Natural Killer Cells: From Bone Marrow to Cytotoxic Cells
Natural killer (NK) cells are lymphoid cells, having central effector functions in immune system without previous sensitization. These cells are related to innate lymphoid cells which are characterized by lymphoid cell morphology with missing antigenspecific receptors. NK cells are defined by lack of expression of CD3 and expression of CD56 (CD3-CD56+). Based on the expression of CD56, human NK cells are classified into two main populations: CD56bright and CD56dim NK cells. Functionally, CD56bright subset is characterized by higher capacity to produce cytokines following activation but has lower cytotoxicity. On the contrary, CD56dim NK cells are more cytotoxic and express higher levels of maturation markers. NK cells have effective cytotoxic activity against virus-infected cells and tumors, and their effector functions are regulated by multiple activating and inhibitory receptors. The activated NK cells are able to kill their target cells in apoptosis process by different mechanisms, including cytolytic granule-dependent exocytosis pathway, death receptor pathway, antibody-dependent cell-mediated cytotoxicity, and the release of cytokines. This review will focus on NK cell differentiation, NK cell receptors as well as their ligands and their role in immune responses. In addition, we will summarize the pathways involved in NK cell effector functions.
Introduction
NK cells are bone marrow-derived lymphocytes arising from the lymphoid lineage which were identified in 1975 as effector lymphocytes due to their ability to lyse virus- infected cells and tumor cells [1, 2]. They develop during fetal life as well as after birth from CD34+ hematopoietic stem cells which differentiate into common lymphoid progenitors that can give rise to NK cell progenitor (Figure
1). It has been commonly reported that the bone marrow is considered to be the main site of NK cell development similar to B cells. However, some studies have notified that NK cell development and maturation can take place in other sites such as the thymus [3] and lymph nodes [4]. Despite their lymphoid origin, NK cells are crucial components of the innate immune defense due to the direct kill of their target cells in the absence of specific immunization and lack of antigen-specific cell surface receptors [5, 6]. Now, NK cells have been reclassified as a subset of cytotoxic innate lymphoid cells [6, 7]. However, they differ from other innate immune cells that they do not mediate phagocytosis and lack bactericidal enzymatic systems. Generally, the effector functions of NK cells are regulated by a balance between activating and inhibitory signals provided by a wide group of activating and inhibitory receptors. When NK cell activating receptors interact with their specific ligands expressed on the target cells, NK cells become cytotoxic cells and are able to kill their target cells by different mechanisms. In this review, we provide an overview of the characteristics of NK cells, their phenotype, differentiation and maturation, receptors and functions.
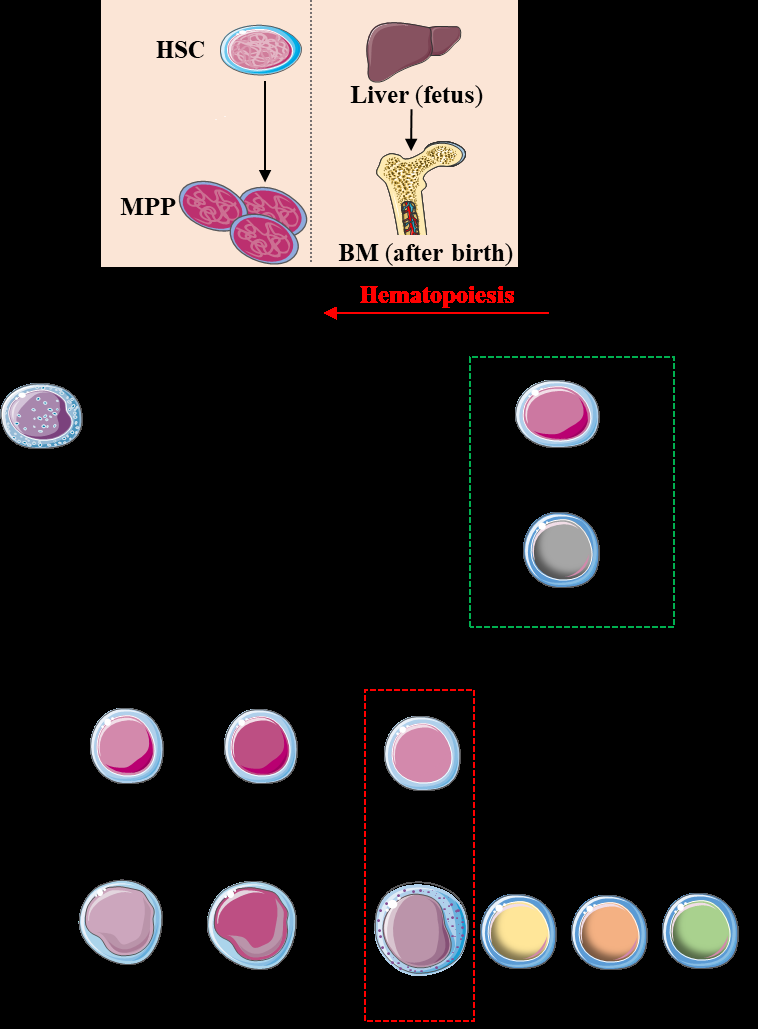
Hematopoietic stem cells (HSCs) divide into multipotent progenitors (MPPs) which lead to common myeloid progenitors (CMPs; the precursors of all myeloid cells) and common lymphoid progenitors (CLPs; the precursors of all lymphoid cells). Then CLPs give rise to the precursors of T cells (TCP), B cells (BCP) and common innate lymphoid cells precursors (CILCPs). The CILCPs lead to different groups of innate lymphoid cells including NK cells through their specific precursors. HSC: hematopoietic stem cell; MPPs: multipotential progenitors; BM: bone marrows; CMP: common myeloid progenitor; CLP: common lymphoid progenitor; CILP: common innate lymphoid progenitor; NKP: NK cell progenitor; TCP: T cell progenitor; BCP: B cell progenitor; ILC: innate lymphoid cells.
NK Cell Phenotype and Subsets
In humans, NK cells are considered as the third largest population of lymphocytes following T and B cells comprising approximately 10 – 15% of all peripheral blood lymphocytes [8, 9]. Phenotypically, they are defined as CD3-CD56+ lymphocytes upon their expression of CD56 (Neural Cell Adhesion Molecule, 140 kDa isoform) and lack of expression of the T cell marker CD3 [8]. Moreover, NK cells can be subdivided into various subsets with diverse functions based on the surface expression of CD56, CD16, inhibitory receptors and/or activating receptors. The main two populations of human NK cells are CD56bright CD16− and CD56dim CD16+ [8].
CD56bright CD16− NK cells are immature, cytokine- producing NK cells (5 – 10% of NK cells), being dominant in the lymph nodes and function as immunoregulatory cells by secreting cytokines such as interferon-γ (IFN-γ), tumor necrosis factor-α (TNF-α), interlukin -10 (IL-10), IL- 13 and granulocyte–macrophage colony-stimulating factor (GM-CSF). CD56bright CD16− cells highly express NKG2A inhibitory receptor and have low to absent expression of killer cell immunoglobulin-like receptors (KIRs) [8].
On the other hand, around 90 - 95% of peripheral blood NK cells are CD56dim CD16+ cells which are cytotoxic cells with weak proliferation activity and low levels of cytokines production at resting state [8, 10, 11]. However, CD56dim NK cells largely secrete proinflammatory cytokines and chemokines more than CD56bright cells after target cell stimulation.
NK Cell Differentiation and Maturation
The differentiation pathway of human NK cell is complex, characterized by losing the expression of specific surface antigens as CD34 and increasing the expression of other surface antigens such as CD56, NKG2A, CD16, KIRs
and CD57 resulting in a heterogeneous CD3−CD56+ NK cell population. Further, a variety of activating and inhibitory receptors are acquired or lost during NK cell maturation. There are many factors regulating NK cell differentiation, including transcription factors and cytokines [12]. Several studies have also shown the importance of microRNAs in the differentiation and maturation of NK cells [13].
Transcription Factors
Transcription factors have a prominent effect on immune cell development and differentiation by regulating gene transcription of specific molecules controlling these processes. There are numerous transcription factors involved in regulation of NK cell differentiation and maturation. For instance, an inhibitor of DNA binding protein (Id2) contains a helix-loop-helix domain which prevents the binding of transcription factors (E2A, E12 and E47) with DNA. Boos, et al. [14] demonstrated that Id2 has a role in NK cell differentiation at later stage because they found that Id2 deficient mice have low number of mature NK cells in the periphery while having normal number of immature NK cells and NK cell progenitors in the bone marrow [14].
