Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
Lipases are vital biocatalysts with extensive industrial and environmental applications. This study aimed to isolate and screen lipase-producing bacteria and fungi from oil-contaminated sites in Makurdi metropolis, Nigeria. A total of 47 microbial isolates, comprising 28 bacteria and 19 fungi, were recovered from seven selected sites. The bacterial isolates included Bacillus spp., Pseudomonas aeruginosa, Staphylococcus aureus, Enterobacter spp., Flavobacterium spp., Streptococcus spp., and Bacillus subtilis. The fungal isolates included Aspergillus spp., Penicillium spp., Rhizopus spp., Mucor spp., and Neurospora spp. Lipolytic activity was assessed using Tween 80 agar and Phenol Red Olive Oil Agar. All isolates exhibited varying degrees of lipase production (p˂0.05), as evidenced by clear zones around colonies. Among the bacterial isolates, Bacillus spp. demonstrated the highest lipolytic activity on Tween 80 agar, with clearance zones ranging from 44 mm to 62 mm. Pseudomonas aeruginosa and Staphylococcus aureus also exhibited significant activity, with peak clearance zones of 58 mm and 55 mm, respectively, at 120 hours. On Phenol Red Olive Oil Agar, Flavobacterium spp. showed the lowest lipase activity (5-9 mm), while Pseudomonas aeruginosa and Enterobacter spp. recorded moderate activity (4-8 mm). Among the fungi, Neurospora spp., Aspergillus flavus, and Aspergillus niger exhibited the highest activity, with clearance zones up to 83 mm. These findings highlight the potential of indigenous microbial strains for biotechnological applications, including bioremediation and industrial enzyme production. Further optimization of fermentation conditions and enzyme purification is necessary to enhance lipase yields for commercial and industrial use.
Ado BV¹*, Ajekwe MS¹ and Omeonu FC²
¹Department of Microbiology, College of Biological Sciences, Joseph Sarwuan Tarka University, Nigeria ²Department of Microbiology, Chrisland University, Nigeria
Introduction
Lipases (triacylglycerol acylhydrolases, EC 3.1.1.3) are versatile enzymes that catalyze the hydrolysis of triglycerides into free fatty acids and glycerol. These enzymes are crucial in numerous industrial applications, including food processing, pharmaceuticals, detergent formulation, and bioremediation [1]. Microbial lipases, particularly those produced by bacteria and fungi, have gained prominence due to their high catalytic efficiency, substrate specificity, stability under diverse conditions, and ease of production through fermentation processes [1, 2]. Identifying and characterizing microbial lipase producers is essential for optimizing enzyme applications in biotechnology and environmental management. Oil-contaminated environments provide a natural reservoir for lipolytic microorganisms, selecting species capable of degrading lipid-rich waste. Studies have highlighted Bacillus spp_., _Pseudomonas spp_., _Enterobacter spp_., and _Staphylococcus spp., among bacteria, and Aspergillus spp_., _Penicillium spp_., _Rhizopus spp_., _Neurospora spp_., and _Mucor spp_._ among fungi, as prominent lipase producers in hydrocarbon-polluted ecosystems [1, 3, 4]. However, variations in lipolytic activity exist across species, strains, and environmental conditions. Screening techniques such as Tween 80 agar and Phenol Red Olive Oil Agar are widely used to differentiate true lipase producers from esterases and to evaluate the enzymatic potential of microbial isolates [5].
Despite extensive research on microbial lipases, comparative assessments of bacterial and fungal lipase activity under different screening conditions remain limited. The selection of an appropriate screening medium is critical, as the effectiveness of lipase detection varies among microbial groups. Tween 80 agar is often more sensitive for bacterial lipase screening, whereas Phenol Red Olive Oil Agar enhances the detection of fungal lipases due to its pH-sensitive colourimetric response [4]. The differences in lipase production among microbial species have significant implications for their industrial and environmental applications. Fungal lipases are particularly valuable due to their higher thermal stability, extracellular production, and adaptability to large-scale fermentation. Aspergillus niger, Rhizopus spp_., and _Neurospora spp_._ are among the most efficient fungal lipase producers, with demonstrated applications in enzyme biotechnology. Unlike bacterial lipases, which typically function optimally under neutral to alkaline conditions, fungal lipases are often acid-labile and perform efficiently in acidic environments [4]. This property enhances their suitability for various industrial processes, including biodiesel production, waste degradation, and pharmaceutical synthesis [1, 4].
This study aimed to isolate and screen lipase-producing bacteria and fungi from oil-contaminated sites within the Makurdi metropolis, Nigeria. The research employed Tween 80 agar and Phenol Red Olive Oil Agar to evaluate the lipolytic potential of microbial isolates and assess their comparative enzyme activity. By identifying and characterizing lipase- producing microorganisms, the study provides valuable insights into their enzymatic potential and highlights their applications in biotechnology and environmental remediation.
