An Update on the Immunotherapy Strategies for the Treatment of Type 1 Diabetes (TID)-How Far have we Reached in Reaching Insulin Independency in TID therapy-A Systematic Review
Type 1 diabetes mellitus (T1D) reflects a heterogenous autoimmune disorder which involves millions of subject’s worldwide. The basic characteristics of T1D are damage of the insulin generating β cells which takes place in view of abnormal activation of various immune effector cells. At present T1D treatment is done by lifelong delivery of novel kinds of insulin which have been synthesized recently. The aim of T1D care ,that has been guided by the Diabetes Control and Complications Trial (DCCT) are to get good glycaemic control, to avoid hyperglycaemia (as it is correlated with long-term microvascular as well as macrovascular complications) along with preventing recurrent hypoglycaemic episodes (as they adversely influence cognitive function). But even following repeated optimization of insulin therapy regimens, the hormonal substituted treatment only works to treat the symptoms without influencing the etiopathogenesis. New approaches which could influence the underlying modes responsible for β cells destruction have been evaluated in detail. These methods on the basis of immunotherapies have got incorporated within a panel of existing therapies for T1D, to block T cell responses against β cells antigens which are quite common at the time of initiation as well as development of T1D. But a total preservation of β cells mass along with insulin independency is not getting achieved despite massive exploration. Due to that right now no existing targeted immunotherapies are capable of replacing the standard insulin delivery. Currently a lot of interest has been concentrated on preventive methods in high risk subjects, on the basis of the posit that a therapeutic intervention once applied the early stage of disease, might aid in sustaining the endogenous β cell function by preserving the residual β cells reserve by the autoimmune attack. In this review we try to evaluate the present status of immunotherapies in T1D by emphasizing on the most significant studies in this field and detailing on novel methods that might get utilized to treat T1D in the future.
Introduction
Type 1 diabetes mellitus (T1D) is a condition that develops via the selective removal of pancreatic insulin generating beta cells brought about by an autoimmune defect. Subsequently, the basic properties of this disease develops at its advanced stage namely hypoglycaemia. This type of DM represents roughly 5-10% of all diabetic patients. Prevalence of this pathology points that >500,000 children suffer from T1D globally, that are located mostly in North America as well as Europe [1]. But the epidemiology points that the incidence of T1D has escalated rapidly in recent yrs [2]. In 2017, the International Diabetes Federation (IDF) declared 132, 600, newly diagnosed T1D cases all over the world. Earlier we had reviewed on the details of etiopathogenesis of T1D. Here we further explore how to get better ways utilizing immunotherapy that will obviate the need for insulin therapy [3, 4, 5].
Methods
We did a search using the PubMed search engine using the MeSH terms Type 1 diabetes (T1D); immunotherapies; antigen dependent; antigen independent; cell therapies; stem cell therapies; role of microbiota; innovative therapies like CAR-T cells or targeting JAK-IFN-pathways from 1980 till date in Apr. 2020.
Results
We found a total of 2463 articles out of which we selected 135 articles for this review. No meta-analysis was done.
Genetics of TID
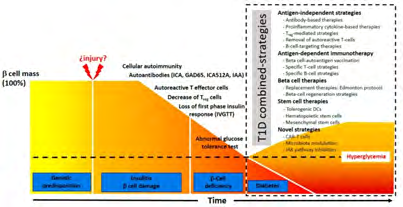
One of the main properties of T1D is the beta cell tolerance getting lost that is a process in which variety of factors participate [6], including genetic correlation with human leukocyte antigen (HLA) and various beta cell specific genes. T1D is thought to be an inflammatory disease where pancreatic islet infiltration with a lot of immune cell types (CD4+ and CD8+T cells), macrophages, dendritic cells (DC’s) as well as B cells play a major role [7]. This propagation of islet infiltration facilitates removal of beta cell which finally causes diabetes initiation. Although few advantages are there, the transplantation of pancreas or pancreatic islets (Edmonton protocol) [8] have given minimal efficacy in view of inadequate quantity of donors with the reactivation of the autoimmunity status inspite of immunosuppression protocols. Further pancreatic transplants have been shown to be just partly successful [9].
Immunological Modes in T1D Aetiopathogenesis
Propagation of type 1 diabetes (T1D) is broken in 3 key stages [10] i) At 1st stage that might go through a long period of time, persons form β cell autoimmunity, checked by serum autoantibodies detection. The commonest autoantibodies in T1D patients are the ones against glutamic acid decarboxylase (GAD) (GAD 65), the tyrosine phosphatases, IA-2 and IA-2β, zinc transporter8 (ZnT8) and insulin [11]. These epitopes might lead to stimulation of CD4+ and CD8+T cells that are the major ones bringing about the β cell damage.
Diabetes autoantibodies have a critical part in detection of preclinical stages of T1D. The Trial Net TN01 has evaluated the significance of autoantibodies markers for identifying diabetes mellitus (DM) [12]. 5% of the people examined in this study were observed to have blood autoantibody. This study also found that 95% of patients who extend to symptomatic T1D had positive autoantibodies by the age of 5yrs [13]. Finding the autoantibodies in the TEDDY (The Environmental Determination of Diabetes of the Young) study demonstrated a peak within 2 and 9yrs of age [14]. People showing a minimum of 2 separate autoantibodies had a marked chance of forming T1D [12, 15].
Further, separate HLA haplotypes were found to be protective or predisposed to DM formation [16]. At the time CD4+ and CD8+T cells started to destroy β cells, the insulin amounts start to reduce, that starts the ‘’second stage’’ of the T1D. At this juncture the basic approach for T1D therapy would aim to repress β cells autoimmunity as well as protecting the rest of the β cells mass persisting. Various studies have shown that at the time of diagnosis that overlap with the 2nd stage still remaining β cells present (Clinical Trial NCT 01030861) [17]. Delivering immunosuppressive drugs in children with new onset of T1D can postpone or reverse the propagation of, but immunosuppression could also end in organ toxicity. The DM propagation restarts after the therapy is withdrawn [18].
The ‘’third stage’’ of T1D takes place in long time patients. At this particular stage, basic aim is to make sure that the functioning of whatever β cells, are there, and persists. The studies have revealed that after the onset of disease marked decrease in C-Peptide levels, a short peptide which connects insulin’s chains in the pro insulin molecule and can be utilized as a surrogate of how much insulin gets formed (Clinical Trial NCT 01030861) [17]. Ensuring the sustenance of a large β cells number might aid in the regulation of hyperglycemia along with decrease the comorbidities of the disease. In human cadaveric T1D pancreata, CD4+ and CD8+ T cells, macrophages, as well as B cells have been illustrated [12, 19]. But the absence of insulitis in certain cadaveric T1D pancreata samples emphasizes the heterogeneity of the disease [20] that might be one of the explanation, why immunotherapies have not been fully efficacious in T1D cases.
Various immunotherapies have been advocated for all 3 stages of T1D cases. One strategy uses the modulation of the immune response by utilizing antibodies which target particular immune mediators. Other strategy takes the aid of β cell antigen specific therapies. Intriguingly, a therapy on the basis of oral insulin delivery shows a delay in the DM onset in non-obese diabetic (NOD) mice [21], an animal model that has been used heavily for evaluating the propagation as well as pathogenesis of T1D.
Animal models of T1D
2 separate animal models have usually been utilized in T1D research. The NOD mouse as well as bio breeding (BB) rat. Both of these models display the major DM symptoms, namely glycosuria, polyuria, weight loss, as well as lymphocytic infiltration of islet of Langerhans [22]. But in view of the involvement of T cell compartment in the etiopathogenesis of T1D, the NOD mouse model has been considered better and has an edge over BB and hence utilized for the study of the diabetogenic T cell formation [23]. NOD mouse displays similar properties to human DM. NOD mice were initially developed in the Cataract Shionogi (CTS) strain [22] Cell invasion in the pancreas of NOD) mice can be seen as fast as 3wks of age This has the recruiting of various innate immune cells into the islets of Langerhans having macrophages as well as neutrophils, before the lymphocytic infiltration [24]. Despite low autoreactive cells present to start with, numbers escalate slowly, in view of some DM particular autoantigens get recognized as well as get stimulated, starting the removal of insulin manufacturing β cells. Even though concentration of T1D research has been on T cell stimulated β cells damage, studies have been done revealing that B cells also play a key role in the DM initiation [25].
Lot of essential data has been received through the NOD mouse model as far as part of the immune cells in DM formation is there. Moreover NOD mice have given a special opportunity for evaluation of immunotherapies (like CTLA4-Ig, as well as anti-CD40 antibodies, as well as IL-4 or IL-10 therapy)-reviewed in detail BY Shoda, et al. [26].
Problem lies that most immune therapies showing benefit in NOD mouse model did not show similar results in human disease. In view of this, trying to humanize NOD mice [27], might aid the research work that would ultimately translate into useful immunotherapy clinical trials. Further some external factors might also have their part in T1D formation. In monozygotic twins work done demonstrated absence of concordance pointing to the role of environmental factors in T1D propagation. A lot of these factors modify the DM susceptibility of NOD mice that are alterations in gut microbiota (GM) [28]. Innate immune components cross reacting with the GM is a burning topic in T1D research.
