Some Aspects of Interactions between Nanomaterials and the Cytoskeleton of Eukaryotic cells
<p>All cellular functions are primarily related to the behavior of the components of cell and its cytoskeleton. Cells in the various tissues respond to the sudden changes in their ambient environment may it be biochemical, physiological, molecular or physicochemical in nature. Cells become the ultimate target of these changes and face their derogative and stressful impacts. Cytoskeleton is primarily concerned with the temporal and spatial organizations in both prokaryotes and eukaryotes; these should be in the form of integrated, coordinated functional capabilities in order to perform their designated functions amicably. The functional aspects of cytoskeleton play essential roles in the deliverance of cellular responses under normal circumstances and also with respect to the changed and stressful stimuli. In these regards the components of cytoskeleton have to put an effort to deal with changed conditions. Cell membrane and cell organelles exhibit responses; these are related to cellular elasticity, mobility, adhesion, invasion, proliferation and differentiation, etc. Nanotechnology enables a series of nonmaterial’s to be synthesized/engineered with precise control over their physicochemical properties, surface modifications with respect to targeted moieties, charge modification, stealth aspects which may be ambiguous but features like size, shape, or other characters can be screened in accordance to the appropriate properties of target-specifications. The impacts of nonmaterial’s on biotic components are related to these features and may inflict biochemical, physiological and physic-physiological adverse effects on the eukaryotic cells. Fairly good amount of references are available on the interactions between nonmaterial’s and cells and cell organelles while there appears to be relatively less references pertaining to the interactions between nonmaterial’s and cytoskeleton; therefore, it was thought imperative to evaluate the impressions of nonmaterial’s on cytoskeleton of eukaryotic cells. In this review, overall impacts of nonmaterial’s on the structural and functional components of the cytoskeleton have been evaluated.</p>
Introduction
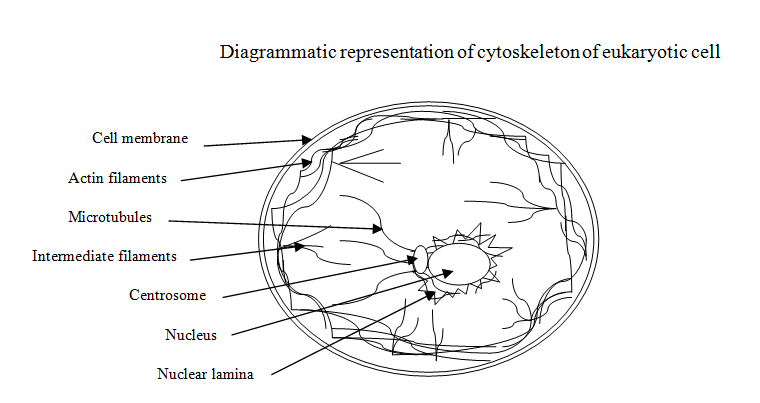
Eukaryotic Cytoskeleton-An Over View
Cell is a structural and functional unit of a biosystem, may it be archaic, prokaryotes or eukaryotes. There is a specific need for cells to have integrated, coordinated functional capabilities to accomplish assigned functions. Cells in a multi-cellular biosystems are surrounded by extracellular matrix; that provides supportive and functional assistance for their successful normal spatial, structural and functional efficacies, [1]. Quite often cells undergo modifications that enable them to perform functions assigned to them. Cytoskeleton is basically concerned with effective cellular organization in a given morphological, histological and functional niche. It provides ability to interact with the concerned environment, to possess correct and suitable shape so that functional fluctuations are appropriately accommodated to maintain the cell in physic- physiologically ‘robust’ conditions and for this the internal structures of the cell should be well structured. Cytoskeleton constitutes the internal frame work of a cell that is responsible for the distinct cellular shape, size and highly elaborated intracellular organization [2]. It is an array of wiry and tubular structures that is highly dynamic matrix; this meets a wide range of functional aspects of cellular entities. Its dynamism is well elaborated in the working of cell and its components like cilia, flagella, pseudopodia along with intracellular processes such as movements of chromosomes during cell division, muscular contraction and relaxation, intracellular positioning and moving of cell organelles within [2, 3]. suggested that during such intra and inter- cellular processes there is an involvement of tyrosine kinas, lipid kinas, phospholipids, serine/heroine kinas etc; for example, poisonings trigger the process of phagocytosis [4]. Fc receptors and other signaling pathways have been identified but the exploitation of dynamic nature of cytoskeleton is among the prime factors involved in these processes; this is well illustrated in the process of phagocytosis in case of macrophages and other cells. The activating signaling molecules induce number of molecules that participate in the remodeling of the components of cytoskeleton. Some of the remodeling molecules are GTPase of Rho family, guanine-nucleotide exchange factors related to control GTPase and the factors that promote nucleation of actin, the actin-nucleation complex Arp2/3 and molecular motors of myosin family. All these factors