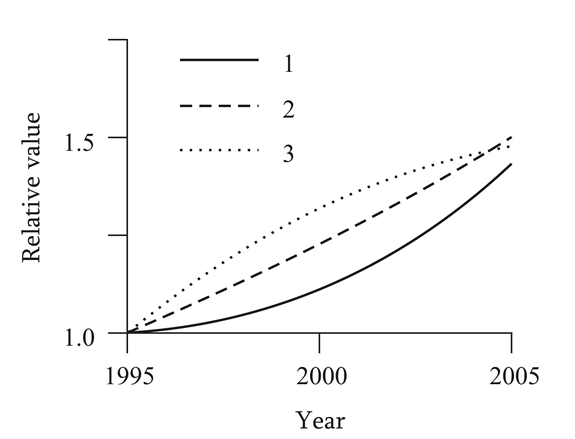
Some evidence of a possible influence of external electric fields on cancer pathogenesis is indirectly suggested also by several statistical and epidemiological findings. (Figure 1) presents some results concerning the production of electricity and synthetic fabrics during the period of 1995- 2005, along with the dynamics of tumor morbidity during the same period [18, 19, 20]. In everyday life and in manufacturing, both wide utilization of electricity and using of synthetic fabrics favour the increase of the level of ESF. It is noteworthy the similarity of the temporal dynamics shown. This may be seen as a potential evidence of a possible cause-effect link between the mentioned factors.
In the present paper, the EAC cell primary culture dynamics of was studied; the inverse correlation of cell mass accumulation in vitro and EAC-infected mice mortality in vivo was shown; and there was observed the phenomenon of cell concentration drop at definite times of cultivation. The effect of ESF on EAC tumor growth was demonstrated. It has been shown that ESF cause an EAC cell damage as well as host leucocyte cell is. But unlike to leucocytes, EAC cells rapidly accelerate growing after the end of the exposure. So, finally ESF favors tumor growing with simultaneous inhibition of host immune defense.
Materials and Methods
Animals and Cells
The objects of our investigation were the primary culture of EAC cells and EAC transplanted albino mice. EAC cells were continuously sub inoculated in mice by intraperitoneal injections every 10th-12thdays of prior inoculation. For in vitro experiments, the cells were cultured on 7th day of sub inoculation.
Cell Culturing Conditions
EAC cells were cultured in RPMI-1640 media supplemented with 0.3g/L L-glutamine, 50IU penicillin, and 10% fetal calf serum. Fresh EAC cells obtained from the peritoneal cavity of mice were suspended in the culture media in concentration 0.5×106 cells per ml and incubated during 24days in conditions of 37oC and 5% CO2 in highly humidified atmosphere. During incubation period the culture media was not refreshed.
ESF Exposure
ESF (E=200kV/m) exposure was applied both to the EAC infected mice (10days, 6hour per day) and to EAC cell culture (1hour). Leucocytes obtained from the blood of intact mice were used also exposed to ESF (1hour). ESF was generated in the special capacitor type device, which was designed on the basis of X-ray apparatus “RUM-17” (former USSR) [21]. Control samples were treated in the same manner with the exception of ESF exposure. Design of the experiment is detailed in the Table 1.
Proliferation and Viability Assay
Cell count was determined at different time intervals. Cells filled in a haemocytometer were scored using light microscope (Micros, Austira) and the results expressed by means of cell count ×106/ml. Cell growth kinetics was evaluated as the function of time. The growth rate constant and generation (doubling) time was quantified for 6-24hours incubation time interval [22]. The initial growth rate was calculated using the differential method. During the exponential phase we have: dN/dt= K’N, where N is the concentration of cells, t the time and K’ is the growth rate constant. Integration of previous equation between the limits of 0 and t and N1 and N2 gives following equation: Ln(N2/ N1)=K’(t2-t1), where N2 is cell number at t2 (24hours) and N1 is the cell number at t1 (6hours). The rate of EAC cell growth in culture was finally described by the time required for the cells duplication, or the generation time (Gen’t). Gen’t was calculated using following equation: Gen’t=Ln2/K’. Trypan blue exclusion method was applied for quantifying the ratio of viable cells [23].