Another example is E4-binding protein 4 (E4BP4, also called NFIL3) transcription factor that has a potential regulatory role in the immune system [15]. E4BP4-deficient mice showed a sharp reduction in both immature NK and mature NK numbers, suggesting that E4BP4 is required for the progression from NK cell precursors to immature NK cells and then from immature NK cells to mature NK cells [16, 17]. Recently, Male, et al. [18] showed that E4BP4 is necessary for NK cell progenitor development from common lymphoid progenitors and for NK cell development at early stage by controlling the expression of Eomes and Id2 [18].
In addition, T-box transcription factors family including Eomes and T-bet (also known as Tbx21) are defined as basic drivers of immune cell development and cytolytic function. In humans, Knox et al. (2014) have found that Eomes was expressed in both CD56bright and CD56dim NK cells, but it was significantly higher in CD56bright suggesting that Eomes is likely essential for CD56bright maturation [19]. With respect to T-bet, it regulates the expression of sphingosine-1 phosphate receptor 5 (S1P5) which plays an important role in NK cell recirculation. T-bet–/ – mice exhibited a reduced number of NK cells in the spleen, liver and peripheral blood, and an increased number in lymph nodes and bone marrow [20, 21]. T-bet–/ – NK cells also showed an impaired cytotoxicity and IFN-γ production [20, 21].
Cytokines
Cytokines play an important regulatory role in several processes in the immune system, including development, proliferation, homeostasis, and activation status. In the context of NK cells, IL-2, IL-15 and IL-21 are the major cytokines that regulate NK cell development, differentiation, survival, and function. Their receptors are signaling through three main transduction pathways: the Janus tyrosine kinases (JAK)-STAT pathway, the phosphatidylinositol-3- kinase pathway, and the mitogen activated protein kinase (MAPK) pathway (Figure 2) [22, 23]. IL-18 can control NK cell proliferation and cytokine production by binding to IL-18 receptor which activates MAPK and nuclear factor- kappa B (NF-kB) pathways [24]. Table 1 summarizes the key interleukins regulating NK cell differentiation and other biological functions.
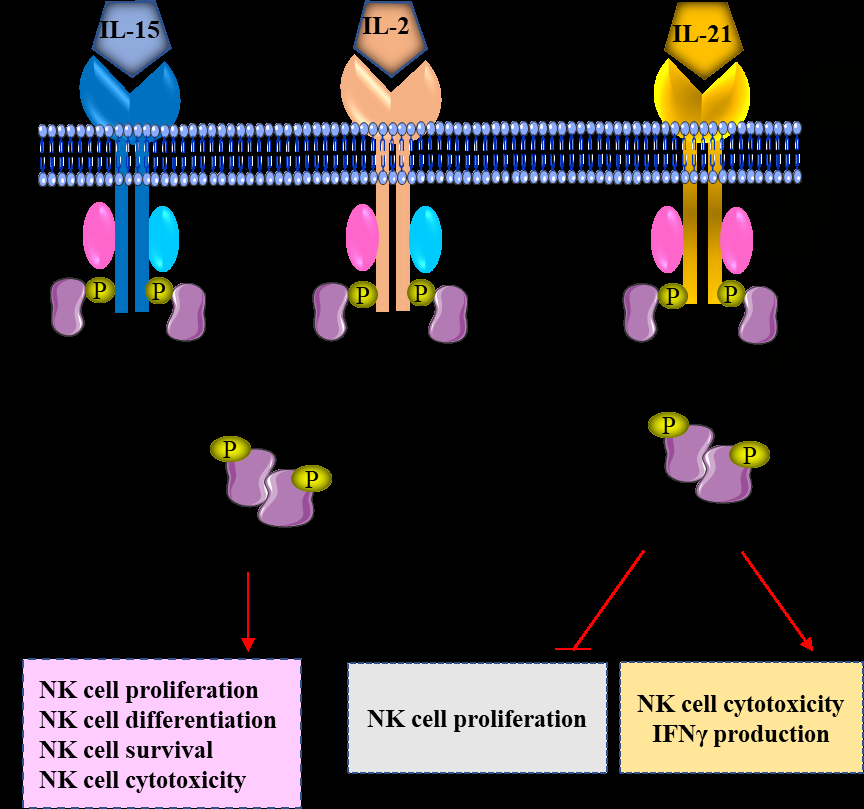
Different cytokines can control NK cells through their receptors. When the cytokines bind with their specific receptors, they can activate specific signaling pathways regulating NK cell biology. IL: interleukin; JAK: Janus kinase; STAT: signal transducer and activator of transcription; IFNγ: interferon-γ.
| Source | Signaling | Function | |
|---|---|---|---|
| IL-2 | T cells | JAK1/3 | Cytokine production, proliferation, survival, enhanced cytotoxicity |
| IL-2 | T cells | STAT3/5 | Cytokine production, proliferation, survival, enhanced cytotoxicity |
| IL-2 | T cells | MAPK | Cytokine production, proliferation, survival, enhanced cytotoxicity |
| IL-7 | Stromal cells | JAK1/3 | Development of thymic NK cells, proliferation |
| IL-7 | Stromal cells | STAT5 | Development of thymic NK cells, proliferation |
| IL-12 | DCs | JAK2 | Differentiation, proliferation, cytokine production, enhanced cytotoxicity |
| IL-12 | Macrophages | STAT3/4/5 | Differentiation, proliferation, cytokine production, enhanced cytotoxicity |
| IL-15 | DCs | JAK1/3 | Cytokine production, proliferation, survival, maturation, enhanced cytotoxicity |
| IL-15 | Macrophages | STAT3/5 | Cytokine production, proliferation, survival, maturation, enhanced cytotoxicity |
| IL-15 | Stromal cells | MAPK | Cytokine production, proliferation, survival, maturation, enhanced cytotoxicity |
| IL-18 | DCs | MyD88 | |
| IL-18 | Macrophages | MAPK | Cytokine production, proliferation |
| IL-18 | NF-kB | ||
| IL-21 | T cells | JAK1/3 | Differentiation, enhanced cytotoxicity, cytokine production, limits proliferation |
| IL-21 | T cells | STAT1/3/5 | Differentiation, enhanced cytotoxicity, cytokine production, limits proliferation |
Table 1: Major interleukins regulating NK cell differentiation and biological functions. IL: Interlukin; JAK: Janus tyrosine kina
MicroRNAs
MicroRNAs are a large family of short non-coding (22 nucleotides) RNA molecules that regulate gene expression post-transcriptionally by targeting the 3’ untranslated regions of mRNAs, resulting in translational inhibition and/or mRNA degradation. There are several microRNAs affecting the development and maturation of NK cells. For example, a consistent increase in the expression levels of microR-181a/b is associated within different stages of NK cell development, suggesting that microR-181 has an impact on human NK cells development from CD34+ hematopoietic progenitors [25]. MicroR-181 establishes its effect on NK cells development through downregulation of nemo-like kinase (a protein kinase that negatively regulates Notch- dependent transcriptional activation pathway). Another example is microRs-15/16 which are highly expressed in NK cells and are involved in regulation of NK cell development by controlling the level of Myb (also known as c-Myb) [26]. Myb is a transcriptional activator factor related to myeloblastosis family that is highly expressed in immature NK cells (CD56bright NK cells), however, its expression is decreased during NK cell maturation (CD56dim NK cells). It has been found that immature NK cells are accumulated, while mature NK cells are reduced in the absence of microR-15/16 [26]. Further, it was noted by Mundy-Bosse, et al. [27] that immature NK cells were extremely reduced in acute myeloid leukemia patients due to a disruption in NK cell differentiation [27]. They also found low levels of T-bet and Eomes transcription factors as a result of elevation level of microR-29b in NK cells of leukemia patients (microR- 29b is a regulator of T-bet and Eomes). As a consequence of microR-29b deletion in NK cells, the immature NK subset is returned to appear again [27].
NK Cell Receptors
During NK cell maturation, they acquire a series of cell surface molecules that regulate NK cell effector functions. These molecules can be classified into different groups: inhibitory receptors, activating receptors, adhesion molecules, cytokine and chemotactic receptors.
Inhibitory Receptors
NK cell functions are negatively controlled by many inhibitory receptors where CD94/NKG2A and inhibitory KIRs are the major inhibitory receptors [28, 29]. Inhibitory
receptors transmit their inhibitory signals through intracellular immunoreceptor tyrosine-based inhibitory motifs (ITIMs), located in the cytoplasmic tail of these receptors. Following binding the ligand with its receptor, the tyrosine residues in ITIMs are phosphorylated, and thus recruit and activate Src homology 2 domains of the intracytoplasmic protein tyrosine phosphatases SHP-1 and SHP-2. Then, these phosphatases inhibit the activation of NK cells by dephosphorylation of specific intracellular signaling molecules at multiple stages of signaling cascade [30, 31].