focus on the synchronized and chronological functioning of cytoskeleton to smoothen cellular functionalities. Cytoskeleton is present in the form of interwoven mesh-work filling the space between nucleus and the cell membrane. It is a dynamic component of cell and is responsible for the cellular size, shape, cell division, growth and most of the other cellular functions. Its configuration maintains cellular temporal and spatial organization [5]. The degree of elaboration of cellular functions varies in prokaryotes and eukaryotes and these are well illustrated by the performance of the cells in their specific environments. All cellular functions are primarily based on the behavior of components of the eukaryotic cytoskeleton that are grouped in to three types namely: intermediate filaments, microtubules, actin filaments (also referred as microfilaments). Intermediate filaments are responsible for providing mechanical strength and resistance to sheer/stress experienced by the cell. Microtubules are dedicated to determine the position of membrane enclosed organelles and the intra cellular transport involved therein. Actin filaments are primarily concerned with the determination of shape of the surface of cell and play an essential role in cellular locomotion partly and/or as one structural and functional unit. Normally, these cytoskeleton filaments are not effective on their own; their efficacy depends on the ‘accessory proteins’ that are linked with each filament and also with cellular components. Accessory proteins are responsible for the controlled, coordinated assembly of filaments of cytoskeleton with respect to specific cellular location. Further, the effectiveness of overall functioning of the filaments of cytoskeleton and accessory proteins is enhanced by “motor proteins”. Cytoskeleton is essential in regulating protein-protein interaction specifically the signal path ways, its study may help in understanding the legend dependant receptor signaling; this feature is very important in case of survival of B-cells [6].
Microfilaments
Commonly these are also called actin filaments. Each filament is a polymer of sub-unit of actin protein, a globular protein. This protein plays an essential role in almost all cellular functions have compared the atomic structures of G-actin and F-actin proteins and found that one molecule of the two parallel F-actin protein rotates through 166˚ then these perfectly come on top of each other exactly resulting into a double helical structure of microfilaments mostly found in the cytoskeleton [7, 8]. Transportation and apportionment of most of the protein complexes is controlled by the dynamics of actin protein and it is related to polymerization and depolymerization [9]. The F-actin-ADP was found to be present in higher concentration on negative end of the filament and this end represented the ‘shrinking end” of the actin filament; the barbed ends of the actin filament are placed towards the cellular membrane while pointed ends were observed to be towards the interior of the cell. Actin can be grouped in to muscular-specific-actin and non-muscular-specific- actin; muscular specific actin is referred as α-actin while non-muscular-specific-actin as β and γ- actin; β-actin is located at the epical region of epithelial cells and γ-actin at the lateral and basal regions of epithelial cells [2]. Vidin & Gunning [10] have identified three main groups of actin isomeric namely, α, β and γ forms among vertebrates. Of these three isomeric forms α-actin constitutes the prime component of contractile aspects of muscular tissue while β and γ- isomeric forms are present in other types of cells as components of their cytoskeleton and are responsible for internal cellular motility; tropomyosins plays a prime role in the formation of wide range of cell structures along with actin so that most of the cellular functions are ensured [10]. Cell signaling plays an essential role in regulating the assembly of the actin in the cell, [11]. Microfilaments/actin protein accomplishes varieties of cellular functions in collaboration with specific group of proteins; these proteins bring about the binding between embedded protein in the cell membrane and microfilaments while proteins like spectrum and ankyrin that are present in erythrocytes and epithelial cells also help microfilaments to carry out normal cellular functions, [2]. The growth of these filaments forces the membrane to thrust outwards. Forman’s proteins facilitate the lengthening of unbranched filament that assist in fast growing ends of the filaments and also prevent them from being capped. Hui-Chiai & William [12] found that ‘Collapsing Response Mediator Protein - 1(CRMP-1)’ acts as an auxiliary cofactor and responsible for demodulating cell motility in response to extracellular