Besides the above mentioned mouse model, another one of use is the DO 11.10XRIP mOVA (DORmO) mouse model, where RIP mOVA animals (mice which express membrane bound OVA in thymus and pancreas) are crossed with DO 11 animals that express an OVA –particular MHC-II TCR. Surprisingly these double transgenic animals develop a huge number of islet specific functional Treg cells, but develop T1D spontaneously by wk 20.Thus, the DORmO model is specifically good to evaluate the part of Treg cells in T1D initiation/propagation [29].
Current Status of T1D immunotherapy
Current Status of T1D immunotherapy might be divided as
- Antigen-independent.
- Antigen dependent.
Antigen-independent or non-antigen specific therapies are
- Drugs which cause immunosuppression.
- Antibody –based treatments which aid in the deletion of polyclonal B or T cells [30].
- Cytokines based therapies [31], and the escalation of tolerogenic DC [32], as well as polyclonal Treg cell numbers [29].
Antigen- specific therapies are the ones that
- Utilize β cell-derived auto antigen dependent vaccines
- Adoptive transfer methods as well as
- Specific amelioration of autoreactive T cells clone by attacking Antigen-presentation modes [33]. The combination of various methods on the basis of immunotherapy treatments are thought to be the most efficacious approach in view of the complicated nature of T1D [34].
Antigen-Independent Approaches Antibody-Based Treatments: Different costimulatory pathways regulate the stimulation of T cells that might be positive or negative. Like signaling via CTLA4 causes an anergic situation in naive - T cells, and hence abatacept, a fusion protein made up of the Fc part of the human Ig G1 that is fused to the extra cellular domain (ECD) of the CTLA4 is utilized for therapy of rheumatoid arthritis [35]. Abatacept, in a recent trial clinically showed its property against T1D by postponing C-peptide depletion in T1D patients [36]. The clinical trial Trial Net is evaluating the advantages of Abatacept in postponing of early T1D onset ((Clinical Trial NCT01773707; www, clinicaltrials.gov).
Anti CD3 monoclonal antibodies which attack CD3/ T cell receptor (TCR) complex, blocking the union of CD3 with TCR and stimulating the development of an anergic state of the T cells have also been analyzed in T1D patients. Teplizumab as well as otelixizumab that are 2 of the major clinically recommended Anti CD3 antibodies have shown some efficiency in T1D patients [37]. Postponement of C-Peptide reduction in T1D treated patients was stimulated by Teplizumab. In the study AbATE, 2 wks Teplizumab therapy caused C-Peptide to get preserved [17] Clinical Trial NCT 01030861; www, clinicaltrials.gov).]. Main results of this Clinical Trial were anticipated in end of 2019.
Of the specific amelioration of T effector cells, deletion of memory T cells would also be essential for getting long- standing outcomes. Inhibition of CD2 signaling would give this objective. The anti- CD2 fusion protein Alefacept effectively blocks T cells getting activated stimulating apoptosis of both memory as well as effector T lymphocytes. Although, just a mimimal trend regarding preservation of C-Peptide levels was attained on checking this posit in the clinical trial TIDAL, comprising of patients at last stage of T1D [18]. Finally anti thymocyte globulin (ATG) has been detailed for having the capacity to eliminate stimulated T cells. Used in low doses ATG along with granulocyte-colony stimulating factor (G-CSF) demonstrated it is safe and could lead to protection of β cell mass [38]. Last clinical trials illustrated that G-CSF alone did not give any extra advantages [39].
Proinflammatory Cytokines based Therapies: It has been known for long that, inflammation as well as Proinflammatory cytokines participate in T1D formation [40]. Expression/inhibition of these molecules might stimulate essential alterations in pancreatic β cells [41]. This particular approach was utilized clinically to treat other autoimmune diseases [40]. Interleukin (IL)-1α as well as IL- 1β are essential immune modulators that monocytes express on β cells [42]. Delivering anti- IL-1α in rheumatoid arthritis is well tolerated in patients [43]. Further IL-1α causes propagation of T1D via stimulation of T helper cells, as well as escalating the quantity of circulatory memory T cells [44]. Another clinical trial performed in T1D patients pointed that IL-1 inhibition might stimulate preservation of pancreatic β cells [45].
TNF-α is the other cytokine acting as an intermediate molecule in autoimmune disease. Due to that TNF-α block has been evaluated for therapy of autoimmunity. Those T1D patients who received Etanacerpt (recombinant TNF-α- receptor –Ig G fusion protein) did improve the preservation of pancreatic β cells mass (as checked by C-peptide amounts) as well as reduced glycate Hb amounts [46]. The property of nicotinamide alone or with Vit E for conserving the functionality of residual β cells mass has been evaluated too. Both therapies helped in conserving the basal C-peptide secretion [47]. The IL-12/23 cytokine pathway, involved in inflammatory cytokines stimulation as well as pathogenic T cells stimulation was also evaluated as a therapy for T1D. Ustekinimab (IL-12/23 blocking molecule) use has been examined in T1D patients (USTID Clinical Trial (Clinical Trial NCT 02117765; www.clinicaltrials.gov). In a subset of cases of T1D, overexpression of IL-6 was found [48]. Secondary to this anti- IL-6 treatment that is also evaluated in therapy of arthritis as well as systemic juvenile idiopathic arthritis [49] was started. Right now the Clinical Trial EXTEND (Clinical Trial NCT02293837; www.clinicaltrials.gov) is evaluating if IL-6 signaling blocking (toclizumab, an IL-6 -receptor antibody) can stimulate a protection of β cells function in T1D patients (6-17yrs age) is being conducted. Considering total data, cytokine inhibition is a viable method that is supplementary for attaining a durable therapy that is efficacious for treating T1D.
Treg Mediated Approaches: In the pathophysiology of T1D Treg involvement has been identified [50]. The part of expanded autologous polyclonal Treg in therapy of T1D patients was evaluated by Bluestone, et al. [50]. Autologous polyclonal Treg infusions were detected to be safe in this Clinical Trial, but could not change the course of the disease. Separate Clinical Trials have also been examined regarding the actions of low doses of IL-2 on Treg action [51]. Despite escalation of total amounts of Treg by IL-2, it didn’t influence a good glycaemic control. Interestingly, recent literature demonstrated that blocking extracellular deposits of the polysaccharides hyaluronan (HA) (Commonly seen in T1D patients [52]) decreased DM in 2 separate mouse models by significantly increasing the percentage of Treg in pancreatic islets as well as avoiding anymore β cells damage [29].
Elimination of Autoreactive T cells: Autoreactive T cells Elimination in T1D patients by targeted removal is a strategy having lot of potential, since its objective is to finish effector T cells that causes the pancreatic β cells damage. Therapy of NOD mice utilizing anti-CD3 antibodies was demonstrated to stimulate energy in T cells [53]. Further increased large amounts of Treg cells were seen in patients given anti-CD3. Decreased insulin needs following therapy with anti-CD3 antibodies was seen in a clinical trial. These data point that anti-CD3 antibodies therapy can be thought of as a potential therapy for treating T1D [53].
B cell Targeting Therapies: As B Cells were believed to be involved in β cells damage via autoantibodies synthesis, B Cell Targeting has also been evaluated in T1D. Deletion of B Cells in NOD mice avoided the collection of autoantibodies, hence preventing the DM initiation [54]. T1D patients receiving anti- CD -20 antibodies displayed > C-peptide amounts as well as lower insulin reliance as compared to placebo group. But this method does not appear to totally avoid C-peptide reduction [55, 56].
Antigen-dependent Approaches: Rather than Antigen- independent Approaches, autoantigen-attacking therapies of T1D might modify, particularly T1D-associated autoimmunity, whereas maintaining the normal immune homeostasis. The basic aim of antigen-attacking therapies is for stimulating tolerance of autoreactive T effector cells as well as expansion of autoantigen –specific Treg cells [33].
Beta Cell autoantigen Vaccine: Once particular antigens get exposed to naive Tcells can stimulate tolerance to that antigen. As per the present literature of T1D, one can posit that antigens formed from β cells which were applied in a non-inflammatory situation, might modify autoreactive T cells, causing β cells conservation [57]. On the basis of this innovative vaccination approaches have been for attaining Tcell-tolerance against particular autoantigens The properly understood Tcell epitopes against insulin as well as glutamic acid decarboxylase (GAD) have been deeply evaluated [58], showing that C19-A2 proinsulin peptide might manipulate autoreactive CD4 + T cells in patients with particular class II allele [59]. Delivery of this peptide in newly diagnosed T1D pts lead to showing > C-peptide amounts without symptoms of systemic or local hypersensibility [60]. Further another T1D autoantigen, GAD65, was targeted in NOD mice for decreasing the amount of GAD65-specific T effector cells [61]. Normoglycaemia was obtained in 70% of NOD mice, as well as in 80% of them this Normoglycaemia continued in long time post antigen delivery.
Inspite of the beneficial data seen with vaccination approaches in NOD mice, the differences in autoantigen, between human as well as mice as well as heterogeneity of T1D in humans makes this approach not very useful for clinical situations [62].