Materials and Methods
Sample Collection
Oil-contaminated soil and fish wastewater samples were aseptically collected from multiple sites within Makurdi metropolis, Benue State, Nigeria. The soil samples were obtained from Joseph Sarwuan Tarka University Oil Palm Mill (JSPM), New GRA Oil Palm Mill (NGPM), North Bank Automechanic Site (NBAS), and Apir Automechanic Site (AAS). Fish wastewater samples were collected from New GRA Fish Pond (NGFP), Joseph Sarwuan Tarka University Fish Pond (JSFP), Wurukum Fish Pond (WFP), and Wurukum Market Fish Processing Water (WMFP). Soil samples were collected at a depth of 2-5 cm using a sterile soil auger, placed into sterile plastic bags, and transported to the Microbiology Laboratory at Joseph Sarwuan Tarka University, Makurdi, for analysis. Samples were either processed immediately or stored at 4 °C until further analysis.
Isolation of Lipolytic Microorganisms
For microbial isolation, 10 g of each soil sample was suspended in 90 mL of sterile physiological saline (1:9 dilution). The suspensions were vortexed and agitated at 120 rpm for 30 minutes to dislodge microorganisms from soil particles. The samples were then allowed to settle, and ten-fold serial dilutions were prepared. Aliquots (100 µL) of the appropriate dilutions (10⁻² to 10⁻⁴) were inoculated onto Nutrient Agar (NA) for bacterial isolation and Potato Dextrose Agar (PDA) for fungal isolation using the spread plate technique. The NA plates were incubated at 37 °C for 48 hours, while PDA plates were incubated at 28 °C for 5-7 days. Pure colonies were subcultured and maintained on NA and PDA slants for further characterization.
Screening for Lipase Activity
Lipolytic Screening on Tween-80 Agar
The preliminary screening for lipase activity was performed on Tween-80 agar. The medium consisted of 1% (v/v) Tween-80, 1% (w/v) peptone, 0.5% (w/v) NaCl, 0.01% (w/v) CaCl₂.2H₂O, and 2% (w/v) agar. The pH was adjusted to 6.0 using 1 M NaOH, and the medium was autoclaved at 121 °C for 15 minutes. After solidification, bacterial and fungal isolates were spot-inoculated onto the plates and incubated at 37 °C for 48 hours (bacteria) and 28 °C for 5-7 days (fungi). The appearance of a clear halo surrounding the colonies indicated positive lipase activity Abubakar A, et al. [6].
Lipolytic Screening on Phenol Red Olive Oil Agar
To further evaluate lipase production, isolates were screened on Phenol Red Olive Oil Agar. The medium contained 0.02% (w/v) phenol red, 1.5% (v/v) olive oil, 0.15% (w/v) CaCl₂, and 3% (w/v) agar in 150 mL of distilled water. The pH was adjusted to 7.3-7.4 using 1 M NaOH before autoclaving at 121 °C for 15 minutes. After cooling, 10 mL of olive oil and a 1 mg/mL phenol red solution were added. Plates were inoculated and incubated at 37 °C for bacteria and 28 °C for fungi. Lipolytic activity was identified by a color change from red to yellow Lee LP, et al. [7].
Identification of Lipolytic Microorganisms
Bacterial Identification
Bacterial isolates were characterized based on colony morphology, including shape, size, elevation, margin, pigmentation, and texture. Microscopic examination was conducted following Gram staining and spore staining. Biochemical tests performed included catalase, coagulase, indole production, hydrogen sulfide (H₂S) production, methyl red-Voges-Proskauer (MR-VP) tests, and carbohydrate fermentation tests using glucose, fructose, maltose, lactose, and sucrose. Identification was confirmed using Bergey’s Manual of Determinative Bacteriology (9th ed) [8].
Fungal Identification
Fungal isolates were identified based on macroscopic and microscopic characteristics. Colony morphology, including color, texture, and growth pattern on PDA, was documented. Microscopic examination was performed using lactophenol cotton blue staining to observe the arrangement and morphology of hyphae, conidiophores, phialides, and spores. Species identification was aided by standard mycological keys such as those provided by the British Mycological Society [9].
Statistical analysis
Results obtained from the study were subjected to analysis of variance using one-way ANOVA, and differences between means of test samples were separated by the Duncan Multiple Range Test [10].