signal [12]. Gunning [13] are of the opinion that actin filaments have exhibited phylogenetic hierarchy at various levels of biological taxonomic domains; these filaments have shown vast diversification in tropomyosins and have attained compositional and structural variation and this has been the bases of diversity in their function [13]. Microtubules Microtubules along with microfilaments and intermediate filaments are responsible for the cell structure; these form the internal structures of cilia and flagella. Vale suggested the role of the microtubules in the form of basic foundation for the movements of secretary vesicles, organelles, macromolecular assemblies within cell, separation of chromosomes during meiosis and mitotic spindles [14]. Microtubules exhibit the phenomenon of nucleation – this process is regulated by ‘Microtubules Organization Center’, centrosome, basal bodies and participates in the formation of mitotic and meiotic spindles; cytoplasm microtubules are concerned with the fixing of the position of nucleus and spindle. Bartolini & Gunderson [15] proposed that the organization of microtubules differs in different types of cell, these are related with the transport of protein vesicles and organelles; in fibroblasts and mesenchymal type of cells, the microtubules are located at the centrosome and radiate with their positive poles towards the exterior or peripheral zone, functionalizing the cellular migration [15]. The polarity of microtubules along with motor proteins facilitates the intracellular organization. The protofilaments exhibit uniform orientation of tubule that results in the formation of two polar region these are structurally and chemically different having inherent polarity [2]. Dynamics of microtubules is dependent on the stochastic balance between polymerization and severing actions; these actions can cause differential spatiotemporal abundance and also distribution of microtubules when cells are undergoing developmental process, differentiation and morphogenesis [16]. Further, when catena and related proteins interact with microtubules during their dynamic- interactions some changes do take place in the structure of microtubules affecting their architect and this technique is suitably exploited to investigate the structural changes in microtubules during cell division, migration and maintaining shape of the cells. Environmental factors, different regulators of microtubules and the modifications in microtubules, all these play an essential role in controlling the dynamics and organization of microtubules through the cell cycle and developmental stages [17].
Tubular proteins are the target for tubule binding chemotherapeutics; these drugs arrest the mitosis by reducing the dynamics of mitotic spindle resulting in the death of the cell. Fluctuations in the stability of isotopes of tubule and changes after translation, cause bad/wrong prediction and also predict the resistance developed towards the chemicals used and specifically in case of hematological cancers. These observations definitely indicate that tubule along with ‘microtubules associated proteins’ reflect wide range of stress that is induced in the cells on the onset of cancer [18]. Some of the drugs affect the functioning of microtubules in varied manner. Colchicine, colcemid (demecolcine) and nocodazole inhibit polymerization by binding to tubule and these prevent the addition of subunits to the positive end. Vinbalstine and vincristine (leurocristine) both are used as chemotherapeutic cancer drugs, aggregate tubulin and lead to depolymerization of microtubule. Texel (generic name paditaxel), paclitaxel- (an anticancer drug), a cytotoxic, euplastic and chemotherapeutic in nature, stabilizes microtubules by binding them to a polymer [2].
Intermediate Filaments
This component of the cytoskeleton includes six classes such as acid and basic cytokeratins, valentine, deskman, GFA proteins, neurofilaments, nuclear lamina and nesting (neuroectodermal stem cell protein); these intermediate filament forms are tissue specific in nature. Hermann, et al. [19] suggested that intermediate filaments are the prime structural component of the cell concerned with mechanical stress absorber and are present in the space between nucleus and the cell membrane [19]. The assembly of intermediate filaments involves dimmer, tetramers, protofilaments; these all get associated in order to accomplish the cellular functions [2]. Kreplok & Fudge [20] have described these filaments to be flexible, extensible and are responsible for the “mechanotransduction” and dedicated to mechanical aspects of tissue [20]. Lee, et al. [21] observed that intermediate filaments possess middle domain having α- helical rod made of four α-helical segments 1A, 1B, 2A and 2B, interspaced with linkers [21].