Specific Tcell Approaches: The imbalance of Treg to T effector cells is an essential factor that decides the onset of T1D [63]. CD8+T cells stimulation gets modulated by the presentation of specific epitopes from professional APC s since DC’s seem to be the main APC s for the CD8+T cells [64]. This process is based on CD4+ T cells crosstalk which stimulates the particular subsets of CD8+T cells that in turn causes islet cell damage [65]. This method of attaining self-tolerant T effector cells can be via utilization of whole antigen or particular peptides. But success in tolerization of T effector cells is based on various factors particularly the finding of the autoantigen which drives the process. For avoiding the β cells damage, the most important T effector clones need to be eliminated.
Specific B cell Approaches: This is based on the amelioration of nonspecific B cells, although not proved to be very efficacious. But, still inhibiting particular autoreactive B cells by depleting insulin reacting B cells is a promising other method [66]. In the NOD mice insulin specific B cells evade the immune control, responding to insulin by enhancing the expression of costimulatory molecules at the time of cross priming of effector T cells.
β Cell Treatments
Edmonton Protocol-Replacement Therapies
The importance of islet transplantation has been demonstrated by Edmonton Protocol by addressing insulin control in T1D patients [7]. As per this protocol, pancreatic islets received via cadaveric donors get infused in immunosuppressed T1D patients. Although trials carried out prior to 1990 utilizing single islets infusion had part success, decreasing insulin requirements as well as >C peptide amounts, but no further work was done for escalating the net islets mass of the transplant in any of the trials [67]. These islet transplantation protocols developed worked out to be a beneficial treatment of T1D in view of the Edmonton Protocol getting introduced in 2000. Currently this remains the only method that can attain glycaemic control without delivering any exogenous insulin [68]. Advantages of only pancreatic islets possesses various benefits over transplantation of a full pancreas, as this needs only a very minute surgical method having low morbidity as well as mortality, with a significantly <cost. The biggest edge of islet transplantation protocols above the conventional insulin treatment is that transplanted islets have >efficiency in sustaining normal blood glucose amounts without synthesizing extra insulin which could result in hypoglycaemic episodes.
Modifications of the Edmonton Protocol on the basis of new immunosuppression regimen have avoided the utilization of corticosteroids that allows the use of a particular combination therapy on the basis of antiinterleukin-2-receptor antibodies in addition to immunosuppressant drugs siromilus as well as tacrolimus. Biggest benefit of this combination therapy is low β Cells toxicity. Islet transplantation has demonstrated certain success as far as insulin independence both in the short as well as long term [69, 70], with the variability of results got with Edmonton Protocol is related to both the organ donor as well as the recipient. Despite the advantages of islet transplantation protocol are accepted, the concerns are standardization of this strategy are the >amounts of islets which need to be transplanted as well as the side effects secondary to immunosuppressive regimen. The 1st difficulty might be overcome by utilizing stem cells which under proper differentiation protocol, can differentiate into glucose sensitive insulin synthesizing cells.
β Cell Regeneration Approaches
Gastrin and Glucagon like peptide -1(GLP1) possess a synergistic action stimulating the regeneration as well as differentiation of β Cells [70, 71]. Adding both these molecules in the NOD mouse model, lead to an escalation of β Cell mass [72]. Further, the combination therapy utilizing dipeptyl- peptidase 4(DPPIV) inhibitors (for enhancing GLP1 amounts) as well as proton pump inhibitors (PPIs-for escalating Gastrin amounts), resulted in escalated C peptide amounts as well as insulin liberation, restoring the normoglycemia in NOD mice [70]. In humans REPAIR-TID study evaluated the action of 1yr of similar therapy utilizing a combination of sitagliptin (DPPIV inhibitors) with lansoprazole (PPIs) in T1D patients [73]. But no variation in C peptide amounts were visualized in the treated visa Vis placebo groups [73]. The authors presumed that the enhancement of Gastrin amounts as well as GLP1 were low, causing noneffective therapy. More clinical trials will be needed.
Stem Cell Therapy Methods
Tolerogenic DC’s: Despite different cell types being evaluated as probable targets for T1D therapy, dendritic cells proved to be of interest. But clinical trials where T1D patients got autologous DC’s demonstrated minimal outcomes. In these clinical trials, DC’s got infused through abdominal intradermal injections after 2 wks repeatedly [74]. Despite the therapy getting well tolerated, no significant variations in glycaemia were seen. Earlier studies showed that DC’s, alone or through various effector cells like Tregs as well as Bregs might have an essential part in the activation status of autoreactive CD8+cytotoxic T cells (CTL) along with effect the balance among T helper cells (Th1 as well as Th2) as well as effector cell populations [75]. Tolerogenic DC’s (t DC’s) populations have been utilized in clinical trials for therapy of autoimmune diseases including TID [74, 76]. The outcome of those studies pointed that t DC’s persist at the delivery site facilitating the production of a lymphoid stroma tissue that in turn helps in enhancing FoxP3+Tregs [77].
The synergistic inter-relationship of t DC’s as well as Tregs, would help in stabilization of FoxP3+expression and would increase the amounts of IL-10, TGFβ, as well as retinoic acid by t DC’s [78]. The Tolerogenic state of the t DC’s would be enhanced through cell-cell interactions or via paracrine mechanisms. This combination strategy might alter the paradigm of how autoimmune diseases are getting treated, addressing the disproportion of the immune effectors developed during the disease onset.
Haematopoietic Stem Cells (HSC): Despite immune dysfunctions associated with T1D are complex, Voltarelli, et al. [79] reported a novel work in which newly diagnosed T1D patients were included in a phase1/2 clinical trial got immunosuppression therapy in addition to infusion of autologous HSC’s. Promising outcomes were documented, practically all pts did not need insulin injections for 6 mths since their C peptide amounts remained stable as well as the ant-GAD amts got reduced [79]. Maximum patients in 2 recent prospective non-randomized trials displayed ne need for insulin injections following HSC transplantation [80]. The outcome of these studies displayed that even after 4yrs post transplantation, the C peptide amounts remained significantly > than pre-transplant ones [80]. The outcomes of a study that utilized autologous non myeloblastive HSC transplantation got published [81]. 59% of the patients who got included in this clinical trial did not need insulin injections whereas 32% remained insulin free for a minimum of 4yrs [81]. Maximum pts enrolled in the autologous HSC’s transplantation clinical trials showed minimal side effects. In only 1 clinical trial a patient was reported to die secondary to pseudomonas aeruginosa sepsis [80]. Despite the side effects associated with immunosuppression protocol reduce this alternative therapy, the delivery of autologous HSC’s is an exciting method to find a cure for T1D.
Mesenchymal Stem Cells (MSC): Mesenchymal Stem Cells (MSC) represent stromal cells which carry out essential roles in tissue repair as well as regeneration [82]. Specific antigen biomarkers get expressed by MSC’s like MHC1, CD90, CD105 as well as a CD73 that helps in identifying them via flow cytometry methods. In regenerative medicine MSC’s have turned out to show a lot of advantages in view of their capacity to produce various cell types like adipocytes, chondrocytes, as well as osteoblasts helping in replacing injured tissues [83]. Additionally MSC’s can get recruited from other damaged tissues, like ischemic heart or pancreas [83]. Due to this, MSC represent a new method which will aid in integrating stem cell transplants in regenerative medicine protocols [84]. MSC’s have been tried for therapy of T1D patients and displayed good results in maintaining blood C peptide amounts [85]. But no variations were seen regarding insulin needs as compared to non-treated group during the study.
Biological characteristics of MSC’s as far as their role in regulating abnormal immune responses were shown in NOD mouse model [86]. In a clinical trials that got sponsorship from Uppsala University Hospital, where T1D patients got transplanted with autologous MSC’s treated patients showed a better mintainance of C peptide amounts [86]. Umbilical cord blood MSC’s (UC-MSC’s) were also evaluated in combination with autologous mononuclear cells derived from bone marrow (ABM-MNC) in a different clinical trial. In fusion of ABM-MNC caused a 30% decrease in insulin needs [87]. A lot of trials are evaluating the use of MSC’s vis various sources to treat T1D that includes the utilization of allogenic MSC’s derived from adipose tissue (NCT 02940418 as well as NCT 02138331). Use of immune regulatory MSC’s till date is proving to hold a lot of promise in the T1D Stem Cells field. Combining MSC’s with other immunotherapies would give innovative ways for treating T1D patients.
Innovative Methods
CAR-T Cell Method
Introduction: Recently an immunotherapy where Tcells that express chimeric antigen receptors (CARs) that are specific against CD19 turned out to be a big breakthrough for cancer therapy of CD19+B-cell leukemia [88]. CARs represent complicated molecules made up of different components, commonest ones are
- An antigen-specific recognition domain, mostly a single chain variable region (scFv) from a monoclonal anti body.
- A hinge area, on the basis of Fc part of a human IgG (IgG1 or IgG4), or initiating from the hinge domains of CD8a or CD28.
- A transmembrane domain as well as
- An intracellular tyrosine-based signaling domain [89].
This signaling domain represents the engine of the receptor. The commonest part is the intracellular part of CD3 that is the major signaling chain of CD3- Tcell receptor (TCR) complex. The maximum benefit of CAR-T Cell being that the receptor’s crosstalk with its antigen is independent of the MHC yet it still stimulates the same TCR’s as well as costimulatory intracellular signaling cascades essential for Tcell activation as well as expansion.