CD94/NKG2A
NKG2A is a transmembrane protein structurally characterized by C-type lectin extracellular domains, covalently coupled to CD94 subunit [32, 33]. CD94/NKG2A receptor is found as a heterodimer that recognizes non- classical human leukocyte antigen (HLA) class I molecule, HLA-E [34]. It is naturally expressed on about half of all NK cells, and its expression is not stable and can be affected by cytokines in the surrounding environment. CD94 has a short cytoplasmic domain lacking signaling function, whereas NKG2A contains ITIM domain in its intracellular structure, which upon tyrosine phosphorylation can recruit SHP-1 or SHP-2 providing inhibitory signal transduction [32, 33].
Inhibitory KIRs
KIRs (CD158) are transmembrane glycoproteins, encoded on chromosome 19q13.4, and belong to immunoglobulin (Ig) superfamily. KIRs family are classified into 14 groups (2DL1 to 2DL5, 3DL1 to 3DL3, 2DS1 to 2DS5, and 3DS1) according to the number of extracellular Ig-like domains, and cytoplasmic tail length (Table 2) [35]. They are monomeric receptors, structurally characterized by either 2 extracellular Ig-like domains (designated D1 and D2 in KIR2D) or 3 extracellular Ig-like domains (designated D0, D1 and D2 in KIR3D). However, these receptors have differences in their cytoplasmic domains and are subdivided into KIRs with long (L) cytoplasmic tails (KIR2DL and KIR3DL) and others with short (S) cytoplasmic tails (KIR2DS and KIR3DS). Based on these variances, KIRs are classified to two functionally distinct groups: inhibitory KIRs as KIR2DL and KIR3DL where the long tails generate an inhibitory signal, and activating KIRs with short tails as KIR2DS and KIR3DS [36].
| KIR | Ligand | |
|---|---|---|
| Inhibitory KIRs | 2DL1 | HLA-C group 2 |
| 2DL2/3 | HLA-C group 1, B46, B73 and some HLA-C group 2 | |
| 3DL1 | HLA-Bw4 | |
| 3DL2 | HLA-A3, A11 | |
| 3DL4 | HLA-G | |
| 3DL5 | Not known | |
| 3DL3 | Not known | |
| Activating KIRs | 2DS1 | HLA-C group 2 |
| 2DS2 | Not known | |
| 2DS3 | Not known | |
| 2DS4 | HLA-A11 and subsets of HLA-C | |
| 2DS5 | Not known | |
| 3DS1 | Not known |
Table 2: Activating and inhibitory KIRs and their ligands.
Inhibitory KIRs signal through their long tails and ITIMs in the cytoplasmic domain. Following engagement with their ligands, the tyrosine residues of ITIMs are phosphorylated and then recruit protein tyrosine phosphatases as SHP- 1, which are critical for mediating inhibitory function. The known ligands for inhibitory KIRs are HLA-class I (HLA-A, B and C) molecules. In contrast to other inhibitory KIRs, engagement of KIR2DL4 by its ligand results in increasing IFN-γ production without activation of cytotoxicity [37].
Activating Receptors
NK cells express a wide group of activating receptors, which upon ligation transmit activating intracellular signals to initiate the effector functions of NK cells. The major activating receptors are NKG2D (Natural-Killer Group 2, member D), natural cytotoxicity receptors (NCRs), DNAX accessory molecule-1 (DNAM-1), CD16 (Fcγ RIII) and activating KIRs [28]. In addition, several co-receptors act in coordination with the activating receptors to activate NK cells such as NKp80, CD2, CD160, and 2B4 [28]. Commonly, NK activating receptors signal through two prominent signaling pathways: immunoreceptor tyrosine-based activating motifs (ITAMs) pathway, and DNAX-activation protein-10 (DAP-10) pathway.
NKG2D
NKG2D is one of the most important activating receptors expressed by NK cells. It is a C-type lectin surface receptor which was identified in 1991 [38]. NKG2D binds to induced- self ligands overexpressed on the abnormal cells, including MHC class I-chain-related proteins A and B (MICA and MICB), and UL16-binding proteins (ULBPs) [39, 40]. There is no ITAM signaling motif in the intracellular domain of NKG2D, therefore, it signals through its association with adaptor protein called DAP10. When the receptor is engaged, this initiates downstream signaling pathways that promote NK cell degranulation and production of cytokines and chemokines such as IFN-γ, TNF-α, GM-CSF, CCL4 and CCL1 [41].
Natural Cytotoxicity Receptors (NCRs)
Natural cytotoxicity receptors (NCRs) are exclusively expressed by NK cells and mainly involved in the mechanisms by which NK cells kill their targets. These receptors, are non-MHC class-I activating receptors and belong to Ig superfamily. In humans, three NCRs (NKp46, NKp44, and NKp30) have been recognized. NKp46 (NCR1 or CD335) and NKp30 (NCR3 or CD337) are constitutively expressed by all peripheral blood NK cells [42, 43], while NKp44 (NCR2 or CD336) is expressed on NK cells mainly after IL-2 stimulation [44]. NCRs include an extracellular ligand- binding domain and a transmembrane domain, while they lack functional cytoplasmic tails. So, each receptor is coupled with single dimeric ITAM-containing adaptor proteins (CD3ζ, FcRIγ and DAP12) to generate their activating signals. Among NCRs, NKp30 and NKp46 associates with FcRγ and/or CD3ζ, whereas DAP12 is connected with NKp44 [45, 46]. Engagement of NCRs by their ligands results in phosphorylation of tyrosine residue in ITAMs of the adaptor molecules by Src family kinases. The phosphorylated ITAMs recruit and activate tyrosine kinases as zeta chain-associated protein kinase 70 (ZAP70). Consequently, these kinases phosphorylate other signaling molecules which lead to increasing cytokines secretion such as IFN-γ and TNF-α and activation of cellular cytotoxicity [47, 48].
Activating KIRs
In contrast to inhibitory KIRs, KIRs with short cytoplasmic tail are activating KIRs. These receptors are associated with ITAM-bearing DAP12 adaptor protein. When activating KIRs are ligated, src family kinases phosphorylate the ITAM- containing adaptor molecule DAP12. Consequently, DAP12 activates ZAP70 and spleen tyrosine kinase which generate the downstream activation cascade [49]. There are several activating KIRs such as KIR2DS1 which has similar Ig-like domains to inhibitory KIR2DL1 and also binds HLA-C2 [50]. KIR2DS4 is the oldest and most predominant activating KIR receptor, having the ability to bind with C1 and C2 epitopes of HLA-C allotypes and HLA-A11 [51].
CD16A (FcγRIIIA)
In humans, several constant fragment gamma receptors (FcγRs) are found on the surface of NK cells, dendritic cells, neutrophils, monocytes, macrophages, B cells and some CD8+ T cells including activating receptors such as (FcγRIIA, FcγRIIC, FcγRIIIA and FcγRIIIB) and a single inhibitory receptor, FcγRIIB [52]. These receptors bind to Fc portion of IgG to initiate intracellular signaling pathways. CD16 is a low-affinity IgG Fc receptor (FcγRIII) that is expressed on the surface of NK cells, neutrophils, dendritic cells, monocytes and macrophages [52]. It is responsible on a mechanism of immune defense called antibody-dependent cell-mediated cytotoxicity (ADCC). There are two isoforms of CD16, FcγRIIIA (CD16A) which is expressed mainly on majority of CD56dim NK cells, and FcγRIIIB (CD16B) which is found on neutrophils. Once CD16A binds to the Fc region of IgG, NK cells release cytotoxic granules containing perforin and granzymes, causing lysis of target cells as tumors and virus- infected cells.
DNAM-1
DNAM-1 (CD226) is an activating receptor of the immunoglobulin superfamily expressed on NK cells, CD8+ T cells, and other immune cells. DNAM-1 recognizes two ligands related to nectin family: poliovirus receptor (CD155, also known as nectin-like molecule 5) and Nectin-2 (CD112) which are expressed by various healthy tissues as well as tumor cells [53, 54]. The interaction between DNAM-1 and its ligands boosts cytokines production and cell-mediated cytotoxicity against dendritic cells, tumor cells and virus infected cells [55, 56, 57].
Acquisition of NK Cell Functions
NK cells play a key role in immuno-surveillance and host defense against certain infected or transformed cells mediated by direct cytolysis via perforin/granzyme, death receptors (Fas, TRAIL) and/or ADCC pathways [58]. Alongside, NK cells secrete cytokines and chemokines which influence the host’s immune response by regulating other immune cells [59, 60]. NK cells have to be educated and primed to be able to recognize their target cells.