Interaction between Nanomaterials and Eukaryotic Cytoskeleton
Nanotechnology enables a series of nonmaterial’s to be synthesized with precise control over the surface modifications like targeting moieties, charge modification, stealth aspects which are either secret or so far unknown, size, shape, or other particle characters; these characters can be screened in accordance to appropriate properties of target-specification. The nonmaterial’s either natural or man-made exhibit some of the specific properties such as design, flexibility, small size, large surface to volume ratio, ability to move across the biological barriers, ease of surface modification with multiple legend to enhance the avidity for the respective target molecules, cellular components, etc. Most of these features make the nonmaterials suitable for their applications in varied forms to biological systems. Nan materials like, carbon based nonmaterial’s, metals, metal oxide and semiconductors, cell membrane and cytoskeleton exhibit derogative changes related to cellular elasticity, mobility, adhesion and invasion; Wu, et al. [22] have elaborated on the biophysical responses on the interaction of nonmaterial’s with cellular interfaces [22]. Cell membrane is among the many biological fronts that encounters the interaction with nonmaterial’s and it is envisaged that cytoskeleton also included in the process as it plays major role in cell behavior [23, 24]. Interactions at the interface between cell membrane and nonmaterial’s should be well studied in order to understand the cell behavior under normal, pathological and other abnormal conditions caused due to the interaction with toxicants, pharmaceutical drugs and other additives used in food but there seems to be a greater need to investigate the impact of nonmaterial’s on the cytoskeleton which plays prime role in the behavior of cell membrane. Berry, et al. [25] found that SPION (supra paramagnetic iron oxide nanoparticles) induced stress in dermal fibroblast cell, these induced particles derived from transferring to get accumulated near cell membrane deceiving receptor dependent endocytosis, cell signaling and cytoskeleton [25]. Lee, et al. [26] reported that SiO2 (5 nm to 150 nm) caused stress on the process of actin filaments formation [26]. Apopka, et al. [27] treated micro vascular endothelial cells with uncoated magnetite NPs and these get internalized (50µg/ml, 10 min to 5 h) as a result permeability of the cells was found to be increased, cortical F-actin filaments in these cells exhibited defective polymerization [27]. The effect of iron nanoparticles (38 nm ± 8.14, 0.1mM, 1, 6 and 24 h) was observed to be dose dependent and resulted in unpredictable viability of human umbilical vein endothelial cells, internalization by endocytosis and induction of apoptosis; these nanoparticles disorganized actin fibers and net work of β- tubule of the cytoskeleton,[28]. Tiwari, et al. [29] mentioned that iron oxide nanoparticles have the ability to alter cytoskeleton, disrupt the cell-signaling, cause stress path ways, produce reactive oxygen species and elevate oxygen stress. Further, these nanoparticles can bring about leakage from lissome, changes in cell cycle and impairment in mitochondria [29]. Soenen, et al. [30] observed that magnetolioposomes (MLs) brought concentration dependant changes in the architecture of actin cytoskeleton affecting focal adhesion and cell division [30]. Actin cytoskeleton was adversely affected when high doses of cationic amphophilic iron oxide nanoparticles were internalized in the marine C 17.2 neural progenitor cell, blood endothelial cells exhibited declined rate of proliferation, defective focal adhesion in the cell membranes were noticed; declined pronuclear accumulation of iron oxide as magnetolioposomes (MLs) reduced the efficiency of protein expression and satirical maturity of actin fibers. These resulted in improper cell migration and cell differentiation; non-toxic doses induced varied degrees of cell stress [30]. Singh, et al. [31] suggested that SPION (uncoated 100µg/ml – safe and non-toxic; coated with bipolar surfactant 0.1 – 10.0µg/ml toxic) with and/or without coating caused changed in the actin cytoskeleton as a result iron homeostasis was disturbed in the cell studied and the affected cells responded abnormally to the stimuli for gene expression and cell regulation [31]. Silicon nanocrystals internalized without causing any damage to cytoskeleton but high doses could affect actin component of cytoskeleton (0.75µg/ml, 7.5µg/ml and 75µg/ml for 24 to 48h; 2.5 to 3.0 nm and aggregates up to 150nm); nanodimonds when internalized got accumulated at the pronuclear region, higher concentration of nanodimonds disorganized the arrangement of actin filaments and higher dose resulted in apoptosis and necrosis respectively [32]. Vijaykumar, et al. [33] have observed that heterogeneous multi- Titanium oxide nanostructures