Results
Forty-seven (47) microbial isolates, comprising twenty- eight (28) bacterial and nineteen (19) fungal species, were recovered from seven oil-rich sites in Makurdi metropolis. The bacterial isolates included Bacillus spp., Enterobacter spp., Pseudomonas aeruginosa, Staphylococcus aureus, Flavobacterium spp., Bacillus subtilis, Streptococcus spp., and Pseudomonas spp. The fungal isolates comprised Penicillium spp., Aspergillus terreus, Saccharomyces spp., Aspergillus fumigatus, Mucor spp., Aspergillus niger, Penicillium chrysogenum, Penicillium spp., Rhizopus spp., and Neurospora spp. The percentage frequency of occurrence showed that Pseudomonas spp. (12.23%), Bacillus spp. (10.21%), and Staphylococcus aureus (8.17%) were the most frequently occurring bacterial isolates, while Aspergillus fumigatus recorded 6.13% among fungal isolates (Table 1). The distribution of isolates varied across sample sites, with Pseudomonas spp. and Bacillus spp. detected in four locations each, while Staphylococcus aureus appeared in three sites (Table 2). Among fungi, Penicillium spp. and Aspergillus fumigatus were recovered from four and three locations, respectively (Table 3). The morphological and biochemical characteristics of bacterial isolates are detailed in Table 4, while Table 5 outlines the cultural and morphological characteristics of the fungal isolates.
| S/No. | Isolate | Number (%) |
|---|---|---|
| 1 | Bacillus spp. | 5(10.21) |
| 2 | Enterobacter spp | 2(4.08) |
| 3 | Pseudomonas aeruginosa | 3(6.13) |
| 4 | Staphylococcus aureus | 4(8.17) |
| 5 | Pesudomonas spp. | 6(12.23) |
| 6 | Flavobacterium spp. | 2(4.08) |
| 7 | Streptococcus spp. | 2(4.08) |
| 8 | Bacillus subtilis | 3(6.13) |
| 9 | Staphylococcus spp. | 1(2.04) |
| 10 | Penicillium spp. | 2(4.08) |
| 11 | Aspergillus terreus | 2(4. 08) |
| 12 | Candida spp. | 2(4. 08) |
| 13 | Aspergillus fumigus | 3(6.13) |
| 14 | Mucor spp. | 2(4.08) |
| 15 | Aspergillus niger | 2(4.08) |
| 16 | Penicillium chrysogenum | 2(4.08) |
| 17 | Rhizopus spp. | 2(4.08) |
| 18 | Neurospora spp. | 2(4.08) |
| Total | 47(100) |
Table 1: Frequency of occurrence of bacteria and fungi isolated from oil-contaminated soils.
| S/No. | Code | Isolate |
|---|---|---|
| 1 | JSPM 1 | Bacillus spp. |
| 2 | JSPM 2 | Enterobacter spp |
| 3 | JSPM3 | Enterobacter spp |
| 4 | JSPM4 | Bacillus spp. |
| 5 | AAS1 | Pseudomonas aeruginosa |
| 6 | AAS 2 | Staphylococcus aureus |
| 7 | AAS3 | Pesudomonas spp. |
| 8 | AAS4 | Staphylococcus aureus |
| 9 | NGPM1 | Staphylococcus aureus |
| 10 | NGPM2 | Bacillus spp. |
| 11 | NGPM3 | Bacillus subtilis |
| 12 | NGPM4 | Staphylococcus aureus |
| 13 | NBAS 1 | Pseudomonas spp. |
| 14 | NBAS 2 | Flavobacterium spp. |
| 15 | NBAS 3 | Pseudomonas spp. |
| 16 | NBAS 4 | Flavobacterium spp. |
| 17 | NGFP1 | Streptococcus spp. |
| 18 | NGFP2 | Pseudomonas aeruginosa |
| 19 | NGFP3 | Pseudomonas aeruginosa |
| 20 | NGFP4 | Streptococcus spp. |
| 21 | JSFP1 | Bacillus spp. |
| 22 | JSFP2 | Pseudomomnas spp. |
| 23 | JSFP3 | Bacillus subtilis |
| 24 | JSFP4 | Pseudomonas spp. |
| 25 | WFP1 | Pseudomonas spp. |
| 26 | WFP2 | Bacillus spp. |
| 27 | WFP3 | Bacillus subtilis |
| 28 | WFP4 | Staphylococcus spp. |
| 1 | JSPM1 | Penicillium spp. |
| 2 | JSPM2 | Aspergillus terreus |
| 3 | JSPM3 | Candida spp. |
| 4 | JSPM4 | Aspergillus fumigatus |
| 5 | AAS1 | Mucor spp. |
| 6 | AAS 2 | Aspergillus niger |
| 7 | AAS3 | Penicillium chrysogenum |
| 8 | NGPM1 | Penicillium spp. |
| 9 | NGPM2 | Aspergillus fumigatus |
| 10 | NGPM3 | Mucor spp. |
| 11 | NGPM4 | Aspergillus terreus |
| 12 | NGPM5 | Rhizopus spp. |
| 13 | NGPM6 | Aspergillus niger |
| 14 | NBAS 1 | Candida spp. |
| 15 | NBAS 2 | Penicillium chrysogenum |
| 16 | NBAS 3 | Aspergillus fumigatus |
| 17 | NGFP1 | Rhizopus spp. |
| 18 | JSFP1 | Neurospora spp. |
| 19 | WMFP2 | Neurospora spp. |
Table 2: Diversity and distribution of bacteria isolates from different oil contaminated sites on Nutrient agar.