CAR-T Cells as well as T1D: On the basis of studies carried on utilizing CARs in cancer as well as >interest of Treg as a potential tool for T1D therapy. One can posit that connecting Tregs with β-cell specific CARs would improve Treg‘s migration into the pancreas as well as pancreatic lymph nodes, hence preventing autoimmune damage of islet cells. Lot of recent studies point that a big possibility of use of CAR-Treg treatment in multiple autoimmune or allograft rejection model systems [90, 91, 92, 93, 94]. An intriguing method regarding utilization of CAR-Treg in the EAE-, mouse model was suggested by Fransson et al. [93]. CD4+T Cells got engineered for expressing both i.e. a CAR specific against myelin oligodendrocyte glycoprotein (MOG35-55) as well as a murine Foxp3 gene for stimulating Treg differentiation, distanced by a 2 A peptide sequence in their study. Intranasal delivery of CAR-Treg lead to a successful administration to the CNS, an effective repression of the ongoing inflammation as well as complete recovery through disease symptoms. Other studies posit the use of CAR-Tregs in transplant rejection by developing HLA-A2-specific CAR-Treg, which were obtained via the host [91, 92]. These HLA-A2- CAR- Treg had great expression of Foxp3, LAP, GARP as well as CTLA4 retained besides sustaining their suppressive function in vitro without a significant cytolytic activity. Despite necessity of corroborating the stability of Treg phenotype, purity, as well as long term survival following the transfer, this approach holds great promise for therapy as well as prevention of transplant rejection by stimulating graft particular tolerance.
These CAR- Tregs were also examined in Haemophilia A, in whose case genetic mutations in F8 gene end in either decreased amounts or changed functionality of the blood clotting protein, factor VIII (FVIII). In cases of severe Haemophilia (no circulating FVIII can be found), high chances for generating bad immune reactions to the externally delivered FVIII protein. Markedly delivering FVIII-specific human CAR-Tregs repressed antibody synthesis in vitro as well as in vivo in a mouse Haemophilia A model. But as FVIII is a soluble protein, the mode of repression is not exactly known [91, 95]. Besides that the study via Hansen’s group gave extra evidence of the concept that CAR- Tregs are a prospective therapy method for multiple autoimmune conditions [94]. They developed CAR against carcinoembryonic antigen (CEA), a glycoprotein that is present on lung adenoepithelia, and then adoptively transferred Tregs expressing the construct in an experimentally stimulated allergic asthma mouse model. The CAR- Tregs collected in the lungs and lymph nodes that are near, decreasing airway hyper-reactivity, inflammation, mucus synthesis as well as eosinophilia.
Problems: Inspite of the lot of potential of CAR- Tregs therapies, there is no clear approach as to how to use this exciting technology for T1D therapy. The main problem is the absence of β-cell specific antibodies that can be gathered to develop islet protective CAR- Tregs. 1 approach for overcoming this challenge is utilizing human islet specific TCR gene transfer to polyclonal human Tregs. In a recent study where polyclonal human Tregs were transduced with TCR chains obtained via 2 human islet specific CD4+ clones demonstrated an improved antigen-specific repression of these cells as well as enhanced potency as compared to polyclonal Tregs [96], But these islet specific Tregs were <responsive to their cognate antigen as compared to Tcells expressing virus-specific TCR’s point that more optimization/ or finding better TCR’ clones is still required.
A new study showed that insulin-specific CAR-Tregs were functional, suppressive as well as surviving in vivo despite their being unable to prevent spontaneous DM in NOD mice [97]. This does not come as an astonishing fact knowing the fact that insulin is a soluble antigen which is present throughout the body, with its levels fluctuating. Further such a method won’t be very effective in T1D patients where endogenous insulin amounts are very little, with daily insulin injections would disturb the normal insulin gradient which might drive the insulin-specific CAR-Tregs into the pancreas.
Hence the discovery as well as study of new β-cell specific molecules which could give proper targeting of CAR- Tregs is required. Although there are specific molecules that are promising like DPP6 [98], FXYD2ƴa [99], as well as NYTPDase 3 [100], all of these would need extra studies to confirm their specificity along with finding proper monoclonal antibodies which would recognize human β-cells in vivo prior to generating a CAR construct for T1D therapy.
Summary: Inspite of advances in the field of CAR- Tregs therapies with their >potential of use in autoimmune disorders, still there is absence of an effective system along with β-cell specific markers which lets the formation of efficient autoantigen specific Tregs which might be used for cell based therapies in T1D.
Modulation of Microbiota
It is well known that both small and large intestine house >a trillion microorganisms that belong to over 100 species. Changes in intestinal bacteria has a role in development of obesity and glucose tolerance along with NAFLD has been proven [reviewed in 101-107]. In case of TID more important publications in both mice models as well as humans benefit of Akkermansia Muciniphila administration insulin sensitivity as well as glucose homeostasis, healthier lipid profile with a proinflammatory tone besides other changes [108]. Giongo, et al. [109] used samples from 8 Finnish children of which 4 cases later developed T1D with rest 4 being controls showed that the case children’s samples had an unsatisfactory formation of GM diversity, that did not become as complex as that of controls as well as had > heterogeneity among cases. They emphasized on the significance of a compromised phylogenetic diversity in a risk of forming autoimmune DM and lay down the basis of potential screening criteria. This was corroborated by a Chinese study [110]. Long cohort studies, and RCT like FINDIA (Finnish Dietary Intervention Trial for the Prevention of type 1 diabetes), BABYDIET (in German infants), TRIGR (Trial to Reduce IDDM in the Genetically at Risk) and TEDDY of other lots gave important knowledge as to the natural history of T1D along with how GM participate [reviewed in 111]. More information about enriched intestinal segmented filamentous bacteria (SFB) was provided by Krigel, et al. [112] regarding formation as well as propagation of DM in NOD mice. Though a protective role of SFB’s could not be assumed, their conclusions were that SFB’s in certain ways ameliorated the propagation of T1D as well as facilitate a boost in certain T helper cell sub-populations. Earlier SFB’s were thought to be latent but present proof gives clues that they; possess part in mucosal immunity as well as immune response.
Summary: From these observations it is clear that GM should not be ruled out regarding management of T1D. With the information present, mainly the protective human studies. Point to a major part of GM in the risk as well as formation of autoimmune disorders. Trying to find specific targets in the GM would aid to increase the efficacy of these innovative methods and give diabetic patients alternative medical therapy.
Inhibition of JAK Pathway
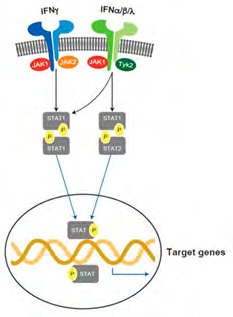
Introduction: Mammalian Janus Kinase (JAK) family consists of 3 JAK’s (JAK 1,2,3) as well as tyrosine kinase2 (TYK2) that binds separate receptor chains [113]. On binding of ligand to its cognate receptor, related JAK’s get activated and phosphorylated, that makes docking sites for the SH2 domain of the cytoplasmic transcription factors known as signal transducers and activators of transcription (STATs). There are 7 STATs in human STAT family, namely STAT1, STAT2, STAT3, STAT4, STAT5A STAT5B, STAT6. After phosphorylation STATs get translocated to the nucleus, dimerize, and the bind to particular DNA sequences for regulation of gene transcription [114]. The JAK-STAT pathway plays a key role in the downstream signaling of inflammatory cytokines like IFNs, ILs as well as growth factors [115]. TID as well as JAK: A type 1 IFN signature antedates the finding of autoantibodies in children genetically at risk for TID [116] as well as IFN-α gets expressed in human islets from TID patients [117]. MHC class 1 overexpression is induced by IFN-α [118] as well as IFN-ƴ [119] in human islets from TID patients and IFN-α further induces β-cell ER stress as well as chemokine synthesis [120]. Receptor engagement by IFN-α triggers JAK-TYK2 heterodimer signaling (Figure 1). TYK2 has been correlated with various autoimmune diseases that include rheumatoid arthritis as well as TID [121]. Six TYK2 single nucleotide polymorphisms (SNP) (rs34536443, rs2304256, rs280523, rs280519, rs12720270, rs12720356) have been evaluated in association with autoimmunity. Importantly the SNP rs2304256 causes a missense mutation in TYK2, and has been observed to be correlated with protection against TID [121].
Downstream IFN-α/ IFN-ƴ signaling is STAT1 dependent (Figure 2) as well as STAT1 gets overexpressed in islets from TID patients with strong correlation with HLA class 1 expression in β-cells [118]. IFN-ƴ also has a role in expression of the CXCL10 that appears to be activated in islets from both TID patients [122] as well as NOD mice [122]. CXCL10 aids in pathogenic T cell infiltration into the pancreatic islets resulting in β-cells apoptosis as well as its neutralization avoids DM in NOD mice [123]. Absence of IFN-ƴ delays the propagation of autoimmune diabetes in NOD mice [124]. Current proof further corroborates this IFN- stimulated JAK-STAT pathway activation markedly adds to TID pathogenesis. Patients having STAT 3 gain of function germline mutations, being predisposed to TID having a median age of onset of 8weeks. Moreover, about 15% of patients treated with immune check point inhibitors develop endocrine autoimmunity [125], that includes pancreatic β-cells targeting [126], that results in TID [127]. Corroborating these findings, inhibition of PD1- PDL1 signaling escalates DM in NOD mice [128].