Assay of Protein Carbonyl Content
Proteins carbonyl content was measured to determine the level of oxidative modification of proteins. Carbonylation was measured by the method of Levine RL, et al. [24] using the reaction with 2,4-dinitrophenylhydrasine (DNPH), in the cultured cell lysates using Spectrophotometer Hitachi 150-20UV-VIS (Japan). The protein carbonyl content was expressed in terms of nmol mg-1 of protein.
Alkaline Comet Assay
Alkaline Comet assay was conducted as described by Venturi M, et al. [25] with minor modifications. Cells were centrifuged at 100×g for 3min, the supernatant discarded and the remaining cells were suspended in 75μl of 0.5% low melting point agarose (made in PBS) and it was immediately embedded on microscope slides. Cells were lysed (2.5M NaCl, 100mM Na2EDTA, 10mM TRIS; containing 1% Triton X-100) for 2h at 4°C. After lysis, the slides were placed in a horizontal gel electrophoresis tank containing fresh and chilled electrophoresis solution (1mM Na2EDTA, 300mM NaOH, pH>13) to allow DNA unwinding (20min), and electrophoresis was conducted for 30min at 26V, 300 mA. After electrophoresis, slides were washed in neutralization buffer (0.4M TRIS, pH 7.5) then stained with 85μl of EB (2μg/ml). Slides were analyzed immediately after staining at 80×magnification using a fluorescence microscope. One hundred randomly selected cells from each slide were scored using CASP image analysis software. The parameters used as a metrics of DNA damage were tail length (TL; µm), tail DNA (TD; %), and Extent tail moment (TM; arbitrary units). Tail moment was defined as: TM=TL×TD/100 [26].
Animals Survival Assay
20 mice were infected with 0.5ml of EAC cells in concentration of 0.5×108 cells per ml. Infected animals were inspected during 2 months, and animals death cases were recorded.
Statistical Analysis
Statistical analysis of the results was done using the statistical functions of the GrafPad InStat software (GraphPad Software, Inc., San Diego California USA). The one-way analysis of variance (ANOVA) and correlation analysis were performed on the data of biochemical variables. P-value of <0.05, <0.01, and <0.001 was considered respectively as statistically significant (*), very significant (), and critically significant (*). All data were expressed as mean ± standard error of mean (SEM). n=12 for in vivo and n=10 for in vitro experiments.
Results and Discussions
The dynamics of EAC cells growing in culture during 24 days were characterized by the following periods (Figure 2A): lag-phase- 1-3days; exponential growing phase- 4-12thdays; stationary phase- 12-16thdays; and phase of culture death- after 16thday of cultivation. Alive cells ratio (Tripan blue negative) had the tendency to gradual decreasing during first four days of cultivation: from 98% to 60%, followed by slight increasing to the constant level (near 80%) till the end of cultivation (not shown).
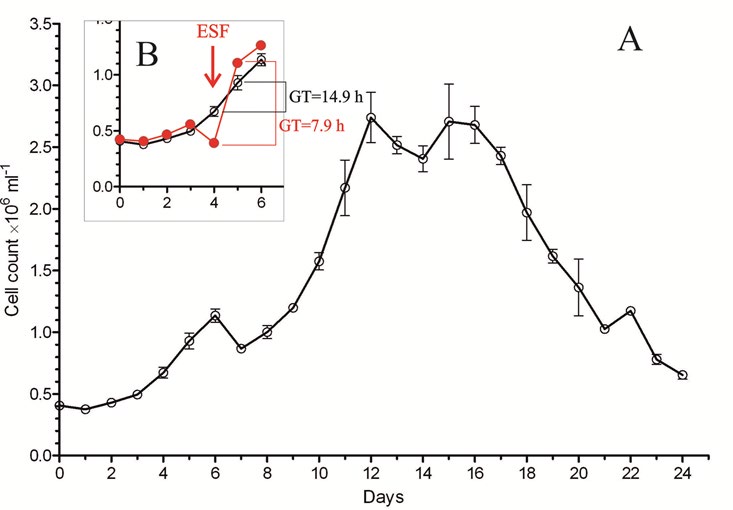
Observed periods of EAC growing in culture is in accordance with described in literature [27, 28]. However, an evident and unexpected decreasing of cell number on 7th, 14th, and 21stdays of cultivation was observed. This phenomenon is not described in literature for in vitro experiments, but there are some cases for collation concerning the in vivo experiments. Particularly, in PhD theses of Zamai AS, et al. [3] the observed character of in vivo growth of EAC cells was similar to that obtained in our in vitro experiment. Zamai AS, et al. [3] explained the data of her experiments by increasing the cell death (apoptosis) on definite day.