declined the degree of adherence of the He La cells and fibroblast cells; when the cells were subjected to anatase-dominent Ti nanostructures the well defined amoeba cells were predominant while in rutile-dominent Ti nanostructures needle shaped cells were found to be more (incubation time 24 and 48 h); this indicated that these cells have ability to sense their ambient environment and give response by transmitting extracellular signals to the nucleus [33]. The nanostructures also affected the rate of cell division, polymerization of actin filaments in these cells, changed the chemistry of cytoskeleton; these changes affected the degree of cell adhesiveness, change in shape of the He La (a human cervical cancer and N1H3T3- fibroblast cells) [33]. Mironava, et al. [34] observed that silver nanoparticles can enter the dermal fibroblast cells after penetrating plasma membrane and get accumulated in vacuoles. They observed that uptake of silver nanoparticles was dependent on duration of exposure, size and concentration. Silver nanoparticles having 13 nm sizes were phagocytosed and 45 nm sized silver nanoparticles were endocytosed via clattering mediation. Silver nanoparticles when interacted with dermal fibroblasts could disrupt the filaments of cytoskeleton; when these nanoparticles were removed these fibers could get rearranged to their original form but β-tubule fibers were not affected. This indicated that silver nanoparticles resulted in contraction of the area and declined the mobility [34]. The damage caused to dermal fibroblast was seen as temporary effect because on removal of silver nanoparticles the cellular functions were restored. Xu, et al. [35] found that silver nanoparticles having 20 nm diameters and with concentrations 1, 5, 10 and 50 μg/ml affected primary rat cortical cell cultures; 1 to 5 μg/ml concentration of silver nanoparticles declined permeability of premature neurons and glial cells (neuroglial cells) and prevented the neural growth [35]. Silver nanoparticles initiated the degeneration of matured neurons and also the components of cytoskeleton such as β-tubulin, filamentous actin (F-actin) were gradually lost. Further, silver nanoparticles reduced the number of synaptic clusters by reducing synaptophysin (also called major synaptic vesicle protein p38) and post synaptic density protein. When macrophages of lungs were exposed to 20 nm and 110 nm sized Ag NPs the responses were based on measurement of cellular mechanism like zero to low up take of Ag NPs (nanoparticles) did not change the mechanism of the cells. When 0.5mg/kg (body weight) of Ag NPs having 20nm and 110nm were internalized they induced rearrangement of cytoskeleton causing stiffness in the cells and later the stiffness became very soft indicating damage of the cytoskeleton [36]. Cooper & Spitzer [37] observed that when neural stem cells were exposed to the concentration of 1.0 μg/ml of silver nanoparticles the formation of F-actin inclusion was initiated thereby the action of ‘actin’ was disturbed or declined, silver nanoparticles were not sequestered [37]. Sub-lethal concentration of silver nanoparticles disturbed the dynamics of actin in sub-ventricular neural stem cells resulting in the disruption of neurogenic process and this may be considered that silver nanoparticles are ineffective in case of brain function. Silver nanoparticles induced oxidative stress, genotoxicity to hemolytic and gill cells in case of Mussel and resulted in enhanced liposomal Ac P activity, adversely affected actin of cytoskeleton and Na/K ATPase activity, further, when the severity was compared between silver nanoparticles, bulk and ionic forms the ionic forms were found to be more destructive, Kutsumiti, et al. [38] & Boehlert, et al. [39] reported that when silver nanoparticles were coated with surfactant possibly affected ‘out-side in signaling’, causing oxidative stress, disrupted canonical pathway related to inter linked kinas, extracellular signal regulated kinas and mutagen activated protein kinas, integrands , adherence and the intercellular tight junctions, further, transcription factors that mediated cellular reactions with respect to oxidative stress and caused changes in cytoskeleton along with cell-cell and cell to matrix links [39]. Carbon nonmaterial’s have their significant role in the many of the scientific and industrial fields and come in contact with bios stems, carbon annotates, fullerenes, carbon nanoparticles along with their derivatives etc, are some of the common nanomaterials which are being exploited in medical, research and development and industrial applications. Pamplona & Florin [40] have proposed that carbon annotates are very much similar to microtubules; the parameter of these nonmaterial’s with respect to stiffness value (Young’s modulus is ~1TPa) and it is around five times more than steel (~210GPa) [40]. Although, microtubules and carbon annotates exhibit similar