Earlier therapy with of in vitro human islets with ruxolitinib (JAK1/2) markedly decreased IFN-α mediated inflammatory as well as ER stress markers [129]. Further therapy of NOD mice with a JAK1/ JAK2 inhibitor (AZD480) blocked MHC class 1upregulation on β-cells and reversed autoimmune insulitis by decreasing immune cell infiltration into islets in newly diagnosed animals [130]. Lastly, pancreas specific genetic knockout studies showed an important role of STAT 3 in islets architecture, although it is dispensable regarding the mature islets function [131]. On the other hand, STAT 5 is only significant for age dependent glucose tolerance [132]. From these studies it is evident that β-cells function is hardly affected by JAK-STAT pathway inhibition.
Summary: Together this shows IFN- stimulated TID pathogenesis can get potentially downregulated by inhibiting the downstream JAK-STAT pathway.
Conclusion
Despite the pathogenesis of T1D has not got clarified, our insight of the natural history has enhanced over years. Especially we can recognize the subjects at risk for developing disease prior to diagnosis, trying to manipulate the preclinical stage with different methods both for primary as well as secondary prevention, with the objective of preserving β-cells mass. Already experimental models have shown that the mucosal immune tolerance, brought about by delivery of autoantigens, can avoid the onset of autoimmune disease. Different immunotherapies used for therapy or prevention of TID have been unmasked in the last decade ,but completed studies so far have not been successful in preservation of β-cells mass from the autoimmune attack .In total various immunotherapies have the objective of targeting pathogenic immunological modes caused by pathogenic autoreactive T cells. By blocking stimulation, inducing exhaustion of the expanded CD8+T cells or blocking the action of proinflammatory cytokines like TNFα,IL -1β. Therapies might also induce Treg cell formation, increase their function and escalate the T reg/Teff ratio. Particular therapies (like Rituximab) can further affect antigen presentation as well as autoantibody synthesis by inducing B lymphocyte depletion or by influence on other immunotypes (particularly omega -3 fatty acids), as well as macrophages, DC’s as well as NK cells [47, 133, 134]. A pilot study (POSEIDON) is going to evaluate the safety and efficacy of a regimen that combines omega -3 fatty acids and Vitamin D in subjects at TID onset (NCT 03406897). Vitamin D is known to have effects on the development of Tregs cells [133]. While omega -3 fatty acids influence various immunotypes that include macrophages, neutrophils, eosinophils, basophils, DC’s, NKs, mast cells as well as B as well as T lymphocytes [134] and moreover they are known to escalate Treg cells differentiation [134]. As per the long term management of the disease, phenomena like ‘’obesity paradox’’ with an inverse correlation between body mass index (BMI) as well as escalated risk of mortality for cardiovascular failure in TID, have been disputed in few epidemiological studies conducted over the last 10yrs [135]. No primary as well as secondary prevention could at current stage of analysis prevent the formation of clinical diseases in at risk subjects. Greater clinical as well as molecular studies are required to get more insight regarding the inflammatory process that takes place in the islets of Langerhans at various stages of TID formation, for improvement of therapeutic methods that have been targeted for avoiding and reversing β-cells mass damage. The best practical strategy is to halt the further evolving of β-cells dysfunction by intervention very close to disease initiation.
More individual as well as combination immunotherapies might potentially be experimented upon in future to abrogate glycemic control as well as preserve β-cells function, hence aiding in >efficient strategy for this disease. Further studies need to depend on improvement in designs in the preclinical as well as clinical settings. Greater refined methods in future need to particularly manipulate islet–specific modes that underlie tolerance, prevent toxic side effects or earlier broad immunosuppressive drugs. Targeted therapies need to be based on >insight of the crosstalk between genetic background, environmental factors, disease triggers as well as immunological mechanisms.
References
-
Patterson C, Guariguata L, Dahlquist G, Soltesz G, Ogle G, et al. (2014) Diabetes in the young-A global view and worldwide estimates of number of children with type 1 diabetes. Diabetes Res Clin Pract 103(2): 161-175.
-
Maahs DM, West NA, Lawrence JM, Davis EJM (2010) Epidemiology of type 1 diabetes. Endocrinol Metab Clin North Am 39(3): 481-497.
-
Kaur KK, Allahbadia GN, Singh M (2017) An Update on Etiopathogenesis and Management of Type 1 Diabetes Mellitus. Open Access Journal of Endocrinology 1(2): 1-23.
-
Kaur KK, Allahbadia GN, Singh M (2019) Restricting Carbohydrates in the Diet-A Possible Method of Meeting the Challenges of Increasing Diabesity in Type1 Diabetes along with Meeting Exercise Performances Requirements-A Review. J Endocrinol 3(1): 1-20.
-
Kaur KK, Allahbadia GN, Singh M (2019) How can we use Empagliflozin as an adjuvant in reducing required need of insulin in type1 diabetes along with lowered HbA1c, weight without fear of DKA-A Minireview. J Clin Case Rep 4(2): 30-38.
-
Barrett JC, Clayton DG, Concannon P, Akolkar B, Cooper JD, et al. (2009) Genome-wide association study and meta-analysis find that over 40loci affect risk of type1 diabetes. Nat Genet 41(6): 703-707.
-
Clark M, Kroger CJ, Tisch RM (2017) Type 1 Diabetes: A chronic-Anti-self-inflammatory response. Front Immunol 8: 1898.
-
Shapiro AM, Ricordi C, Hering BJ, Auchingcloss H, Lindblad R, et al. (2006) International trial of the Edmonton Protocol for islet transplantation. N Engl J Med 355(13): 1318-1330.
-
Perseghin G, Fiorina P, Cobelli FD, Scifo P, Esposito A, et al. (2005) Cross-sectional assessment of the effect of kidney and kidney-pancreas transplantation in resting left ventricular energy metabolism in type 1 diabetes- uremic patients: A phosphorus-31 magnetic resonance spectroscopy study. J Am Coll Cardiol 46(6): 1085-1092.
-
Regnell SE, Lenmark A (2017) Early prediction of autoimmune (type 1) diabetes. Diabetologia 60(8): 1370-1381.
-
Di Lorenzo TP, Peakman M, Roep BO (2007) Translational mini-review series on type 1 diabetes: Systematic analysis of Tcell epitopes in autoimmune diseases. Clin Exp Immunol 148(1): 1-16.
-
Wherrett DK, Chang JL, Delamater AM, DiMeglio LA, Gitelman SE, et al. (2015) Defining pathways for development of disease modifying therapies in children with type 1 diabetes: A consensus report. Diabetes Care 38(10): 1975-1985.
-
Parikka V, Nanto-Salonen K, Saarinen M, Simell T, Ilonen J, et al. (2012) Early seroconversion and rapidly increasing autoantibody concentrations predict pubertal prepubertal manifestation of type 1 diabetes in children at genetic risk. Diabetologia 55(7): 1926-1936.
-
Krischer JP, Lynch KF, Schatz DA, Ilonen J, Lenmark A, et al. (2015) The 6 year incidence of diabetes-associated autoantibodies in genetically at risk children: the TEDDY study. Diabetologia 58(5): 980-987.
-
Bosi E, Boulware DC, Becker DJ, Buckner JH, Geyer S, et al. (2017) Impact of age and antibody type on progression from single to multiple autoantibodies in type 1 diabetes relatives. J Clin Endocrinol Metab 102(8): 2881-2886.
-
American Diabetes Association (2019) Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2019. Diabetes Care 42(1): 13-28.
-
Herold KC, Gitelman SE, Ehlers MR, Gottlieb PA, Greenbaum CJ, et al. (2013) Teplizumab (antiCD3mAb) treatment preserves C-peptide responses in patients with new-onset type 1 diabetes in a randomized controlled trial: Metabolic and immunologic features at baseline identify a subgroup of responders. Diabetes 62(11): 3766-3774.
-
Rigby MR, DiMeglio LA, Rendell MS, Felner EJ, Dostou JP, et al. (2013) Targeting of memory T cells with alefacept in new-onset type 1 diabetes (TIDAL study): 12 month results of a randomized, double blind placebo controlled phase 2 trial. Lancet Diabetes Endocrinol 1(4): 284-294.
-
Mahon JL, Sosenko JM, Rafkin-Mervis L, Krause-Steinrauf H, Lachin JM, et al. (2009) The Trial Net Natural History Study of the Development of type 1 diabetes: Objectives, design and initial results. Pediatr Diabetes 10(2): 97- 104.
-
Zhao Z, Miao D, Michaels A, Steck A, Dong F, et al. (2016) A multiplex assay combining insulin, GAD, IA2 and transglutaminase autoantibodies to facilitate screening for pre type 1 diabetes and celiac disease. J Immunol Methods 430: 28-32.
-
Slama Slim IBH (2013) Cardiovascular risks in type 1 diabetes mellitus. Indian J Endocrinol Metab 17(1): 7-13.