Aiming to find some explanation for the phenomenon of cell concentration descend at definite days of cultivation, we calculated the velocity of cell growth (number of duplications, n), using the following equation: n=3,32 log (cell number at next day/cell number at previous day). The curve for this parameter revealed a cyclic character of development of cultured EAC cells with the 7-days long period (Figure 3).
Particularly, the growing kinetics at 1-6days was almost the same as the kinetics at next 7-14days. After 14thday, the curve is deteriorated, but in general, saves the same character.
Besides, subtraction of a previous day alive cells number from the next day total cell number may reflect the number of cells divided at the previous day. The data in Figure 4 manifest, that at days 6, 13 and 20 the cells did not duplicate or proliferation was diminished. Actually, at these days the culture proliferation was arrested, therefore at next days 7, 14 & 21 the number of cells was dropped (Figure 4).
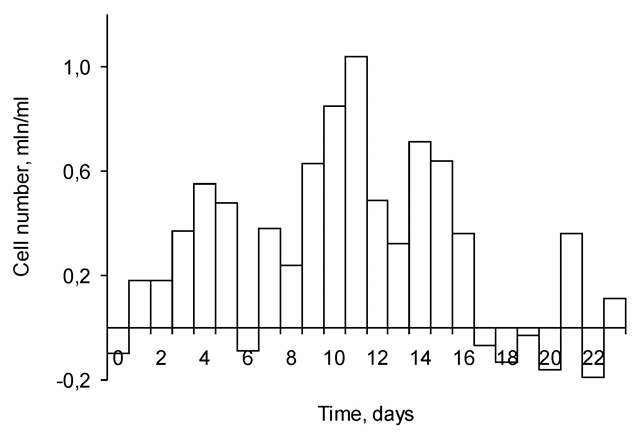
Hence, we inclined to explain the observed phenomenon of decreasing of number of cultivated cells at definite days by the cell cycle arrest at the preceding days. Thereafter, the survival of EAC-infected mice was monitored. The results (Figure 5) show the typical for EAC malignancy [29] high mortality rate of animals at the period of 10-14days of infection.
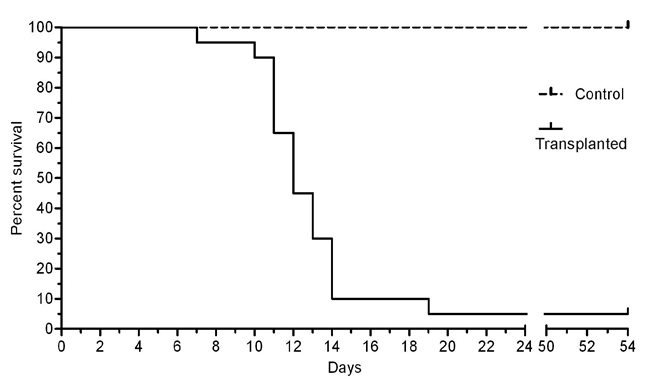