mechanical behaviors as elastic module but both have different chemical compositions. Microtubules have very flexible adaptability with respect to dynamism and this is reflected during their assembly (growth) and disassembly (shrinkage) phase. Under normal conditions globular actin plays an essential role in forming a net work that assists the maintenance of the cellular shape and also help in cellular mobility and cell division; the network functions in generation of force to accomplish the functional aspects. Further, when cells were treated with purified SWCNTs dispersion the actions in these cells showed bundling of its fibers; this feature reduced cell division and elevated actin related defective cell division and induced chronic cellular functionalities [41]. Ying, et al. [42] have suggested that carbon nonmaterials like fullerenes and their derivatives, carbon annotates, grapheme etc, influence the actin component of the cytoskeleton in three basic pathways: interaction with polymerization of actin, extracellular signaling pathway that initiate the interaction and production of reactive species of oxygen [42]. Boyer, et al. [43] have observed that SWCNTs were found to be attached to the tail domain of the lamin-1 (B-1) while the carbon nonmaterials were delivered to nucleus; more than 50% of globular protein was disorganized but retained its central hydrophobic core. This core was suitably bounded to SWCNTs while lamin-1 activated the import of SWCNTs to nucleus [43]. Apoptosis is one of the natural processes devoted to cellular homeostasis and to eliminate unwanted, defunctionalized cells from the bios stem, Lahir [44] the role of cytoskeleton in apoptosis still needs elaboration but Youle & Karbowski [45] observed that fullerene facilitated fragmentation of microtubules and also of the membrane of mitochondria [45]. Johnston, et al. [46] proposed that molecular dynamics is very essential to investigate the movement or interaction of fullerene with cell membrane. They found that untreated and pure fullerene can easily move across the lipid membrane involving the ‘transient micropore’ or ‘jump’ through the membrane while C-60(OH)20 could not enter through the membrane possibly because of their hydrophilic nature. Thus, pristine fullerene and treated/derivatized forms of fullerene while interactin with cell membrane exhibit differences in their toxic impact. Further, they concluded that the potential of fullerene should be evaluated in the oxidative dependent responses, inflammatory and genotoxical aspects [46]. Ratnikoval, et al. [47] reported that fullerene derivative C-60(OH)20 in micro-molar concentration did not enhance the polymerization of microtubules, this inhibition of polymerization was accomplished by the hydrogen bonding involving fullerene nanomaterials and heterodimers of tubulin and inhibited the release of inorganic phosphate [47].The fullerene derivative was found to be bound to tubulin at molecular ratio 9:1 as observed by isothermal calorimetric technique; these observations were based on docking and molecular dynamic simulation [47]. Guiliano [48] suggested that C-60 (Bucky balls) is related with the formation and the ability to withstand the functional and structural aspects of microtubules, further, the quantum computational aspect can facilitate in understanding the role of microtubules in the cellular functionality. Grebowski, et al. [49] observed that fullerenol C-60(OH)36 got bound to ‘band 3protein’ and did not allow it to degrade, this was because of many OH groups present on the surface of fullerene cage got adsorbed on the protein component of cytoskeleton of red blood cell [49]. Graphene has been observed to be highly biocompatible, having higher loading capacity with lower degree of toxicity; graphene enhanced the process of stem cell differentiation in to estrogenic, neuronal and adipogeneic cell-line [50]. Hongwei and Shuming [51] have developed cell penetrating quantum dots (multivalent and endosomal disrupting having surface coating, luminescent and soluble with ligand exchanged), these quantum dots get internalized the cell and resulted in the disorganization of cell organelles and cytoskeleton; the ligand used were endosomes-disrupting polymers; like polyethylene (PEG) grafting with hyper branched polyethylene mine (PEG-g- PEI); these were used to investigate the movements of exchange of intracellular molecules [51]. Rivolta, et al. [52] observed that low temperature (from 37°C to 4°C) affected the intracellular distribution of coumarin-6- linked to solid nanoparticles, but the cytoskeleton was distorted and was detected with cytochalasin D, but when the effect of ATP dependent molecular motors were studied and least inhibition in the distribution of the detector was encountered [52]. These investigations were seen in the light of diffusion gradient based on interaction energy linked with the actin component of cytoskeleton.