-
Makino S, Kunimoto K, Muraoka Y, Mizushima Y, Katagiri K, et al. (1980) Breeding of a nonobese, diabetic strain of mice. Jikken Dobutsu 29(1): 1-13.
-
Jackson R, Rassi N, Crump T, Haynes B, Eisenbarth GS (1981) The BB diabetic rat: Profound T-cell lymphocytopenia. Diabetes 30(10): 887-889.
-
Matsumoto M, Yagi H, Kunimoto K, Kawaguchi J, Makino S, et al. (1993) Transfer of autoimmune diabetes from, diabetic NOD mice to NOD athymic nude mice: The roles of Tcell subsets in the pathogenesis. Cell Immunol 148(1): 189-197.
-
Pontesilli O, Carotenuto P, Gazda LS, Pratt PE, Prowse SJ (1987) Circulating lymphocyte populations and autoantibodies in non-obese diabetic(NOD) mice: A longitudinal study. Clin Exp Immunol 70(1): 84-93.
-
Shoda LK, Young DL, Ramanujan S, Whiting CC, Atkinson MA, et al. (2005) A comprehensive review of interventions in the NOD mouse and implications for translation. Immunity 23(2): 115-126.
-
Pow Sang L, Majji S, Casares S, Brumeanu TD (2014) Long term silencing of autoimmune diabetes and improved life expectancy by a soluble PHLA-DR4 chimera in a newly humanized NOD/DR4/B7 mouse. Human Vaccines Immunother 10(3): 693-699.
-
Peng J, Naraasimhan S, Marchesi JR, Benson A, Wong FS, et al. (2014) Long term effects of gut microbiota transfer on diabetes development. J Autoimmun 53: 85-94.
-
Nagy N, Kaber G, Johnson PY, Gebe JA, Preisinger A, et al. (2015) Inhibition of hyaluronan synthesis restores immune tolerance during autoimmune insulitis. J Clin Invest 125(10): 3928-3940.
-
Makhlouf L, Grey ST, Dong V, Csizmadia E, Arvelo MB, et al. (2004) Depleting anti-CD4 monoclonal antibody cures new–onset diabetes, prevents recurrent autoimmune diabetes and delays graft rejection in non-obese diabetic mice with advanced insulitis. Transplantation 77(7): 990-997.
-
Cameron MJ, Arreaza GA, Waldhauser L, Gaudie J, Delvotich TL (2000) Immunotherapy of spontaneous type 1 diabetes in non-obese diabetic mice by systemic interleukin 4 treatment employing adenovirus vector – mediated gene transfer. Gene Ther 7(21): 1840-1846.
-
Feil-Hariri M, Falkner DH, Gambotto A, Papworth GD, Watkins SC, et al. (2003) Dendritic cells transduced to express interleukin 4 prevents diabetes in non-obese diabetic mice with advanced insulitis. Human Gene Ther 14(1): 13-23.
-
Clemente-Casares X, Tsai S, Huang C, Santamaria P (2012) Antigen-specific therapeutic approaches in type 1 diabetes. Cold Spring Harb Perspect Med 2(2): a007773.
-
Bone RN, Evans-Molina C (2017) Combination immunotherapy for type 1 diabetes. Curr Diabetes Rep 17(7): 50.
-
Vital EM, Emery P (2006) Abatacept in the treatment of rheumatoid arthritis. Ther Clin Risk Manag 2(4): 365- 375.
-
Orban T, Bundy B, Becker DJ, DiMeglio LA, Gitelman SE, et al. (2011) Costimulation modulation with Abatacept in patients with recent-onset type 1 diabetes: a randomized, double blind placebo controlled trial. Lancet 378(9789): 412-419.
-
Herold KC, HagopianWA, Auger JA, Poumian-Ruiz E, Taylor L, et al. (2002) Anti-CD3 monoclonal antibody in new-onset type 1 diabetes. N Engl J Med 346(22): 1692- 1698.
-
Haller MJ, Gitelman SE, Gottlieb PA, Michaels AW, Rosenthal SM, et al. (2015) Antithymocyte globulin/G- CSF treatment preserves beta cell function in patients with established type 1 diabetes. J Clin Investig 125(1): 448-455.
-
Haller MJ, Schatz DA, Skyler JS, Krischer JP, Bundy B, et al. (2018) Low dose Antithymocyte globulin (ATG) Preserves beta cell function and improves Hb A1 C in new-onset type 1 diabetes. Diabetes Care 41(9): 1917- 1925.
-
LopesM, Kutlu B, Miani M, Bang-Berthelson CH, Stortling J, et al. (2014) Temporal profiling of cytokine-induced genes in pancreatic beta cells by meta-analysis and network inference. Genomics 103(4): 264-275.
-
Nepom GT, Ehlers MR, Mandrup-Poulsen T (2013) Anti- cytokine therapies in T1D: Concepts and strategies. Clin Immunol 149(3): 279-285.
-
Cabrera SM, Wang X, Chen YG, Jia S, Kaldunski ML, et al. (2016) Type 1 diabetes Trial Net Canakinumab Study Group, Mandrup-Poulsen T, Group AS, Hessner MJ. Interleukin -1 antagonism modulates the inflammatory state associated with Type 1 diabetes during clinical trials conducted at disease onset. Eur J Immunol 46(4): 1030-1046.
-
Den Broeder AA, De Jong E, Franssen MJ, Jeurissen ME, Flendrie M, et al. (2006) Observational study on efficacy, safety, and drug survival of anakinra in rheumatoid arthritis patients in clinical practice. Ann Rheum Dis 65(6): 760-762.
-
Mandrup-Poulsen T, Pickersgill L, Donath MY (2010) Blockade of Interleukin -1 in Type 1 diabetes. Nat Rev Endocrinol 6: 158-166.
-
Sumpter KM, Adhikari S, Grishman EK, White PC (2011) Preliminary studies related to anti-Interleukin-1 beta therapy in children with newly diagnosed type 1 diabetes. Pediatr Diabetes 12(7): 656-667.
-
Mastrandrea L, Yu J, Behrens T, Buchlis J, Albini C, et al. (2009) Etanercept treatment in children with new-onset type 1 diabetes: Pilot randomized, placebo controlled double blind study. Diabetes Care 32(7): 1244-1249.
-
Crino A, Schiaffini R, Manfrini S, Mesturino C, Visalli N, et al. (2004) A randomized trial of nicotinamide and vitamin E in children with recent-onset type 1 diabetes (IMDIAB IX). Eur J Endocrinol 150(5): 719-724.
-
Hundhausen C, RothA, Whalen E, Chen J, Schneider A, et al. (2016) Enhanced T cell responses to IL-6 in type 1 diabetes are associated with early clinical disease and increased IL-6 receptor expression. Sci Transl Med 8(356): 356ra119.
-
Tanaka T, Narazaki M, Kishimoto T (2018) Interleukin- 6(IL-6) Immunotherapy. Cold Spring Harb Perspect Biol 10.
-
Bluestone JA, Buckner JH, Fitch M, Gitelman SE, Gupta S, et al. (2015) Type 1 diabetes Immunotherapy using polyclonal regulatory T cells. Sci Transl Med 7(315): 315ra189.
-
Hartemann A, Bensimon G, Payan CA, Jacqueminet S, Bourron O, et al. (2013) Low dose Interleukin -2 in patients with type 1 diabetes: A phase ½ randomized, double blind, placebo controlled-trial. Lancet Diabetes Endocrinol 1(4): 295-305.
-
Bogdani M, Johnson PY, Potter-Perigo S, Nagy N, Day AJ, et al. (2014) Hyaluronan and Hyaluronan binding protein accumulate in both human type 1 diabetic islets and lymphoid tissues and associate with inflammatory cells and insulitis. Diabetes 63(8): 2727-2743.
-
Kuhn C, Weiner HL (2016) Therapeutic anti CD3 monoclonal antibodies: From bench to bedside. Immunotherapy 8(8): 889-906.
-
Marino E, Silviera PA, Stolp J, Grey ST (2011) B-cell directed therapies in type 1 diabetes. Trends Immunol 32(6): 287-294.
-
Pescowitz MD, Greenbaum CJ, Krause-Steinrauf H, Becker DJ, Gitelman SE, et al. (2009) Rituximab, B-lymphocyte depletion and preservation of beta cells function. N Engl J Med 361: 2143-2152.
-
Townsend MJ, Monroe JG, Chan AC (2010) B-cell targeted therapies in human autoimmune diseases: an updated perspective. Immunol Rev 237(1): 264-283.
-
Coppieters K, Von Herrath M (2017) Antigen specific peptide Immunotherapy for Type 1 diabetes: proof of safety, hope for efficacy. Cell Metab 26(4): 595-597.
-
Polychronakos C, Li Q (2011) Understanding type 1 diabetes through genetics: Advances and prospects. Nat Rev Genet 12(11): 781-792.
-
Thrower SL, James L, Hall W, Green KM, Arif S, et al. (2009) Proinsulin peptide Immunotherapy in Type 1 diabetes: reports of a first-in-man Phase 1 safety study. Clin Exp Immunol 155(2): 156-165.
-
Alhadj Ali M, Liu YF, Arif S, Tatovic D, Shariff H, et al. (2017) Metabolic and immune effects of Immunotherapy with Proinsulin peptide in new-onset type 1 diabetes. Sci Transl Med 9(402).