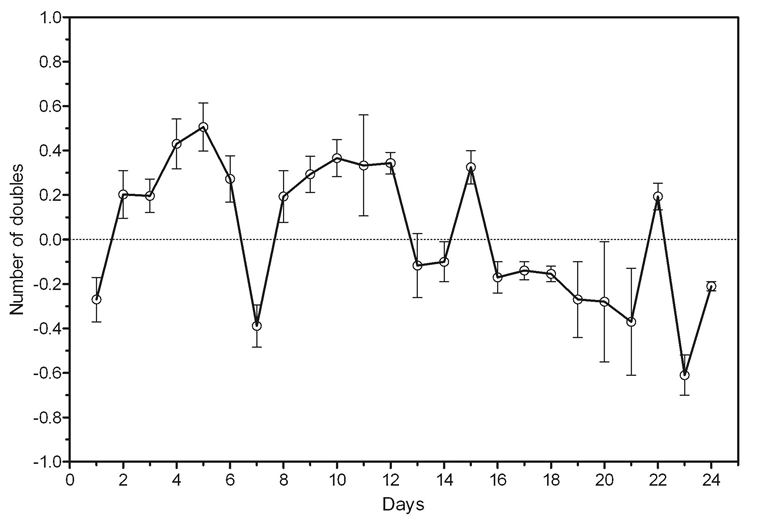
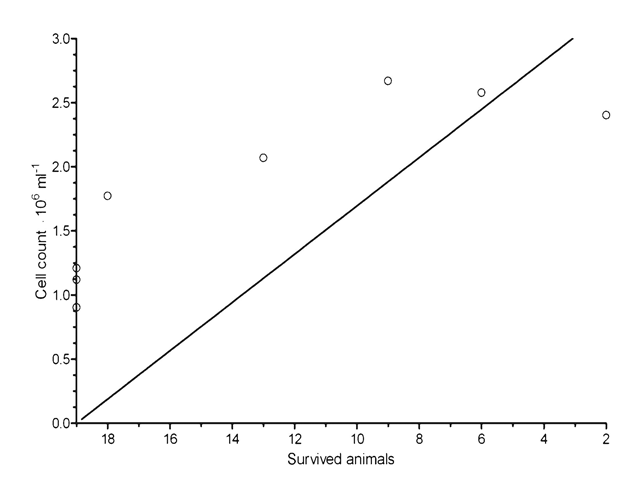
The time intervals of in vivo animal death and in vitro high cell concentration were coincided. The correlation curve for the number of cells in culture and the mortality of mice was analyzed (Figure 6). Negative correlation between those parameters was shown for the period of 7-15days (Spearman r=0.89; P = 0.0020**).
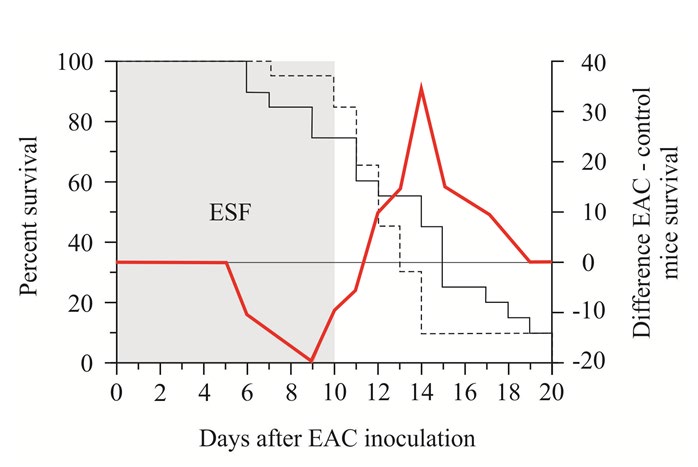
EAC cell infected mice exposed to ESF (during 10days, 6hours per day) characterized by higher mortality during period of exposure-till 11thday, and lower mortality-at the next days, compared to non-exposed control animals (Figure 7). It was proposed that ESF exposure bring to accelerated tumor growth, which brings to higher mortality rate at exposed animals. To clarify this hypothesis ESC cells were cultured and the culture were exposed to ESF (during an hour). EAC cell count drop (by 24.5%) was shown just after the end of exposure (Figure 2B). But on the next day the cell cont were higher than in the non-exposed control. For this day the generation time (GT) was almost twice higher (7.9hours) than in non-exposed cells (14.9hours).
Next, to prove the cytotoxic effect of ESF to EAC cells DNA-comet test was applied. Simultaneously, control leucocytes exposed to ESF also have been tested by Comet. We have shown earlier the genotoxic effect of ESF towards rats’ peripheral blood leucocytes (Figure 8) [30].