JSPM: Joseph Sarwuan Tarka University Oil Palm Mill; NGPM: New GRA Oil Palm Mill; NBAS: North Bank Auto Mechanic Site; AAS: Apir Auto mechanic Site; NGFP: New GRA Fish Pond; JSFP: Joseph Sarwuan Tarka University Fish Pond; WFP: Wurukum Fish Pond; WMFP: Wurukum Market Fish Processing Water. Table 2: Diversity and distribution of bacteria isolates from different oil contaminated sites on Nutrient agar.
JSPM: Joseph Sarwuan Tarka University Oil Palm Mill; NGPM: New GRA Oil Palm Mill; NBAS: North Bank Auto Mechanic Site; AAS: Apir Auto mechanic Site; NGFP: New GRA Fish Pond; JSFP: Joseph Sarwuan Tarka University Fish Pond; WFP: Wurukum Fish Pond; WMFP: Wurukum Market Fish Processing Water. Table 3: Diversity and distribution of fungi isolates from different oil contaminated sites on on Potato Dextros agar.
| Code Isolate | Isolate | Cell shape | Gram Reaction | Cit. | Coa. | Cat. | Indol. | TS.I. | H2S production | Lac. | Mal. | Fruc. | Suc. | Glu. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| JSPM1 | Bacillus spp. | Rod | + | + | - | + | - | K/A | - | - | + | + | + | + |
| JSPM2 | Enterobacter spp. | Rod | - | + | - | + | - | A/A | - | + | + | + | + | + |
| JSPM3 | Enterobacter spp. | Rod | - | + | - | + | - | A/A | - | + | + | + | + | + |
| JSPM4 | Bacillus spp. | Rod | + | + | - | + | - | K/A | - | - | + | + | + | + |
| AAS1 | Pseudomonas aeruginosa | Rod | - | + | - | + | - | K/K | - | - | - | + | - | - |
| AAS2 | Staphylococcus aureus | Cocci | + | - | + | + | - | A/A | - | + | + | + | + | + |
| AAS3 | Pseudomonas spp. | Rod | - | + | - | + | - | K/K | - | - | - | + | - | - |
| AAS4 | Staphylococcus aureus | Cocci | + | - | + | + | - | A/A | - | + | + | + | + | + |
| NGPM1 | Staphylococcus aureus | Cocci | + | - | + | + | - | A/A | - | + | + | + | + | + |
| NGPM2 | Bacillus spp. | Rod | + | + | - | + | - | K/A | - | - | + | + | + | + |
| NGPM3 | Bacillus subtilis | Rod | + | + | - | + | - | K/A | - | - | + | + | + | + |
| NGPM4 | Staphylococcus aureus | Cocci | + | - | + | + | - | A/A | - | + | + | + | + | + |
| NBAS1 | Pseudomonas spp. | Rod | - | + | - | + | - | K/K | - | - | - | + | - | - |
| NBAS2 | Flavobacterium spp. | Rod | - | - | - | - | - | K/A | - | - | - | + | - | + |
| NBAS3 | Pseudomonas spp. | Rod | - | + | - | + | - | K/K | - | - | - | + | - | - |
| NBAS4 | Flavobacterium spp. | Rod | - | - | - | - | - | K/A | - | - | - | + | - | + |
| NGFP1 | Streptococcus spp. | Cocci | + | - | - | - | - | A/A | - | + | + | + | + | + |
| NGFP2 | Pseudomonas aeruginosa | Rod | - | + | - | + | - | K/K | - | - | - | + | - | - |
| NGFP3 | Pseudomonas aeruginosa | Rod | - | + | - | + | - | K/K | - | - | - | + | - | - |
| NGFP4 | Streptococcus spp. | Cocci | + | - | - | - | - | A/A | - | + | + | + | + | + |
| JSFP1 | Bacillus spp. | Rod | + | + | - | + | - | K/A | - | - | + | + | + | + |
| JSFP2 | Pseudomonas spp. | Rod | - | + | - | + | - | K/K | - | - | - | + | - | - |
| JSFP3 | Bacillus subtilis | Rod | + | + | - | + | - | K/A | - | - | + | + | + | + |
| JSFP4 | Pseudomonas spp. | Rod | - | + | - | + | - | K/K | - | - | - | + | - | - |
| WFP1 | Pseudomonas spp. | Rod | - | + | - | + | - | K/K | - | - | - | + | - | - |
| WFP2 | Bacillus spp. | Rod | + | + | - | + | - | K/A | - | - | + | + | + | + |
| WFP3 | Bacillus subtilis | Rod | + | + | - | + | - | K/A | - | - | + | + | + | + |
| WFP4 | Staphylococcus spp. | Cocci | + | - | + | + | - | A/A | - | + | + | + | + | + |
Table 3: Morphological and biochemical characterization of bacteria isolated from oil contaminates samples and sites.