NK Cell Education
NK cells express many inhibitory receptors that recognize diverse self-molecules to prevent self-reactivity against healthy cells, however, self-major histocompatibility complex (MHC) class I molecules recognized by NK inhibitory receptors are essential for NK cell education [61, 62]. NK cell education, also known as licensing, is a process developed during NK cell maturation to acquire the effector functions that are adapted to the host in which they develop. It has become clear by several reports that binding of inhibitory receptors on NK cells by self-MHC class I molecules is important to determine whether an NK cell will be functionally capable of mediating missing-self recognition, or it will be hyporesponsive following stimulation. Interestingly, Fauriat, et al. [63] demonstrated that NK cell education can be also mediated by activating KIRs. Their results showed that the expression of KIR2DS1 (an activating KIR) and its ligand (HLA-C2) reduces the responsiveness of NK cells against their targets in both presence and absence of NKG2A [63]. They also found that hyporesponsiveness is limited to target cell recognition because KIR2DS1+ NK cells are stimulated by exogenous cytokines [63].
NK Cell Priming
Resting human NK cells show little cytotoxic functions when incubated in vitro with tumor target cells, suggesting that resting NK cells require additional signals for their complete activation [64]. Resting NK cells should be activated in a process called priming to achieve their effector functions. Several in vitro studies have demonstrated the role of myeloid cells in activation of resting NK cells. In this context, dendritic cells play a critical role in priming resting NK cells through secretion of cytokines as IL-15 [65], IL-18 [66] and IL-27 [67].
Inhibitory/Activating Signals Balance
Under normal circumstances, NK cells can discriminate between normal healthy cells and abnormal cells (infected or transformed) through MHC class I molecules. Healthy cells constitutively express MHC class I molecules which bind to NK cell inhibitory receptor (CD94/NKG2A) to avoid NK cell- mediated lysis (Figure 3A) [68]. On contrary, NK cells are able to recognize and kill their target cells due to the imbalance between inhibitory and activating signals via two models: “missing-self” recognition where the abnormal cells lose the expression of MHC class I ligands to escape the cytotoxic T cells (Figure 3B), and “stress-induced recognition” in which the abnormal cells show an upregulation of damage- associated proteins (Figure 3C).
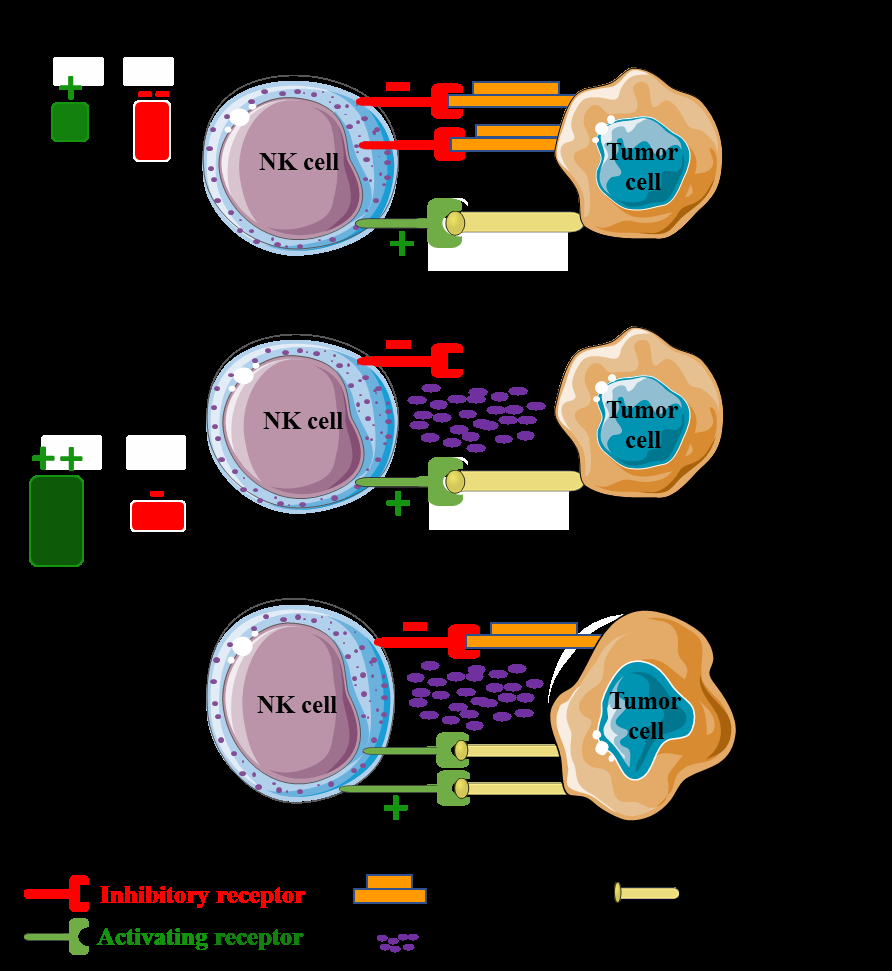
A) NK cells are tolerant to healthy host cells as healthy cells constitutively express MHC class I molecules which bind to NK cell inhibitory receptor to avoid NK cell-mediated lysis. B) Activation of NK cells by missing-self status in which tumor cells lose expression of MHC class I molecules. C) NK cells are activated by stressed cells, upregulating the expression of activating ligands for NK cell activating receptors (stress-induced recognition).
NK Cell Effector Functions
NK cells are key players in the effector arm of the immune system that maintain the homeostasis via recognition and killing abnormal and pathogen-infected cells. Several damage-associated proteins have been found in tumor cells such as MICA, MICB, ULBPs binding with NKG2D, ligands of NKp30 (B7-H6 and HLA- BAT3), mixed-lineage leukemia protein (a ligand of NKp44), and CD155 and CD112 which interact with DNAM-1 [69]. Beside its activation by tumor cells and pathogens, NK cells can directly or indirectly receive signals from other immune cells (Figure 4). For instance, dendritic cells can regulate NK cell proliferation and function by secreting IL-12, type I IFNs, and IL-15 [69]. Monocytes/ macrophages also play a role in NK cell activation and cytotoxicity by secreting cytokines such as IL-2, IL-12, IL- 18, IL-1β and IFN-β. CD4+ T cells can secrete IL-2, which is critical for NK cell survival and proliferation [69].
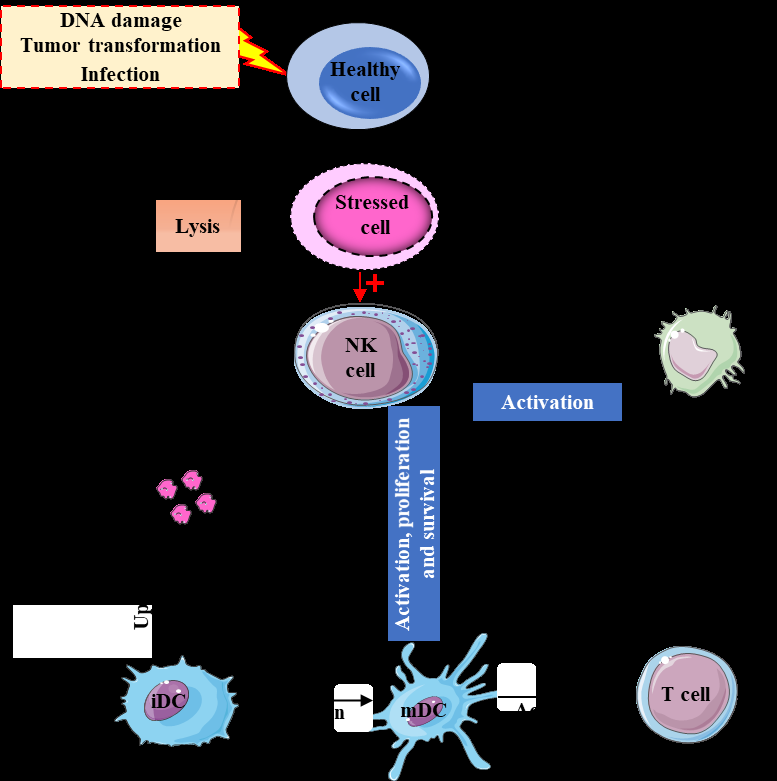
NK cells can recognize and kill stressed cells either directly after their activation by the stressed cells or indirectly by stimulation of other immune cells through cytokine secretion. In addition, other immune cells are able to regulate and activate NK cells. IL: interleukin; IFNγ: interferon-γ; iDC: immature dendritic cell; mDC: mature dendritic cell.