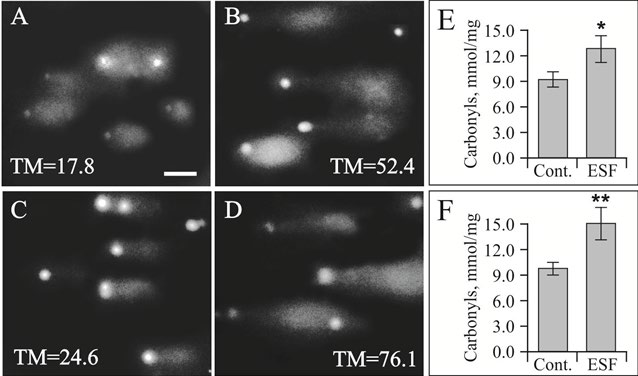
It was observed the genotoxic effect of ESF towards EAC cells was more pronounced (TM=76.1±9.4; P <0.01) than towards intact leucocytes (TM=52.4±12.3; P<0.05) (Figure 8A,B,C,D). The same situation shown for carbonyls. Protein carboxylation was measured to show probable way of ESF dependent cell damage. That is due to ESF derived oxidative stress. Indeed, ESF brings to substantial increase in protein carboxylation (by 30-40%). So facilitated tumor growth in ESF exposed animals might be a consequence of both EAC cell growth stimulation and host immune defense damage.
Conclusion
The EAC cells culturing was studied. The observed phases of cell growth are in line with the classical in vitro cultivation. The decreasing of cell concentration at definite days, and cyclic character of proliferation velocity (n) was shown. Negative correlation between animal mortality in vivo and cell growing in vitro was manifested. These results let us to assume that observed kinetics of growth of EAC cells is similar to the EAC fate in mice organism. This may be a substantial contribution to the knowledge of the tumor development in vivo, important in malignant treatment designing. Above it, we have shown that the ESF exposure facilitates EAC cell growth both in vivo and in vitro. Higher rate of EAC infected mice mortality might be ascribed both to the tumor growth acceleration and to the host immunity inhibition.
Acknowledgment
Author is grateful to Dr. Rafayel Muradyan head of laboratory of Toxicology of A. Mndjoyan Institute of Fine Organic Chemistry NAS RA for kind providing EAC tumor cells strain.
References
-
Müller M, Bockenheimer J, Zellenberg U, Klein N, Steger GG, et al. (2000) Relationship between in vivo drug exposure of the tumor interstitium and inhibition of tumor cell growth in vitro: a study in breast cancer patients. Breast Cancer Res Treat 60(3): 211-217.
-
Harutyunyan H (2010) New aspects of erlich ascites carcinoma development. The FEBS Journal 277 (S1): 56- 57
-
Zamai AS (2007) Regulation of proliferative activity of ascitic cells in Erlich Carcinom growth dynamics. Irkutsk.
-
Sauer H, Rahimi G, Hescheler J, Wartenberg M (1999) Effects on electrical fields on cardiomyocyte differentiation of embryonic stem cells. J Cell Biochem 75(4): 710-723.
-
Ahsan H, Ali A, Ali R (2003) Oxygen free radicals and systemic autoimmunity. Clin Exp Immunol 131(3)**:** 398- 404.
-
Knight JA (1995) Diseases related to oxygen-derived free radicals. Ann Clin Lab Sci 25(2): 111-121.
-
Waris G, Alam K (1998) Attenuated antigenicity of ribonucleoproteins modified by reactive oxygen species. Biochem Mol Biol Int 45(1): 33-45.
-
Lunec J, Holloway KA, Cooke MS, Faux S, Griffiths HR, et al. (2002) Urinary 8-oxo-2’-deoxyguanosine: redox regulation of DNA repair in vivo?**.** Free Rad Biol Med 33(7): 875-885.
-
Ames BN, Shigenaga MK, Gold LS (1993) DNA lesions, inducible DNA repair, and cell division: three key factors in mutagenesis and carcinogenesis. Environ Health Perspect 101(S5): 35-44.
-
MeConn J, Dietrich F, Rafferty C, Martin AO (1993) A critical review oft he genotoxic potential of electric fields. Mutat Res 297(1): 61-95.