-
Tian J, Clare SM, Herschenfeld A, Middleton B, Newman D, et al. (1996) Modulating autoimmune responses to GAD inhibits disease progression and prolongs islet graft survival in diabetes-prone mice. Nat Med 2(12): 1348- 353.
-
Iwasaki A, Medzhitov R (2010) Regulation of adaptive immunity by the innate immune system. Science 327(5963): 291-295.
-
Wherrett DK, Bundy B, Becker DJ, DiMeglio LA, Gitelman SE, et al. (2011) Antigen-based therapy with glutamic acid decarboxylase (GAD) vaccine in patients with recent-onset type1 diabetes: A randomized, double blind trial. Lancet 378(9788): 319-327.
-
Yu H, Paiva R, Flavell RA (2018) Harnessing the power of regulatory T-cells to control autoimmune diabetes: overview and perspective. Immunology 153(2): 161- 170.
-
Guilliams M, Ginhoux F, Jazubzick C, Naik SH, Onai N, et al. (2014) Dendritic cells, monocytes and macrophages: A united nomenclature based on ontogeny. Nat Rev Immunol 14(8): 571-578.
-
Henry RA, Kendall PL, Thomas JW (2012) Autoantigen– specific B-cell depletion overcomes failed immune tolerance in Type 1 diabetes. Diabetes 61(8): 2037-2044.
-
Balamurugan AN, Nazirruddin B, Lockridge A, Tiwari M, Loganathan G, et al. (2014) Islet product characteristics and factors related to successful human islet transplantation from the Collaborative islet transplant Registry (CITR) 1999-2010. Am J Transplant 14(11): 2595-2606.
-
Faradji RN, Tharavanij T, Messinger S, Froud T, Pileggi T, et al. (2008) Long term insulin independence and improvement in insulin secretion after supplemental islets infusion under exenatide and etanercept. Transplantation 86(12): 1658-1665.
-
Gangemi A, Salehi P, Hatimoglu B, Martelotto J, Barbaro B, et al. (2008) Islet transplantation for brittle Type 1 diabetes: The UIC protocol. Am J Transplant 8(6): 1250- 1261.
-
Suarez PWL, Cembrowski GS, Rabinovitch A (2009) Combination therapy with a dipeptyl-peptidase 4 inhibitor and a proton pump inhibitor restores normoglycaemia in diabetic NOD mice. Diabetologia 52(8): 1680-1682.
-
Suarez PWL, Rabinovitch A (2011) Combination therapy with a dipeptyl peptidase-4 inhibitor and a proton pump inhibitor induces β Cells neogenesis from adult human pancreatic duct cells implanted in immunodeficient mice. Cell Transplant 20(9): 1343-1349.
-
Suarez PWL, Power RF, Yan Y, Wasserfall C, Atkinson M, et al. (2008) Combination therapy with Glucagon like peptide-1 and Gastrin restores normoglycaemia in diabetic NOD mice. Diabetes 57(12): 3281-3288.
-
Griffin KJ, Thompson PA, Gottschalk M, Kyollo JH, Rabinovitch A (2014) Combination therapy with sitagliptin and lansoprazole in patients with recent onset type 1 diabetes (REPAIR-TID): 12months results of a multicentre randomized, placebo controlled, phase 2 trial. Lancet Diabetes Endocrinol 2(9): 710-718.
-
Giannoukakis N, Philips B, Finegold D, Harnaha J, Trucco M (2011) Phase 1 (safety)study of autologous tolerogenic Dendritic cells in type 1 diabetic patients. Diabetes Care 34(9): 2026-2032.
-
Bell GM, Anderson AE, Diboll J, Reece R, Eitherington O, et al. (2017) Autologous tolerogenic Dendritic cells for rheumatoid and inflammatory arthritis. Ann Rheum Dis 76(1): 227-234.
-
Vigourox S, Yvon E, Biagi E, Brenner MK (2004) Antigen induced regulatory T cells. Blood 104(1): 26-33.
-
Van de Pavert SA, Mebius RE (2010) New insights into the development of lymphoid tissues. Nat Rev Immunol 10(9): 664-674.
-
Di Caro V, Phillips B, Engman C, Harnaha J, Trucco M, et al. (2013) Retinoic acid producing, ex-vivo generated human tolerogenic Dendritic cells induce the proliferation of immunosuppressive B lymphocytes. Clin Exp Immunol 174(2): 302-317.
-
Voltarelli JC, Couri CE, Stracleri AB, Oliviera MC, Moraes DA, et al. (2007) Autologous nonmyeloblastic Haematopoietic Stem Cells transplantation in newly diagnosed type 1 diabetes mellitus. JAMA 297(14): 1568-1576.
-
Snarski E, MJilczarezyk A, Halaburda K, Torosian T, Paluszewska M, et al. (2016) Immunoablation and Autologous Haematopoietic Stem Cells transplantation in the treatment of new-onset diagnosed type 1 diabetes mellitus: long term observations. Bone Marrow Transplant 51(3): 398-402.
-
DAddio F, Valderrama VA, Ben NM, Franek E, Zhu D, et al. (2014) Autologous nonmyeloblastic Haematopoietic Stem Cells transplantation in new-onset type 1 diabetes: A multicenter analysis. Diabetes 63(9): 3041-3046.
-
Murphy MB, Moncivais K, Caplan AI (2013) Mesenchymal Stem Cells: Environmentally responsive therapies for regenerative medicine. Exp Mol Med 45: 54.
-
Bianco P, Cao X, Frenette PS, Mao JJ, Robey PG, et al. (2013) The meaning, the sense and the significance: Translating the science of Mesenchymal Stem Cells into medicine. Nat Med 19(1): 35-42.
-
Wang Y, Chen X, Cao W, Shi Y (2014) Plasticity of Mesenchymal Stem Cells in immunomodulation: pathological and therapeutical implications. Nat Immunol 15(11): 1009-1016
-
Moreira A, Kahlenburg S, Hornsby P (2017) Therapeutic potential of Mesenchymal Stem Cells for diabetes. J Mol Endocrinol 59(3): 109-120.
-
Carlsson PO, Schwarcz E, Korsgren O, Le Blanc K (2015) Preserved beta cell function in type1 diabetes by Mesenchymal Stem Cells. Diabetes 64(2): 587-592.
-
Cai J, Wu Z, Wu X, Liao L, Chen J, et al. (2016) Umbilical cord Mesenchymal Stromal Cells with Autologous Bone Marrow Cells transplantation in established type 1 diabetes: A pilot randomized, controlled open-label Clinical Study to assess safety and impact on insulin secretion. Diabetes Care 39(1): 149-157.
-
Maude S, Barrett D (2016) Current status of chimeric antigen receptor therapy for hematological malignancies. Br J Haematol 172(1): 11-22.
-
Grupp SA, Kalos M, Barrett DM, Aplenc R, Porter D, et al. (2013) Chimeric antigen receptor-modified T cells for acute lymphoid laekaemia. N Engl J Med 368: 1509-1518.
-
Yoon J, Schmidt A, Zhang AH, Konigs C, Kim YC, et al. (2017) FVIII-specific human Chimeric antigen receptor- T-regulatory cells suppress T and B cells responses to FVIII. Blood 129(2): 238-245.
-
Macdonald KG, Hoeppli RE, Huang Q, Gilles J, Luciani DS, et al. (2016) Alloantigen-specific regulatory T cells generated with a chimeric antigen receptor. J Clin Invest 126: 1413-1424.
-
Noyan F, Zimmermann K, Hardtke-Wolenski M, Knoefel A, Schulde E, et al. (2017) Prevention of allograft rejection by use of regulatory T cells with an MHC- Specific Chimeric antigen receptor. Am J Transplant 17(4): 917-930.
-
Fransson M, Piras E, Burman J, Nilsson B, Essand M, et al. (2012) CAR/ Foxp3- engineered T regulatory cells target the CNS and suppress EAE upon intranasal delivery. J Neuroinflamm 9: 112.
-
Skuljec J, Chmielewski M, Happle C, Habener A, Busse M, et al. (2017) Chimeric antigen receptor –Redirected regulatory T cells suppress experimental allergic airway inflammation,a model of asthma. Front Immunol 8: 1125.
-
Kim YC, Zhang AH, Su Y, Rieder SA, Rossi RJ, et al. (2015) Engineered antigen-specific human -regulatory T cells: immunosuppression of FVIII-specific T and B-cells responses. Blood 125: 1107-1115.
-
Hull CM, Nickolay LE, Estorninho M, Richardson MW, Riley JL, et al. (2017) Generation of human islet-specific regulatory T cells by TCR gene transfer. J Autoimmun 79: 63-73.
-
Tenspolde M, Zimmermann K, Weber LC, Hapke M, Lieber M, et al. (2019) Regulatory T cells engineered with a novel insulin specific Chimeric antigen receptor as a candidate immunotherapy for type 1 diabetes. J Autoimmun 103: 102289.
-
Balhuizen A, Massa S, Mathijs I, Turatsinze JV, DeVos J, et al. (2017) A nanobody –based tracer targeting DPP6 for noninvasive imaging of human pancreatic endocrine cells. Sci Rep 7(1): 15130.