Otherwise, Treg cells can suppress NK cell proliferation and activity by secreting transforming growth factor- β (TGF-β) and IL-10 [69]. Activated NK cells are able to kill their target cells in a process called apoptosis (a process of programmed cell death) though a variety of mechanisms, including cytolytic granule-dependent exocytosis pathway, signaling through the TNF death receptor family members such as FAS (CD95) and TRAIL (TNF-related apoptosis- inducing ligand), ADCC via CD16, and the release of cytokines IFN-γ and TNF-α [70].
NK Cell Cytotoxicity
Cytolytic Granule-Dependent Exocytosis
The cytotoxic granules of NK cells are specific secretory lysosomes contain a mixture of cytotoxic mediators (perforin, granzymes, and granulysin) that cause death of the target cells. Moreover, the membrane of these granules comprise highly glycosylated membrane proteins called lysosomal-associated membrane protein-1 (LAMP-1 or CD107a) and LAMP-2 or CD107b [71]. CD107a expression on NK cell surface has been described as a marker of NK cell degranulation and is upregulated on the cell surface following NK cell [72]. Perforin Perforin is a pore-forming protein (67 kDa), where its gene is constitutively transcribed in NK cells and is regulated by receptor activation signals and cytokines (IL-2 and IL-15). The exact molecular mechanism induced by perforin remains indefinable, but there are two suggested hypotheses describe its function. The first one proposes that perforin makes pores in the membrane of target cell, so this allows granzymes to diffuse into the target cell and also allows an ionic exchange, which causes an osmotic unbalance. The second suggests that perforin binds to the target-cell membrane (through electrostatic or possibly receptor-mediated interactions) and is internalized into its endosomes (containing granzymes) to disrupt them; thereby granzymes are released into the cytosol.
Granzymes
Granzymes are proteases related to serine proteases family. In humans, there are five granzymes (A, B, H, K and M) have been defined where granzymes A and B are the most abundant granzymes [73]. Granzymes are synthesized as pro-enzymes and are activated by cathepsin C and H. Although the most accepted suggestion is that the perforin pores can serve as passive conductors of granzymes through the target cell membrane, Motyka, et al. [74] have shown that in the absence of perforin, granzyme B can form a complex with the mannose-6-phosphate receptor on the surface of target, and then the complex is internalized by endocytosis. Compared with other granzymes, granzyme B has the strongest pro-apoptotic activity. It induces targets death based on caspase-dependent apoptosis rapidly through two pathways. In the first pathway, granzyme B directly activates caspase 3, which enhances DNA destruction [75]. The second is characterized by promotion of permeability of the mitochondrial outer membrane and cleaving of a molecule from the Bcl-2 family called BID (BH3-interacting-domain death agonist). In turn, BID induces cytochrome-C release from mitochondria triggering the activation of caspase 9 which enhances caspase 3 activation [76].
Granulysin
Granulysin is a cationic protein related to the saposin- like protein family which is converted to its active form by proteolytic cleavage from 3 to 5 days after cell activation. It makes pores in the mitochondrial membrane of target cell, disrupting the transmembrane potential (Δψ) in mitochondria [77]. Moreover, granulysin induces cell death through activation of caspase-3 [77].
Death Receptor Pathway
Death receptors are cell surface receptors related to TNF super family that transmit apoptotic signals initiated by specific ligands such as Fas ligand (FasL; CD178), TNF-α and TRAIL (CD253). They play an important role in apoptosis via activation of caspase cascade. Death receptors contain an intracellular death domain (DD), which upon ligand binding associates directly with an adaptor protein called Fas-associated death domain (FADD) or indirectly via TNFR- associated death domain (TRADD). FADD also contains death effector domain which interacts with pro-caspase-8 to form a complex at the receptor called the death inducing signaling complex (DISC). The final step in this process is mobilization of caspase 8 to the DISC causing its activation and initiation of apoptosis [78].
Fas (CD95)
Fas and its ligand (FasL) play an important role in killing targets such as virus-infected cells or cancer cells by NK cells. NK cells express FasL on their surface to suppress tumor growth [79]. In some cases, tumor cells do not express Fas, but NK cells have the ability to induce Fas expression on these cells via IFN-γ secretion [80]. Binding of Fas with FasL promotes receptor clustering, DISC formation and activation of caspase 8 by auto-proteolysis process [81]. Accordingly, caspase 8 converts procaspase 3 to its active form (caspase 3) which initiates apoptosis process via DNA cleavage [81]. Further, caspase 8 can also hydrolyze BID, which destroys the mitochondrial membrane and triggers cytochrome c release [81].
Trail
TRAIL, also known as Apo2 ligand, is a member of TNF superfamily which is capable to induce NK cell-mediated apoptosis for many tumors and transformed cells. Five receptors have been recognized for TRAIL in humans where two of them, TRAIL-R1 (death receptor DR4) and TRAIL-R2 (DR5), are able to initiate an apoptotic signal [82]. Ligation of TRAIL to its receptor (DR4 and DR5) results in engagement of FADD proteins in their cytoplasmic domain, and then formation of DISC. As a result, caspase-8 is activated which is able to trigger the apoptosis process as mentioned above in Fas.
Antibody-Dependent Cell-Mediated Cytotoxicity
Antibody-dependent cell-mediated cytotoxicity (ADCC), also called antibody-dependent cellular cytotoxicity, is an immune mechanism whereby the effector cells bearing Fc receptors can recognize and lyse antibody-coated target cells (opsonized cells). Activating low affinity FcγRIIIA receptor (CD16A) is highly expressed on the cytotoxic CD56dim CD16+ NK cell subset, and mediates ADCC by binding to the Fc portion of IgG antibody. After recognition of their targets, CD56dim CD16+ NK cells interact with the Fc region of IgG antibody coating the target cell which is followed by phosphorylation of ITAMs via cellular src kinases to activate the signaling pathways in NK cells. As a result, cytotoxic granules secrete their contents as perforin and granzymes, causing lysis of target cells [83, 84].
NK Cell Cytokines Secretion
Beside their cytotoxicity, NK cells also have important regulatory function mediated by cytokines and chemokines secretion following their stimulation [85, 86]. Moreover, NK cells participate in a complex interaction network with other lymphocytes, dendritic cells, and macrophages to effectively control immune responses [87]. For instance, NK cells are considered as a major producer of IFN-γ in response to their stimulation, which has immune regulatory activity as well as direct effector activity [88, 89]. Activated NK cells also secrete TNF-α which has both direct cytotoxic activity by triggering caspase-8-mediated apoptosis and immune regulatory activity by inducing dendritic cells maturation [90, 91]. Additionally, NK cells have the ability to secrete other factors as GM-CSF, immunoregulatory cytokines (IL- 5, IL-10, IL-13), and chemokines (MIP-1α, MIP-1β, IL-8, and RANTES) allowing NK cells to attract and co-localize with other immune cells at inflammation sites.
NK Cells and Cancer
NK cells play a major role in the immunosurveillance of malignancy (hematological or solid tumors) by direct killing of malignant cells and/or releasing a number of cytokines that regulate both innate and adaptive immune responses. The cytotoxic activity of NK cell against tumors have been firstly confirmed in animal models, where transplanted hematopoietic tumors or chemically-induced tumors were rejected in various mice models due to NK activity. In contrast, eradicating NK cells in such models often led to an aggressive tumor growth and metastasis [92].
In humans, several studies have shown a relationship between low levels of NK cell activity and the increase in the risk of cancer. For instance, a prospective epidemiological follow-up study has performed in Japan for 11 years and included 3625 individuals showed that a low NK cell cytotoxicity is associated with an increased risk of cancer occurrence [93]. In addition, the infiltration of NK cells into tumors is associated with good prognosis in patients with colorectal carcinoma, gastric carcinoma and lung cancer. Many clinical trials have been used NK cells as adoptive immunotherapy based on the alloreactivity of donor’s NK cells to treat the hematological malignancies and solid tumors [94, 95, 96]. More recently, a variety of ex vivo expansion and activation methods have been used to increase both number and function of the infused NK cells to improve their antitumor activity [97, 98, 99]. However, tumor cells can develop various strategies to escape immunosurveillance of NK cell and other effector cells, which can be explained by general mechanisms such as saturation of the immune system by rapid growth of the tumor as well as alterations of NK cells and other immune cells leading to dysfunction of the immune system [100, 101, 102, 103, 104, 105, 106, 107].