Positive: +; Negative: -; A: Acidic reaction (yellow); K: Alkaline reaction (red); A/K: acidic butt and alkaline slant; A/A: both slant and butt acidic; K/A: alkaline butt, acidic slant; K/K: both butt and slant alkaline. Table 4: Morphological and biochemical characterization of bacteria isolated from oil contaminates samples and sites.
| S/N | Code | Isolate | Margin | Elevation/Appearance | Colour |
|---|---|---|---|---|---|
| 1 | JSPM1 | Penicillium spp. | Entire | Umbonate elevation with velvety-powdery texture, reverse color (bottom of colony) | blue-green with white tinges |
| 2 | JSPM2 | Aspergillus terreus | Entire | Umbonate elevation, no cotton growth | cinnamon-brown |
| 3 | JSPM3 | Candida spp. | Entire | mucoid with no mycelia growth | creamy coloured |
| 4 | JSPM4 | Aspergillus fumigatus | Entire | Umbonate elevation with twisted colony | dark green |
| 5 | AAS1 | Mucor spp. | Filiform | Raised elevation with fine cotton growth | creamy color |
| 6 | AAS 2 | Aspergillus niger | Entire | Umbonate elevation with black spore heads | black coloured |
| 7 | AAS3 | Penicillium chrysogenum | Entire | Umbonate elevation with velvety-powdery texture, reverse color (bottom of colony - pale yellow) | green in center with white edges. |
| 8 | NGPM1 | Penicillium spp. | Entire | Umbonate elevation with velvety-powdery texture reverse color (bottom of colony), | blue-green with white tinges |
| 9 | NGPM2 | Aspergillus fumigatus | Entire | Umbonate elevation with twisted colony | dark green |
| 10 | NGPM3 | Mucor spp. | Filiform | Raised elevation with fine cotton growth | creamy color |
| 11 | NGPM4 | Aspergillus terreus | Entire | Umbonate elevation, no cotton growth | cinnamon-brown |
Table 4: Cultural and morphological characterization of fungi isolated from oil contaminates samples and sites.
- 12
- NGPM5
- Rhizopus spp.
- Filiform
- Raised elevation, cotton growth with spore heads creamy-brown
- 13
- NGPM6 Aspergillus niger
- Entire
- Umbonate elevation with black spore heads black coloured
- 14
- NBAS 1
- Candida spp.
- Entire mucoid with no mycelia growth creamy coloured
- Penicillium chrysogenum
- Entire
- Umbonate elevation with velvety-powdery texture, reverse
- 15
- NBAS 2 color (bottom of colony - pale yellow)
- Aspergillus fumigatus
- Entire
- Umbonate elevation with twisted colony dark green
- 16
- NBAS 3
- 17
- NGFP1
- Rhizopus spp.
- Filiform
- Raised elevation, cotton growth with spore heads creamy-brown
- 18
- JSFP1
- Neurospora spp. Filiform
- Raised elevation with cotton growth orange color
- 19
- WMFP2
- Neurospora spp. Filiform
- Raised elevation with cotton growth orange color
Table 5: Cultural and morphological characterization of fungi isolated from oil contaminates samples and sites.
Screening for lipolytic activity on Tween 80 agar revealed that all bacterial isolates exhibited varying degrees of activity, characterized by progressive zones of clearance. Bacillus spp. demonstrated the high activity with zones of 44-62 mm, peaking at 120 hours. Pseudomonas aeruginosa and Staphylococcus aureus followed similar trends, reaching 58 mm and 55 mm at peak activity, respectively (Figure 1). There was significant difference in the lipolytic activity of the isolates at p˂0.05. In contrast, lipase activity on Phenol Red Olive Oil Agar was significantly lower (p˂0.05), with no activity observed from 24 to 72 hours. Flavobacterium spp. exhibited minimal activity (5-9 mm) at 96-120 hours, while Pseudomonas aeruginosa and Enterobacter spp. produced zones of 4-8 mm and 4-7 mm, respectively (Figure 2). Fungal screening on Tween 80 agar showed progressive lipolytic activity from 24 to 72 hours. Neurospora spp., Aspergillus flavus, and Aspergillus niger exhibited the high clearance zones of 33-83 mm, 17-83 mm, and 13-83 mm, respectively, while Aspergillus terreus and Rhizopus spp. produced zones of 79 mm and 72 mm (Figure 3). Lipase production on Phenol Red Olive Oil Agar was generally higher, with Aspergillus flavus (44-83 mm), Rhizopus spp. (38-83 mm), and Aspergillus niger (38-83 mm) showing the most significant activity (p˂0.05). Mucor spp. recorded the low activity (2-17 mm) (Figure 4).