-
Harutyunyan HA, Sahakyan GV (2016) Biological effects of electrostatic field: red blood cell-related alterations of oxidative processes in blood. International Journal of Biometeorology 60(1): 99-111.
-
Harutyunyan HA, Artsruni GV (2013) Biological effects of static electric field: plasma/serum proteome analysis of rats. Electromagnetic Biology and Medicine 32(1): 79- 94.
-
Harutyunyan H, Mkrtchyan V, Sukiasyan K, Sahakyan G, Poghosyan G, et al. (2015) Effects of in vivo and in vitro exposures to electrostatic field on some hematological parameters in rats. Bio Electro Magnetics 37(8): 513- 526.
-
Harutyunyan HA, Artsruni GG (2014) Investigation of genotoxity of the external electrostatic field of the higher tension by the method of DNA-comet (Arm). Proceedings of YSMU Scientific Report Cenference dedicated to 50th anniversary of Scientific Research Center of YSMU, pp: 55-60.
-
Harutyunyan HA (2015) Carbonylation of plasma proteins and blood hemolysis induced by the action of external electrostatic field In vitro. Medicinal Chemistry 5(5): 226-230.
-
MeConn J, Dietrich F, Rafferty C, Martin AO (1993) A critical review oft he genotoxic potential of electric fields. Mutat Res 297(1): 61-95.
-
(2014) Russia Statistic Population health care.
-
Shilo DS (2012) Worldwide electricity production dynamics. Minsk.
-
Aizenstein EM (2003) World market of textile raw materials: a look into the future. Light industry market, Morbidity by the main classes of diseases.
-
Artsruni GG, Zilfian AV, Azgaldian NR, Dovlatian RA (1987) Effect of an external electrostatic field on catecholamine secretion by rat adrenals. Kosm Biol Aviakosm Med 21(6): 67-70.
-
Levasseur M, Thompson PA, Harrison PJ (1993) Physiological acclimation of marine phytoplankton to different nitrogen sources. J Phycol 29(5): 587-595.
-
Westwood OMR, Hay FC (1980) Practical Immunology, 4th (Edn.), Blackwell Scientific Publications, Oxford, pp: 29-31.
-
Levine RL, Garland D, Oliver CN, Amici A, Climent I, et al. (1990) Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol 186: 464-478.
-
Venturi M, Hambly RJ, Glinghammar B, Rafter JJ, Rowland IR (1997) Genotoxic activity in human faecal water and the role of bile acids: a study using the alkaline comet assay. Carcinogenesis 18(12): 2353-2359.
-
Marczynski B, Rihs HP, Rossbach B, Hölzer J, Angerer J, et al. (2002) Analysis of 8-oxo-7,8-dihidro-2’- deoxyguanosine and DNA strand breaks in white blood cells of occupationally exposed workers: comparison with ambient monitoring, urinary metabolites and enzyme poymorphisms. Carcinogenesis 23(2): 273-281.
-
Sadettin SO, Bernhard P (1990) Effect of initial cell density on hybridoma growth, metabolism, and monoclonal antibody production. Journal of Biotechnology 16(3-4): 259-78.
-
Griffits B (1986) Growing of animal cells in culture. In Animal Cell Culture-A Practical Approach, Oxford- Washington DC.
-
Ingevatkin EV (2004) Manual for experimental oncology on the example of Erlich Ascites Carcinoma. Krasnoyarsk.
-
Harutyunyan H, Mkrtchyan V, Sukiasyan K, Sahakyan G, Poghosyan G, et al. (2016) Biological effects of electrostatic field: effects of in vivo and in vitro exposures to electrostatic field on some hematological parameters in rats. Bioelectromagnetics 37(8): 513-526.
- Cancer Diagnosis from RNA Sequence of Blood Cells by Using AI
- Field Cancerization in Oral Cavity, Case Report and Review of Literature. Oncologic Program Salud Integral Hospital, Managua, Nicaragua
- Identification of B Lymphocytes in Cancer Patient’s Blood
- A Case Report of a Breast Cancer Patient Developing Pneumonitis as a Result of Abemaciclib Therapy
- Immune Checkpoint Therapeutics for Today’s Fight and Beyond
- The Amalgamated Sophomore-Gonadoblastoma