Conclusion
NK cells are distinct lymphocytes playing a prominent role in the innate immune responses against different types of cancer and virus-infected cells. Their effector functions are controlled by activating and inhibitory signals of a wide group of inhibitory and activating receptors expressed on their surface. Phenotypically, NK cells are categorized into two subsets: CD56bright and CD56dim NK cells based on the expression of CD56 molecules, where each subset has characteristic maturation profile and functions. In order to be able to recognize their targets, NK cells must be educated by host MHC class I molecules, and primed via specific cytokines. After that, NK cells can be activated by their target cells as well as signals from other immune cells. Consequently, the target cells are destroyed via the activated NK cells in a process called apoptosis occurred though different mechanisms. Based on their ability to sense the altered expression of MHC molecules and stress markers through various receptors, tumor cells become reachable targets for NK cell lysis due to down-regulation of MHC expression as well as enhancement of stress ligands expression. Many attempts and clinical trials have been previously and still performed to utilize NK cells as immunotherapy to treat solid and hematopoietic tumors (https://clinicaltrials.gov).
Declarations of Interest
None
References
-
Herberman RB, Nunn ME, Lavrin DH (1975) Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic and allogeneic tumors. I. Distribution of reactivity and specificity. Int J Cancer 16(2): 216-229.
-
Kiessling R, Klein E, Wigzell H (1975) “Natural” killer cells in the mouse. I. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotype. Eur J Immunol 5(2): 112-117.
-
Vosshenrich CAJ, García-Ojeda ME, Samson-Villéger SI, Pasqualetto V, Enault L, et al. (2006) A thymic pathway of mouse natural killer cell development characterized by expression of GATA-3 and CD127. Nature Immunology 7(11): 1217-1224.
-
Freud AG, Becknell B, Roychowdhury S, Mao HC, Ferketich AK, et al. (2005) A Human CD34(+) Subset Resides in Lymph Nodes and Differentiates into CD56 bright Natural Killer Cells. Immunity 22(3): 295-304.
-
Moretta L, Bottino C, Pende D, Vitale M, Mingari MC, et al. (2005) Human natural killer cells: Molecular mechanisms controlling NK cell activation and tumor cell lysis. Immunology Letters 100(1): 7-13.
-
Vivier E, Raulet DH, Moretta A, Caligiuri MA, Zitvogel L, et al. (2011) Innate or adaptive immunity? The example of natural killer cells. Science 331(6013): 44-49.
-
Spits H, Artis D, Colonna M, Diefenbach A, Di Santo JP, et al. (2013) Innate lymphoid cells–a proposal for uniform nomenclature. Nature reviews Immunology 13(2): 145.
-
Cooper MA, Fehniger TA, Caligiuri MA (2001) The biology of human natural killer-cell subsets. Trends in Immunology 22 (11): 633-640.
-
Timonen T, Reynolds CW, Ortaldo JR, Herberman RB (1982) Isolation of human and rat natural killer cells. Journal of Immunological Methods 51(3): 269-277.
-
Fehniger TA, Cooper MA, Nuovo GJ, Cella M, Facchetti F, et al. (2003) CD56bright natural killer cells are present in human lymph nodes and are activated by T cell-derived IL-2: a potential new link between adaptive and innate immunity. Blood 101(8): 3052-3057.
-
Vivier E (2006) What is natural in natural killer cells? Immunology Letters 107(1): 1-7.
-
Boos MD, Ramirez K, Kee BL (2008) Extrinsic and intrinsic regulation of early natural killer cell development. Immunologic Research 40(3): 193-207.
-
Kumar Kingsley SM, Vishnu Bhat B (2017) Role of MicroRNAs in the development and function of innate immune cells. International Reviews of Immunology 36(3): 1-22.
-
Boos MD, Yokota Y, Eberl G, Kee BL (2007) Mature natural killer cell and lymphoid tissue–inducing cell development requires Id2-mediated suppression of E protein activity. Journal of Experimental Medicine 204(5): 1119-1130.
-
Male V, Nisoli I, Gascoyne DM, Brady HJM (2012) E4BP4: an unexpected player in the immune response. Trends Immunol 33(2): 98-102.
-
Gascoyne DM, Long E, Veiga-Fernandes H, de Boer J, Williams O, et al. (2009) The basic leucine zipper transcription factor E4BP4 is essential for natural killer cell development. Nat Immunol 10(10): 1118-1124.
-
Kamizono S, Duncan GS, Seidel MG, Morimoto A, Hamada K, et al. (2009) Nfil3/E4bp4 is required for the development and maturation of NK cells in vivo. Journal of Experimental Medicine 206(13): 2977-2986.
-
Male V, Nisoli I, Kostrzewski T, Allan DS, Carlyle JR, et al. (2014) The transcription factor E4bp4/Nfil3 controls commitment to the NK lineage and directly regulates Eomes and Id2 expression. Journal of Experimental Medicine 211(4): 635-642.
-
Knox JJ, Cosma GL, Betts MR, McLane LM (2014) Characterization of T-Bet and Eomes in Peripheral Human Immune Cells. Frontiers in Immunology.
-
Jenne CN, Enders A, Rivera R, Watson SR, Bankovich AJ, et al. (2009) T-bet–dependent S1P 5 expression in NK cells promotes egress from lymph nodes and bone marrow. Journal of Experimental Medicine 206 (11): 2469-2481.
-
Townsend MJ, Weinmann AS, Matsuda JL, Salomon R, Farnham PJ, et al. (2004) T-bet regulates the terminal maturation and homeostasis of NK and Vα14i NKT cells. Immunity 20(4): 477-494.
-
Romee R, Leong JW, Fehniger TA (2014) Utilizing Cytokines to Function-Enable Human NK Cells for the Immunotherapy of Cancer. Scientifica, pp: 1-18.
-
Vosshenrich CAJ, Ranson T, Samson SI, Corcuff E, Colucci F, et al. (2005) Roles for Common Cytokine Receptor γ-Chain-Dependent Cytokines in the Generation, Differentiation, and Maturation of NK Cell Precursors and Peripheral NK Cells in Vivo. Journal of Immunology 174(3): 1213-1221.
-
Dinarello CA, Novick D, Kim S, Kaplanski G (2013) Interleukin-18 and IL-18 Binding Protein. Frontiers in Immunology.
-
Cichocki F, Felices M, McCullar V, Presnell SR, Al-Attar A, et al. (2011) Cutting Edge: MicroRNA-181 Promotes Human NK Cell Development by Regulating Notch Signaling. The Journal of Immunology 187(12): 6171- 6175.
-
Sullivan RP, Leong JW, Schneider SE, Ireland AR, Berrien-Elliott MM, et al. (2015) MicroRNA-15/16 Antagonizes Myb to Control NK Cell Maturation. Journal of Immunology 195(6): 2806-2817.
-
Mundy-Bosse BL, Scoville SD, Chen L, McConnell K, Mao HC, et al. (2016) MicroRNA-29b mediates altered innate immune development in acute leukemia. Journal of Clinical Investigation 126(12): 4404-4416.
-
Lanier LL (2003) Natural killer cell receptor signaling. Current Opinion in Immunology 15(3): 308-314.
-
Pegram HJ, Andrews DM, Smyth MJ, Darcy PK, Kershaw MH, et al. (2011) Activating and inhibitory receptors of natural killer cells. Immunol Cell Biol 89(2): 216-224.
-
Long EO, Sik Kim H, Liu D, Peterson ME, Rajagopalan S, et al. (2013) Controlling Natural Killer Cell Responses: Integration of Signals for Activation and Inhibition. Annual Review of Immunology 31(1): 227-258.
-
Ravetch JV, Lanier LL (2000) Immune inhibitory receptors. Science 290(5489): 84-89.
-
Brooks AG, Posch PE, Scorzelli CJ, Borrego F, Coligan JE (1997) NKG2A complexed with CD94 defines a novel inhibitory natural killer cell receptor. Journal of Experimental Medicine 185(4): 795-800.
-
Carretero M, Palmieri G, Llano M, Tullio V, Santoni A, et al. (1998) Specific engagement of the CD 94/NKG 2-A killer inhibitory receptor by the HLA-E class Ib molecule induces SHP-1 phosphatase recruitment to tyrosine- phosphorylated NKG 2-A: evidence for receptor function in heterologous transfectants. Eur J Immunol 28(4): 1280-1291.
-
Braud VM, Allan DSJ, O’Callaghan CA, Ogg GS, Lazetic S, et al. (1998) HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 391(6669): 795-799.
-
Marsh SGE, Parham P, Dupont B, Geraghty DE, Trowsdale J, et al. (2003) Killer-cell immunoglobulin-like receptor (KIR) nomenclature report, 2002. Immunogenetics 55(4): 220-226.