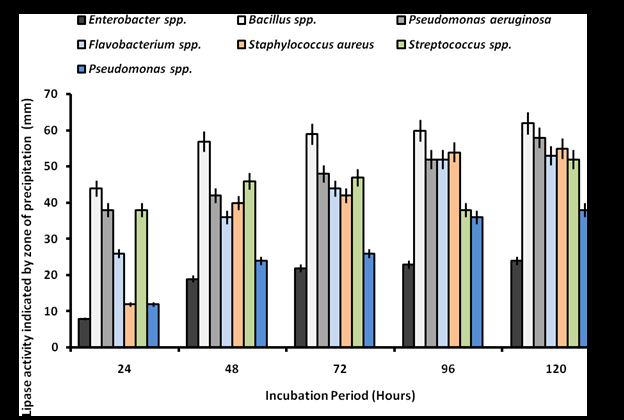
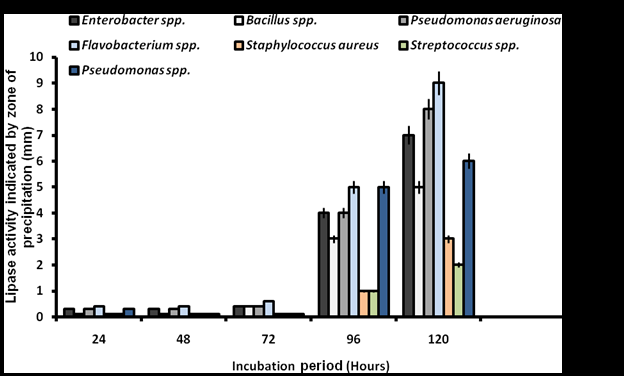
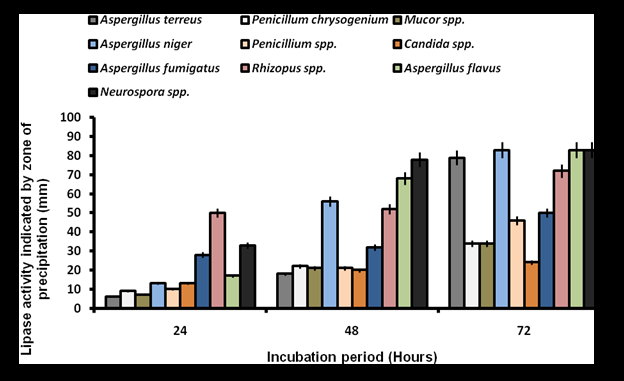
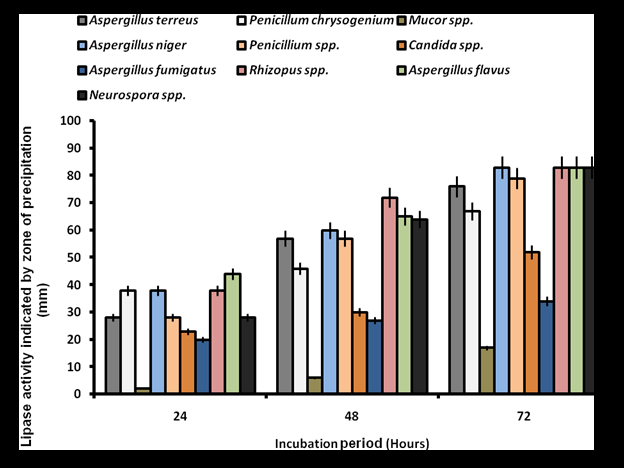
These findings indicate that both bacteria and fungi isolated from oil-contaminated sites in Makurdi exhibit significant lipolytic potential, with notable differences in sensitivity to Tween 80 and Phenol Red Olive Oil Agar-based screening.
Discussion
The presence of microbial isolates with significant lipolytic activity from oil-rich sites in Makurdi metropolis highlights their potential role in bioremediation and industrial applications. The observed bacterial and fungal diversity, coupled with their varying lipase production on different screening media, emphasizes the metabolic adaptability of these microorganisms in oil-contaminated environments.
Microbial Diversity and Distribution
The study revealed a diverse range of bacterial and fungal species isolated from oil-contaminated sites. The predominant bacterial genera included Bacillus, Pseudomonas, Enterobacter, Staphylococcus, Flavobacterium, and Streptococcus, while the fungal isolates comprised species of Aspergillus, Penicillium, Rhizopus, Neurospora, and Mucor (Alabdalall et al., 2020; Ali et al., 2023). The prevalence of Pseudomonas spp. and Bacillus spp. in multiple locations is consistent with previous studies highlighting their ability to degrade hydrocarbons and thrive in petroleum- polluted environments [1, 11, 12]. Staphylococcus aureus was also prevalent, suggesting possible anthropogenic contamination from human activities [13]. Among the fungal isolates, Aspergillus spp. and Penicillium spp. were the most frequently encountered [3], aligning with findings from other studies that have identified these genera as dominant in oil-contaminated environments due to their enzymatic capabilities and hydrocarbon degradation potential [14, 15]. The presence of Neurospora spp. and Rhizopus spp. further suggests the involvement of fungi in lipid metabolism and organic matter decomposition.