Panariti, et al. [53] mentioned that component of cytoskeleton like actin and microtubules along with molecular motors-myosin-dynein and kinins controlled traffic along the membrane; molecular motors move on microtubules and actin filaments using them as tracks [53]. Katsumiti, et al. [54] have proposed the mechanism related to the interaction of CdS quantum dots and sub- lethal concentration (0.31 to 5 mg Cd/L) hemocytes and cells of gills of Mytilus galloprovincialis; this involved the production of ROS, catalase activity, DNA damage, lysosomal acid phosphatase activity, multixenobiotic resistance transport activity, Na/K ATPase activity in the cell of gills, phagocytic activity, damage to actin cytoskeleton in case of hemolytic, as a result of interaction with quantum dots but this interaction is not uniform in all the tissues [54]. Soenen, et al. [55] noticed the disturbance in cytoskeleton caused by the higher concentration of intercellular iron oxide nanoparticles and further the focal-adhesion mediated signaling was also affected adversely [55]. Conclusion and Prospectus Cell behavior is an overall reflection of the health of the cell and its ability to respond to intra and inter cellular stimuli. Cells, in all prime domains of organisms, are the epicenter of metabolic, biochemical and physic- physiological, and mechanotransductive interactions; these interactions may be beneficial and/or derogative in nature. Recent times have seen a bloom in the advancement of nanotechnology and nonmaterial’s natural and manmade; this has been the cause of multifaceted exploitation of these products in almost all fields like food technology [56]. Cosmetics, pharmaceuticals, nanoceuticals, industrial, communications, domestic, research and development etc. As a result biological systems, tissues and cells face the great influx of non-materials and the structural and functional unit of Bios stem – the cell has to face their impetus. In this review, overall impact of nonmaterial’s on the cytoskeleton – the structural and functional component of cell is evaluated. Cytoskeleton is the basis of temporal and spatial organization of cell. When nonmaterial’s like, carbon based nonmaterial’s, metals, metal oxide and semiconductors, are internalized and/or remain in contact with cell membrane, the cell membrane and cytoskeleton exhibit derogative changes related to cellular elasticity, mobility, adhesion and invasion, the degree of impact varies depending on the properties of nonmaterial’s and the nature of the cell membrane and the cell itself [22]. The nonmaterial’s are likely to be the cause of pathogenesis, clinical aberrations and the deviations from the normal pattern of cellular functioning; the interactions between cell membrane, cell organelles and nonmaterial’s may be sub toxic, sub lethal, toxic and lethal in nature. This scenario compels to look out for the consequences (good and bad both) in order to understand the mechanism involved in these interactions and to weed out the probable dangers lurking. Techniques like phase contrast, fluorescence technique and electron microscopy, cry electron microscopy can be utilized in investigation of cytoskeleton at different levels and to study the molecular changes in the components of cytoskeleton. There appears to be huge scope for further studies to understand the seemingly simple functional unit but complex in reality the cytoskeleton.
Conflicts of Interest
Author declares no conflict of the statement.
References
-
Lahir YK (2015) a dynamic component of tissues- extracellular matrix: structural, functional and adaptive approach. Biochem Cell Arch 15(2): 331- 347.
-
Becker WM, Kleinsmith L, Hardin J (2004) the world of the cell, 5th (Edn), Pearson Education.
-
Hermann H, Aebi U (2004) Intermediate filaments: molecular structure, assembly mechanism integration into functionally distinct intracellular scaffolds. Annul Review Biochem 73: 749-789.
-
Garcia E (2005) Molecular mechanism of phagocytosis, Ed Carlos Rosales, Landes Boscience/Euekah.com; Springer Science Business Media Inc USA.
-
Wickstead B, Gull K (2011) The evolution of the cytoskeleton. J Cell Biol 194(4): 513-525.
-
Mattila PK, Batista FD, Treanor B (2016) Dynamics of actin cytoskeleton mediator receptor Cross talk: an emerging concept in tuning receptor signaling. J Cell Biol 212(3): 267-280.
-
Riesler E (1993) Actin molecular structure and function. Current Opinion Cell Biol 5(1): 41-47.
-
Graceful P, Dominguez R (2003) Crystal structure of monomer actin in the ATP state, Structure basis of nucleotide-dependent actin dynamics. J Biol Chem 278(36): 34172-34180.
-
Narita A, Takeda S, Yamashita A, Maeda Y (2006) Structural basis of actin filament capping at the barbed end: a cryo-electron microscopy study. EMBOJ 25(23): 5626-5633.
-
Vindin H, Gunning P (2013) Cytoskeleton tropomyosins: choreographers of actin filament functional diversity. J Muscle Research Cell Motility 34(3-4): 261-274.