-
Rajalingam R (2011) Human diversity of killer cell immunoglobulin-like receptors and disease. Korean J Hematol 46(4): 216-218.
-
Rajagopalan S (2010) Endosomal signaling and a novel pathway defined by the natural killer receptor KIR2DL4 (CD158d). Traffic 11(11): 1381-1390.
-
Houchins JP, Yabe T, McSherry C, Bach FH (1991) DNA sequence analysis of NKG2, a family of related cDNA clones encoding type II integral membrane proteins on human natural killer cells. J Exp Med 173(4): 1017-1020.
-
Cosman D, Müllberg J, Sutherland CL, Chin W, Armitage R, et al. (2001) ULBPs, novel MHC class I–related molecules, bind to CMV glycoprotein UL16 and stimulate NK cytotoxicity through the NKG2D receptor. Immunity 14(2): 123-133.
-
Raulet DH, Gasser S, Gowen BG, Deng W, Jung H (2013) Regulation of ligands for the NKG2D activating receptor. Annu Rev Immunol 31: 413-441.
-
Upshaw JL, Arneson LN, Schoon RA, Dick CJ, Billadeau DD, et al. (2006) NKG2D-mediated signaling requires a DAP10-bound Grb2-Vav1 intermediate and phosphatidylinositol-3-kinase in human natural killer cells. Nature Immunology 7(5): 524-532.
-
Pende D, Parolini S, Pessino A, Sivori S, Augugliaro R, et al. (1999) Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J Exp Med 190(10): 1505-1516.
-
Pessino A, Sivori S, Bottino C, Malaspina A, Morelli L, et al. (1998) Molecular cloning of NKp46: a novel member of the immunoglobulin superfamily involved in triggering of natural cytotoxicity. J Exp Med 188(5): 953-960.
-
Vitale M, Bottino C, Sivori S, Sanseverino L, Castriconi R, et al. (1998) NKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non–major histocompatibility complex– restricted tumor cell lysis. J Exp Med 187(12): 2065- 2072.
-
Joyce MG, Sun PD (2011) The Structural Basis of Ligand Recognition by Natural Killer Cell Receptors. J Biomed Biotechnol.
-
Koch J, Steinle A, Watzl C, Mandelboim O (2013) Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 34(4): 182-191.
-
Spaggiari GM, Carosio R, Pende D, Marcenaro S, Rivera P, et al. (2001) NK cell-mediated lysis of autologous antigen-presenting cells is triggered by the engagement of the phosphatidylinositol 3-kinase upon ligation of the natural cytotoxicity receptors NKp30 and NKp46. Eur J Immunol 31(6): 1656-1665.
-
Tassi I, Presti R, Kim S, Yokoyama WM, Gilfillan S, et al. (2005) Phospholipase C- 2 Is a Critical Signaling Mediator for Murine NK Cell Activating Receptors. The Journal of Immunology 175(2): 749-754.
-
Vilches C, Parham P (2002) K IR : Diverse, Rapidly Evolving Receptors of Innate and Adaptive Immunity. Annual Review of Immunology 20(1): 217-251.
-
Stewart CA, Laugier-Anfossi F, Vély F, Saulquin X, Riedmuller J, et al. (2005) Recognition of peptide-MHC class I complexes by activating killer immunoglobulin- like receptors. Proceedings of the National Academy of Sciences of the United States of America 102(37): 13224-13229.
-
Graef T, Moesta AK, Norman PJ, Abi-Rached L, Vago L, et al. (2009) KIR2DS4 is a product of gene conversion with KIR3DL2 that introduced specificity for HLA-A*11 while diminishing avidity for HLA-C. The Journal of Experimental Medicine 206(11): 2557-2572.
-
Nimmerjahn F, Ravetch JV (2007) Fc-receptors as regulators of immunity. Advances in Immunology 96: 179-204.
-
Bottino C, Castriconi R, Pende D, Rivera P, Nanni M, et al. (2003) Identification of PVR (CD155) and Nectin-2 (CD112) as Cell Surface Ligands for the Human DNAM-1 (CD226) Activating Molecule. The Journal of Experimental Medicine 198(4): 557-567.
-
Tahara-Hanaoka S, Shibuya K, Onoda Y, Zhang H, Yamazaki S, et al. (2004) Functional characterization of DNAM-1 (CD226) interaction with its ligands PVR (CD155) and nectin-2 (PRR-2/CD112). Int Immunol 16(4): 533-538.
-
Magri G, Muntasell A, Romo N, Sáez-Borderías A, Pende D, et al. (2011) NKp46 and DNAM-1 NK-cell receptors drive the response to human cytomegalovirus-infected myeloid dendritic cells overcoming viral immune evasion strategies. Blood 117(3): 848-856.
-
Pende D, Spaggiari GM, Marcenaro S, Martini S, Rivera P, et al. (2005) Analysis of the receptor-ligand interactions in the natural killer-mediated lysis of freshly isolated myeloid or lymphoblastic leukemias: evidence for the involvement of the Poliovirus receptor (CD155) and Nectin-2 (CD112). Blood 105(5): 2066-2073.
-
Pende D, Castriconi R, Romagnani P, Spaggiari GM, Marcenaro S, et al. (2006) Expression of the DNAM- 1 ligands, Nectin-2 (CD112) and poliovirus receptor (CD155), on dendritic cells: relevance for natural killer- dendritic cell interaction. Blood 107(5): 2030-2036.
-
Smyth MJ, Cretney E, Kelly JM, Westwood JA, Street SE, et al. (2005) Activation of NK cell cytotoxicity. Molecular Immunology 42(4): 501-510.
-
Assarsson E, Kambayashi T, Persson CM, Ljunggren HG, Chambers BJ (2005) 2B4 co-stimulation: NK cells and their control of adaptive immune responses. Molecular Immunology 42(4): 419-423.
-
Zingoni A, Sornasse T, Cocks BG, Tanaka Y, Santoni A, et al. (2005) NK cell regulation of T cell-mediated responses. Molecular Immunology 42(4): 451-454.
-
Anfossi N, André P, Guia S, Falk CS, Roetynck S, et al. (2006) Human NK Cell Education by Inhibitory Receptors for MHC Class I. Immunity 25(2): 331-342.
-
Kim S, Poursine-Laurent J, Truscott SM, Lybarger L, Song YJ, et al. (2005) Licensing of natural killer cells by host major histocompatibility complex class I molecules. Nature 436(7051): 709-713.
-
Fauriat C, Ivarsson MA, Ljunggren HG, Malmberg KJ, Michaëlsson J (2010) Education of human natural killer cells by activating killer cell immunoglobulin-like receptors. Blood 115(6): 1166-1174.
-
Bryceson YT, March ME, Ljunggren HG, Long EO (2006) Synergy among receptors on resting NK cells for the activation of natural cytotoxicity and cytokine secretion. Blood 107(1): 159-166.
-
Lucas M, Schachterle W, Oberle K, Aichele P, Diefenbach A, et al. (2007) Dendritic Cells Prime Natural Killer Cells by trans-Presenting Interleukin 15. Immunity 26(4): 503-517.
-
Chaix J, Tessmer MS, Hoebe K, Fuséri N, Ryffel B, et al. (2008) Cutting Edge: Priming of NK Cells by IL-18. The Journal of Immunology 181(3): 1627-1631.
-
Ziblat A, Domaica CI, Spallanzani RG, Iraolagoitia XL, Rossi LE, et al. (2015) IL-27 stimulates human NK- cell effector functions and primes NK cells for IL-18 responsiveness: Immunomodulation. Eur J Immunol 45(1): 192-202.
-
Moretta L, Moretta A (2004) Unravelling natural killer cell function: triggering and inhibitory human NK receptors. EMBO 23(2): 255-259.
-
Fang F, Xiao W, Tian Z (2017) NK cell-based immunotherapy for cancer. Seminars in Immunology 31 37-54.
-
Chavez-Galan L, Arenas-Del Angel MC, Zenteno E, Chávez R, Lascurain R, et al. (2009) Cell death mechanisms induced by cytotoxic lymphocytes. Cellular & molecular immunology 6(1): 15-25.
-
Winchester BG (2001) Lysosomal membrane proteins. European Journal of Paediatric Neurology 5: 11-19.
-
Alter G, Malenfant JM, Altfeld M (2004) CD107a as a functional marker for the identification of natural killer cell activity. Journal of Immunological Methods 294(1– 2): 15-22.
-
Grossman WJ, Revell PA, Lu ZH, Johnson H, Bredemeyer AJ,et al. (2003) The orphan granzymes of humans and mice. Current Opinion in Immunology 15(5): 544-552.