Lipase Activity on Tween 80 and Phenol Red Olive Oil Agar
The screening of microbial isolates for lipase production revealed significant variations in enzyme activity across bacterial and fungal species. On Tween 80 agar, bacterial isolates exhibited differential lipolytic activity, with Bacillus
spp. demonstrating the highest lipolytic activity, followed by Pseudomonas aeruginosa and Staphylococcus aureus (p < 0.05). The strong lipolytic activity of Bacillus spp. corroborates previous reports of its industrial application in enzyme production [7, 16]. Fungal isolates generally exhibited higher lipase activity compared to bacteria, with Neurospora spp., Aspergillus flavus, and Aspergillus niger producing the highest clearance zones (ranging 33 mm - 83 mm). The ability of Aspergillus spp. to produce high lipase yields is well- documented in enzyme biotechnology [3, 4]. These enzymes do not require cofactors to enhance their catalytic efficiency, making them highly effective in various reactions. Due to their versatility, fungal lipases are widely recognized as potential biocatalysts in both industrial and biotechnological applications. When screened on Phenol Red Olive Oil Agar, bacterial isolates exhibited significantly lower lipolytic activity (p < 0.05), with no enzyme production detected within the first 72 hours [17]. Flavobacterium spp. produced the lowest clearance zone (9 mm at 120 hours), followed by Pseudomonas aeruginosa (8 mm) and Enterobacter spp. (7 mm). This suggests that Phenol Red Olive Oil Agar is less sensitive for bacterial lipase detection. In contrast, fungal isolates exhibited significantly higher enzyme production on Phenol Red Olive Oil Agar (p < 0.05). Aspergillus flavus, Rhizopus spp., and Aspergillus niger recorded the highest clearance zones (83 mm), confirming the higher sensitivity of Phenol Red Olive Oil Agar for fungal lipase screening. However, Mucor spp. exhibited the lowest lipase activity (17 mm), suggesting variable enzyme production among fungal species.
Comparative Sensitivity of Screening Media
The results indicate that Tween 80 agar is a more effective medium for detecting bacterial lipase activity, as the clearance zones around bacterial colonies are more pronounced on this medium. This increased activity can be attributed to variations in bacterial response based on factors such as substrate specificity, pH preference, and detection methods. During growth, bacterial lipases efficiently hydrolyze Tween 80, leading to the formation of clear zones around the colonies [4]. Additionally, species like Bacillus, Pseudomonas, and Staphylococcus thrive in the neutral to alkaline pH range of 7.0 to 9.0 [18] in this medium, further enhancing lipase detection. The use of Tween 80 as a substrate allows for effective detection of extracellular lipases, as it presents an easily hydrolyzable ester bond suitable for enzyme action [4]. Conversely, fungi exhibit higher lipolytic activity on Phenol Red Olive Oil Agar, which utilizes a pH- sensitive detection system. Fungal lipases generate fatty acids that lower the pH, causing a visible colour change in phenol red. This medium supports acidic conditions (4.5 - 6.5) optimal for fungal enzyme production and allows for longer incubation periods, facilitating extracellular lipase accumulation. The study demonstrated that lipolytic activity of bacterial and fungal isolates on Tween 80 and Phenol Red Olive Oil Agar was significantly different (p < 0.05). This aligns with general fact that bacterial lipases function optimally in neutral to alkaline pH conditions, whereas acidic pH levels promote the production of fungal lipases [4]. This explains the greater fungal lipase activity on Phenol Red Olive Oil Agar. These findings stress the need for multiple screening methods when evaluating microbial lipase activity, as different organisms exhibit varying enzymatic responses depending on the substrate and detection system.
Industrial and Environmental Implications
The ability of bacterial and fungal isolates from oil- rich environments to produce lipases highlights their biotechnological potential in industrial and environmental applications. Lipases are widely utilized in bioremediation, detergent formulation, pharmaceuticals, and food processing [19]. The strong lipolytic activity of Bacillus spp., Pseudomonas spp., Aspergillus spp., and Rhizopus spp. suggests their suitability for enzyme production at an industrial scale. Additionally, the presence of these microorganisms in oil-contaminated sites suggests their role in natural biodegradation processes, making them potential candidates for bioremediation strategies aimed at cleaning up oil spills and hydrocarbon pollution. Previous studies have demonstrated that Pseudomonas spp. and Bacillus spp. are among the most efficient microbial degraders of hydrocarbons [1, 12].