-
Burtea C, Laurent S, Crombez D, Delcambre S, Sermeus C, et al. (2015) Development of a peptide –functionalized imaging nanoprobe for the targeting of (FXYD2) gamma as highly specific biomarker of pancreatic beta cells. Contrast Media Mol Imaging 10(5): 398-412.
-
Saunders DC, Brissova M, Phillips N, Shrestha S, Walker JT, et al. (2019) Ectonucleoside Triphosphate Diphosphohydrolase -3 Antibody Targets adult human pancreatic beta cells for in vitro and in vivo analysis. Cell Metab 29(3): 745-754.
-
Cabello-Olmo M, Arana M, Radichev I, Smith P, Huarte E, et al. (2019) New insights into immunotherapy strategies for treating Autoimmune Diabetes. Int J Mol Sci 20(19): 4789.
-
Kulvinder Kochar Kaur, Allahbadia GN, Singh M (2016) An Update on a etiopathogenesis and Management of Obesity. Obes Control Ther 3(1): 1-17.
-
Kulvinder Kochar Kaur, Allahbadia GN, Singh M (2017) Hypothalamic inflammation and glioses as aetiopathogenetic factor in high fat diet induced obesity and various therapeutic options to resolve it. Obes Res Open J 4(2): 44-60.
-
Kulvinder Kochar Kaur, Allahbadia GN, Singh M (2018) Current advances in pathogenesis in obesity: Impact of Hypothaalamic glioses. J Obes Weight Loss 3(8): 1-11.
-
Kulvinder Kochar Kaur, Allahbadia GN, Singh M (2019) Have Probiotics and Synbiotics passed the test of time to be implemented in management of obesity and related metabolic disorders-a comprehensive review. Adv Obes Weight Manag Control 9(1): 21-28.
-
Kulvinder Kochar Kaur, Allahbadia GN, Singh M (2018) Weight loss Associated with high protein Intake in Obesity: Interactions of Gut Microbiota in Protein Sources influencing this positive effect. Acta Scientific Nutritional Health 2(7): 80-89.
-
Kulvinder Kochar Kaur, Allahbadia GN, Singh M (2020) Will Probiotics Provide the Answer for Therapy of Non-alcoholic Fatty Liver Disease (NAFLD)? – A Systematic Review. Biochem Physiol 9(1): 257.
-
Depommier C, Everard A, Druart C, Plovier H, Van Hul M, et al. (2019) Supplementation with Akkermansia Muciniphila in overweight and obese human volunteers: A proof of concept exploratory study. Nat Med 25(7): 1096-1103.
-
Giongo A, Gano KA, Crabb DB, Mukherjee N, Novelo LL, et al. (2011) Toward defining the autoimmune microbiome for type 1 diabetes. ISME J 5(1): 82-91.
-
Huang Y, Li SC, Hu J, Ruan HB, Guo HM, et al. (2018) Gut microbiota profiling in Han Chinese with type 1 diabetes. Diabetes Res Clin Pract 141: 256-263.
-
Paun A, Yau C, Danska JS (2017) The influence of microbiome on type 1 diabetes. J Immunol 198(2): 590- 595.
-
Kriegel MA, Sefik E, Hill JA, Wu HJ, Benoist C, et al. (2011) Naturally transmitted segmented filamentous bacteria segregate with diabetes protection in non- obese diabetic mice. Proc Natl Acad Sci USA 108(28): 11548-1563.
-
Villarno AV, Kanno Y, O’Shea JJ (2017) Mechanisms and consequences of JAK-STAT signaling in the immune system. Nat Immunol 18(4): 374-384.
-
Rawlings JS, Rosler KM, Harrison D (2004) The JAK- STAT signaling pathway. J Cell Sci 117: 1281-283.
-
Howell MD, Fitzsimons C, Smith PA (2018) JAK-STAT inhibitors and other small molecule cytokine antagonists for the treatment of allergic disease. Ann Allergy Asthma Immunol 120(4): 367-375.
-
Ferreira RC, Guo H, Coulson RM, Smyth DJ, Pekalski ML, et al. (2014) A type 1 interferon transcriptional signature precedes autoimmunity in children genetically at risk for type 1 diabetes. Diabetes 63(7): 2538-2550.
-
Huang X, Yuang J, Goddard A, Foulis A, James RF, et al. (1995) Interferon expression in the pancreases of patients with type 1 diabetes. Diabetes 44(6): 658-664.
-
Foulis AK, Farquharson MA, Meager A (1987) Immunoreactive alpha interferon in insulin secreting β-cells in type 1 diabetes mellitus. Lancet 2(8573): 1423-1427.
-
Richardson SJ, Rofdriques-Calvo T, Gerling IC, Mathews CE, Kaddis JS, et al. (2016) Islet cell hyperexpression of HLA Class 1 antigens: A defining feature in type 1 diabetes. Diabetologia 59(11): 2448- 2458.
-
Marroqui L, Dos Santos RS, Op de Beeck A, Coomans de Brachene A, Marselli L, et al. (2017) Interferon alpha mediates human β-cells HLA Class 1 over expression, endoplasmic reticulum stress and apoptosis, three hallmarks of early human type 1 diabetes. Diabetologia 60(4): 656-667.
-
Tao JH, Zou YF, Feng XL, Li J, Wang F, et al. (2011) Meta-analysis of TYK2 gene polymorphisms association with susceptibility to autoimmune and inflammatory diseases. Mol Biol Rep 38(7): 4663-4672.
-
Roep BO, Kleijwegi FS, Van Halteren AG, Bonato V, Boggi U, et al. (2010) Islet inflammation and CXCL10 in recent onset type 1 diabetes. Clin Exp Immunol 159(3): 338-343.
-
Morimoto J, Yoneyama H, Shimada A, Shigihara T, Yamada S, et al. (2004) CXC chemokine ligand 10 neutralization suppresses the occurrence of diabetes in non-obese diabetic mice through advanced β-cells proliferation without affecting insulitis. J Immunol 173(11): 7017-7024.
-
Savinov AV, Wong FS, Chernovsky AV (2001) IFN-γ Affects Homing of Diabetogenic T Cells. J Immunol 167(11): 6637-6643.
-
Barrosa-Sousa R, Ott PA, Hodi FS, Kaiser UB, Tolaney SM, et al. (2018) Endocrine dysfunction induced by immune check point inhibitors: practical recommendations for diagnosis and clinical management. Cancer 124(6): 1111-1121.
-
Cuckier P, Santini FC, Scaranti M, Hoff AO (2017) Endocrine side effects of cancer immunotherapy. Endocr Relat Cancer 24(12): 331-347.
-
Stamouli AM, Quandt Z, Perdigoto AL, Clark PL, Kluger H, et al. (2018) Collateral damage: Insulin dependent diabetes Induced with check point inhibitors. Diabetes 67(8): 1471-1480.
-
Ansari MJ, Salama AD, Chitnis D, Smith RN, Yagita H, et al. (2003) The programmed death(PD1)pathway regulates autoimmune Diabetes in non-obese diabetic (NOD) mice. J Exp Med 198(1): 63-69.
-
Coomans de Brachene A, Dos Santos RS, Marroqui L, Colli ML, Marselli L, et al. (2018) IFN-alpha induces a preferential long lasting expression of MHC class 1 in human pancreatic beta cells. Diabetologia 61(3): 636- 640.
-
Trivedi PM, Graham KL, Scott NA, Jenkins MR, Majaw S, et al. (2017) Repurposed JAK1/ JAK2 inhibitor reverses established autoimmune insulitis in NOD mice. Diabetes 66(6): 1650-1660.
-
Kostromina E, Wang X, Han W (2010) Altered Islet morphology but normal islet secretory function in vitro in a mouse model with microvascular alterations in the pancreas. PLoS One 8(7): 71277.
-
Lee JY, Gavrilova O, Davani B, Na R, Robinson GW, et al. (2007) The transcription factors STAT5a/b are not required for islet development but modulate pancreatic β-cells physiology upon aging. Biochim Biophys Acta 1773(9): 1455-1461.
-
Issazadeh-Navikas S, Teimer R, Boeckermann R (2012) Influences of dietary components on regulatory T cells. Mol Med 18(1): 95-110.
-
Gutierrez S, Svahn SL, Johansson ME (2019) Effects of omega -3 fatty acids on immune cells. Int J Mol Sci 20(20): 5028.
-
Edqvist J, Rawshani A, Adiels M, Bjorck L, Lind M, et al. (2019) BMI, Mortality and Cardiovascular outcomes in type 1 diabetes: Findings against an Obesity Paradox. Diabetes Care 42(7): 1297-1304.
- Shaping Healthy Futures: Pediatric Endocrine Breakthroughs of 2025
- Precision Medicine in Obesity: Customizing Treatment for 2025
- The Thyroid Revolution: How 2025 is Redefining Hormone Health
- Editorial- Targeting Immunometabolism for Generating Innovative Therapies for Cancer
- Current Knowledge of Chickenpox
- Correlation of Preinjection Values of Gonadotropins and Estradiol Level with Clinical and Radiologic Evidence of Sufficient Pubertal Suppression in Girls with Central Precocious Puberty