-
Motyka B, Korbutt G, Pinkoski MJ, Heibein JA, Caputo A, et al. (2000) Mannose 6-phosphate/insulin-like growth factor II receptor is a death receptor for granzyme B during cytotoxic T cell-induced apoptosis. Cell 103(3): 491-500.
-
Metkar SS, Wang B, Ebbs ML, Kim JH, Lee YJ, et al. (2003) Granzyme B activates procaspase-3 which signals a mitochondrial amplification loop for maximal apoptosis. J Cell Biol 160(6): 875-885.
-
Pinkoski MJ, Waterhouse NJ, Heibein JA, Wolf BB, Kuwana T, et al. (2001) Granzyme B-mediated Apoptosis Proceeds Predominantly through a Bcl-2-inhibitable Mitochondrial Pathway. Journal of Biological Chemistry 276(15): 12060-12067.
-
Kaspar AA, Okada S, Kumar J, Poulain FR, Drouvalakis KA, et al. (2001) A Distinct Pathway of Cell-Mediated Apoptosis Initiated by Granulysin. The J Immunol 167(1): 350-356.
-
Thorburn A (2004) Death receptor-induced cell killing. Cell Signal 16(2): 139-144.
-
Bradley M, Zeytun A, Rafi-Janajreh A, Nagarkatti PS, Nagarkatti M (1998) Role of spontaneous and interleukin-2-induced natural killer cell activity in the cytotoxicity and rejection of Fas+ and Fas- tumor cells. Blood 92 (11): 4248-4255.
-
Screpanti V, Wallin RP, Ljunggren HG, Grandien A (2001) A central role for death receptor-mediated apoptosis in the rejection of tumors by NK cells. J Immunol 167(4): 2068-2073.
-
Carrington PE, Sandu C, Wei Y, Hill JM, Morisawa G, et al. (2006) The structure of FADD and its mode of interaction with procaspase-8. Molecular Cell 22(5): 599-610.
-
Wang S, El-Deiry WS (2003) TRAIL and apoptosis induction by TNF-family death receptors. Oncogene 22(53): 8628-8633.
-
Seidel UJE, Schlegel P, Lang P (2013) Natural Killer Cell Mediated Antibody-Dependent Cellular Cytotoxicity in Tumor Immunotherapy with Therapeutic Antibodies. Front Immunol 4: 76.
-
Wang W, Erbe AK, Hank JA, Morris ZS, Sondel PM (2015) NK cell-mediated antibody-dependent cellular cytotoxicity in cancer immunotherapy. Front Immunol 6: 368.
-
Fauriat C, Long EO, Ljunggren HG, Bryceson YT (2010) Regulation of human NK-cell cytokine and chemokine production by target cell recognition. Blood 115(11): 2167-2176.
-
Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S (2008) Functions of natural killer cells. Nature Immunology 9(5): 503-510.
-
Newman KC, Riley EM (2007) Whatever turns you on: accessory-cell-dependent activation of NK cells by pathogens. Nat Rev Immunol 7(4): 279-291.
-
Martin-Fontecha A, Thomsen LL, Brett S, Gerard C, Lipp M, et al. (2004) Induced recruitment of NK cells to lymph nodes provides IFN-γ for TH1 priming. Nat Immunol 5(12): 1260-1265.
-
Wang R, Jaw JJ, Stutzman NC, Zou Z, Sun PD (2012) Natural killer cell-produced IFN-γ and TNF-α induce target cell cytolysis through up-regulation of ICAM-1. J Leukocyte Biology 91(2): 299-309.
-
Gerosa F, Baldani-Guerra B, Nisii C, Marchesini V, Carra G, et al. (2002) Reciprocal Activating Interaction between Natural Killer Cells and Dendritic Cells. J Exp Med 195(3): 327-333.
-
Wang L, Du F, Wang X (2008) TNF-α Induces Two Distinct Caspase-8 Activation Pathways. Cell 133(4): 693-703.
-
Waldhauer I, Steinle A (2008) NK cells and cancer immunosurveillance. Oncogene 27(45): 5932-5943.
-
Imai K, Matsuyama S, Miyake S, Suga K, Nakachi K (2000) Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: an 11-year follow-up study of a general population. Lancet 356(9244): 1795- 1799.
-
Geller MA, Cooley S, Judson PL, Ghebre R, Carson LF, et al. (2011) A phase II study of allogeneic natural killer cell therapy to treat patients with recurrent ovarian and breast cancer. Cytotherapy 13(1): 98-107.
-
Miller JS, Soignier Y, Panoskaltsis-Mortari A, McNearney SA, Yun GH, et al. (2005) Successful adoptive transfer and in vivo expansion of human haploidentical NK cells in patients with cancer. Blood 105(8): 3051-3057.
-
Ruggeri L, Mancusi A, Burchielli E, Aversa F, Martelli MF, et al. (2007) Natural killer cell alloreactivity in allogeneic hematopoietic transplantation. Curr Opinion oncol 19(2): 142-147.
-
Denman CJ, Senyukov VV, Somanchi SS, Phatarpekar PV, Kopp LM, et al. (2012) Membrane-Bound IL-21 Promotes Sustained Ex Vivo Proliferation of Human Natural Killer Cells. PLoS ONE 7(1): 30264.
-
Fujisaki H, Kakuda H, Shimasaki N, Imai C, Ma J, et al. (2009) Expansion of Highly Cytotoxic Human Natural Killer Cells for Cancer Cell Therapy. Cancer Research 69(9): 4010-4017.
-
Sutlu T, Stellan B, Gilljam M, Quezada HC, Nahi H, et al. (2010) Clinical-grade, large-scale, feeder-free expansion of highly active human natural killer cells for adoptive immunotherapy using an automated bioreactor. Cytotherapy 12(8): 1044-1055.
-
Chretien AS, Fauriat C, Orlanducci F, Galseran C, Rey J, et al. (2017) Natural Killer Defective Maturation is Associated with Adverse Clinical Outcome in Patients with Acute Myeloid Leukemia. Frontiers in Immunology 8: 573.
-
Chretien AS, Devillier R, Fauriat C, Orlanducci F, Harbi S, et al. (2017) NKp46 expression on NK cells as a prognostic and predictive biomarker for response to allo-SCT in patients with AML. Onco Immunology 6(12): 1307491.
-
Costello RT, Sivori S, Marcenaro E, Lafage-Pochitaloff M, Mozziconacci MJ, et al. (2002) Defective expression and function of natural killer cell–triggering receptors in patients with acute myeloid leukemia. Blood 99(10): 3661-3667.
-
Diermayr S, Himmelreich H, Durovic B, Mathys- Schneeberger A, Siegler U, et al. (2008) NKG2D ligand expression in AML increases in response to HDAC inhibitor valproic acid and contributes to allorecognition by NK-cell lines with single KIR-HLA class I specificities. Blood 111(3): 1428-1436.
-
Fauriat C, Just-Landi S, Mallet F, Arnoulet C, Sainty D, et al. (2007) Deficient expression of NCR in NK cells from acute myeloid leukemia: evolution during leukemia treatment and impact of leukemia cells in NCRdull phenotype induction. Blood 109(1): 323-330.
-
Khaznadar Z, Boissel N, Agaugué S, Henry G, Cheok M, et al. (2015) Defective NK Cells in Acute Myeloid Leukemia Patients at Diagnosis Are Associated with Blast Transcriptional Signatures of Immune Evasion. The J Immunol 195(6): 2580-2590.
-
Sanchez-Correa B, Morgado S, Gayoso I, Bergua JM, Casado JG, et al. (2011) Human NK cells in acute myeloid leukaemia patients: analysis of NK cell-activating receptors and their ligands. Cancer Immunology, Immunotherapy 60(8): 1195-1205.
-
Stringaris K, Sekine T, Khoder A, Alsuliman A, Razzaghi B, et al. (2014) Leukemia-induced phenotypic and functional defects in natural killer cells predict failure to achieve remission in acute myeloid leukemia. Haematologica 99(5): 836-847.
- Acido Labile or Gastro Irritant Apis and Enteric Release in Galenic Practice: An Overview
- A Study on Knowledge, Attitude and Practice of Hand Hygiene among Healthcare Professionals at a Tertiary Care Hospital, India
- Influence of Inoculum Concentration on In Vivo Incubation Period of Emmia lacerata, Pathogenesis and Management of Wilt in Pepper (Capsicum annuum L.)
- Vanilla’s Chemistry
- Marine Anti-Cancer Compounds and Adverse Effects of Global Warming on Oceans: An Overview
- Serological Investigation of Chikungunya Virus Antibody among Malaria-Suspected Febrile Patients in Some Healthcare Facilities in Rivers State