Conclusion and Future Prospects
This study demonstrates the significant lipolytic potential of bacterial and fungal isolates from oil- contaminated sites in Makurdi metropolis. The differential enzyme activity observed on Tween 80 and Phenol Red Olive Oil Agar suggests that multiple screening techniques are necessary to obtain a comprehensive assessment of microbial lipase production. The higher lipase activity observed in fungal isolates, particularly Aspergillus flavus, A. niger, and Rhizopus spp., highlights their suitability for industrial enzyme applications. Future research should focus on optimizing culture conditions to enhance lipase production, genetic characterization of high-yielding isolates, and pilot-scale enzyme production for potential industrial and bioremediation applications.
References 1. Ali S, Khan SA, Hamayun M, Lee IJ (2023) The Recent Advances in the Utility of Microbial Lipases: A Review. Microorganisms 11(510): 510.
2. Yao W, Liu K, Liu H, Jiang Y, Wang R, et al. (2021) A
Valuable Product of Microbial Cell Factories: Microbial Lipase. Front Microbiol 12: 743377.
3. Alabdalall AH, ALanazi NA, Aldakeel AS, AbdulAzeez S, Borgio JF (2020) Molecular, physiological, and biochemical characterization of extracellular lipase production by Aspergillus niger using submerged fermentation. PeerJ 8: e9425.
4. Kumar A, Verma V, Dubey VK, Srivastava A, Garg SK, et al. (2023) Industrial applications of fungal lipases: a review. Front Microbiol 14: 1142536.
5. Ramnath L, Sithole B, Govinden R (2017) Identification of lipolytic enzymes isolated from bacteria indigenous to Eucalyptus wood species for application in the pulping industry. Biotechnology Reports 15: 114-124.
6. Abubakar A, Abioye OP, Aransiola SA, Maddela NR, Prasad R (2024) Crude oil biodegradation potential of lipase produced by Bacillus subtillis and Pseudomonas aeruginosa isolated from hydrocarbon contaminated soil. Enviromental chemistry and Ecotoxicology 6: 26- 32.
7. Lee LP, Karbul HM, Citartan M, Gopinath SC, Lakshmipriya T, et al. (2015) Lipase-Secreting Bacillus Species in an Oil-Contaminated Habitat: Promising Strains to Alleviate Oil Pollution. Biomed Res Int 2015: 820575.
8. British Mycological Society (2025) Keys.
9. Holt JG, Krieg NR, Sneath PHA, Staley JT, Willaims ST, et al. (1994) Bergey’s manual of determinative bacteriology. 9th (Edn.), Willaims & Wilkins.
10. Duncan DB (1955) Multiple range and multiple F-test.
Biometrics 11: 1-42.
11. AI-Jawhari IFH, Flayyih AI, Mohammed DQ (2022) Ability
of Pseudomonas aeruginosa and Bacillus subtilis to remove petroleum hydrocarbons. International Journal of Life Science Research Archive 3(2): 90-95.
12. Okoye AU, Selvarajan R, Chikere CB, Okpokwasili GC,
Mearns K (2024) Characterization and identification of long-chain hydrocarbon-degrading bacterial communities in long-term chronically polluted soil in Ogoniland: an integrated approach using culture- dependent and independent methods. Environ Sci Pollut Res 31: 30867-30885.
13. Fritz SA, Hogan PG, Singh LN, Thompson RM, Wallace
MA, et al. (2014) Contamination of environmental surfaces with Staphylococcus aureus in households with children infected with methicillin-resistant S aureus. JAMA Pediatr 168(11): 1030-1038.
14. Zhang J, Xue Q, Gao H, Ma X, Wang P (2015) Degradation of crude oil by fungal enzyme preparations from Aspergillus spp. for potential use in enhanced oil recovery. Journal of Chemical Technology and Biotechnology 91(4): 865- 875.
15. Al-Hawash AB, Alkooranee JT, Abbood HA, Zhang J, Sun J,
et al. (2018) Isolation and characterization of two crude oil-degrading fungi strains from Rumaila oil field, Iraq. Biotechnology Reports 17: 104-109.
16. Kazeem MO, Mayaki MA, Musa D (2024) Response Surface
Optimization of Lipase Production by Pseudomonas sp. on a Low-Cost Shea-nut Cake using Solid State Fermentation. Ife Journal of Science 26(1): 1-12.
17. Alkabee HJJ, Alsalami AK, Alansari BMM (2020)
Determination the Lipase Activity of Staphylococcus sp. Strain Isolated from Clinical Specimens. J Pure Appl Microbiol 14(1): 437-446.
18. Abel ES, Evivie SE (2022) Effect of Varying Environmental
Conditions on the Growth and Viability of Selected Microorganisms using Conventional Cultures. J Appl Sci Environ Manage 26(3): 393-397.
19. Patel N, Rai D, Shahane S, Soni S, Mishra U (2018) Lipases:
Sources, Production, Purification, and Applications. Recent Patents on Biotechnology 12(1): 1-12.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania
- Immobilization of Trametes Versicolor Laccase onto Silica Aerogel via Covalent Attachment Method