-
Bruce A, Alexander J, Julian L, Martin R, Keith R et al. (2002) Molecular biology of cell, 4th Edition, Taylor and Francis, ISBN 0-8153-4072-9.
-
Hui-Chia YK, William MB (2016) Collapsing Response Mediator Protein-1 (CRMP-1) Regulates Arp2/3 dependent Actin assembly. J Biol Chem 291(2): 658- 664.
-
Gunning PW, Ghosh DU, Whitaker S, Popp D, Robinson RC (2015) The evolution of compositionally and functionally distinct actin filaments J Cell Sci 128 : 2009-2019.
-
Vale RD (2003) the molecular motor tool-box for intracellular transport. Cell 112(4): 467-480.
-
Bartolilni F, Gundrsen GG (2006) Generation of non- centrosomal microtubule arrays. J Cell Sci 119(20): 4155-4163.
-
Ghosh DK, Dasgupta D, Guha A (2012) Models, regulations and functions of microtubule Severing by katanin. ISRN Molecular Biology 2012.
-
Hashimoto T (2015) Microtubules in plants, The Arabidopsis Book 13: 179.
-
Parker AL, Kavallais M, McCarroll JA (2014) Microtubules and their role in cellular stress In cancer. Frontiers in Oncology 4: 153.
-
Hermann H, Bar H, Kreplak L, Strelkov SV, Aebi U (2007) Intermediate filaments: from cell architecture to nanomechanics. Nature Review Molecular Cell Biol 8(7): 562-573.
-
Kreplok L, Fudge D (2007) Biomechanical properties of intermediate filaments: from tissues to single filaments and back. Bioassays 29(1): 26-35.
-
Lee CH, Kim MS, Chung BM, Leahy DJ, Coulombe PA (2012) Structural basis for Heteromeric assembly and perinuclear organization of keratin filaments. Nat Sturct Mol Biol 19(7): 707-715.
-
Wu YL, Putcha N, Nq KW, Leong DT, Lim CT, et al. (2013) Biophysical Responses upon the interaction of nanomaterials with cellular interfaces. Ace Chem Res 46(3): 782-779.
-
Robert B, Johnson A, Lewis J, Martin R, Keith R, et al. (2002) Molecular Biology of Cell, 4th ed, G S Garland Sc, Taylor and Francis Group USA 10: 0-8153-3218-1.
-
Gerald KG (2014) Cell Biology , 7th (Edn) John Wiley & Sons, Singapore, (P) Ltd.
-
Berry CC, Charles S, Wells S, Dalby MJ, Curtis AS (2004) The influence of transferring Stabilized magnetic NPs on human dermal fibroblast in culture. Int J Pharm 269(1): 211-225.
-
Lee J, Chu BH, Chen KH, Ren F, Lele TP (2009) randomly oriented upright SiO2 coated nanorods for reduced adhesion of mammalian cells. Biomaterials 30(27): 4480-4493.
-
Apopa PL, Qian Y, Shao R, Guo NL, Schwegler Berry D, et al. (2009) Iron oxide NPs induce human microvascular endothelial cell Permeability through reactive species of oxygen production and microtubule remodeling. Part Fibre Toxicol 6: 1.
-
Wu X, Tan Y, Mao H, Zhang M (2010) Toxic effects of iron oxide on human umbilical vein epithelial. Cells Int J Nanomedicine 5: 385-399.
-
Tiwari A, Patra H, Choi JW (2015) Advanced theranostics materials, Scrivener Publishing, Wiley Co-published by John Wiley and Sons, Inc Hoboken, New Jersey.
-
Soenen SJ, Illyes E, Vercauterenen D, Breackman SK, Majer Z, et al. (2009) The Role of nanoparticles concentration dependant induction of cellular stress in the Internalization of non-toxic cationic magnetolioposomes. Biomaterials 30(36): 6803-6813
-
Singh N, Jenkins GJS, Asadi R, Doak SH (2010) Potential toxicity of super paramagnetic iron oxide nanoparticles, Nano Reviews 1: 5358.
-
Fucikova A, Valetta J, Plant I, Kalbacova MH, Broz A, et al. (2014) Silicon nanocrystals and nanodimonds in line cells: photoluminescence characterization cytotoxicity and interaction with cell cytoskeleton. RSC Adv 4.10334-10342.
-
Vijaykumar CC, Venkatkrishnan K, Bo Tan (2015) Harmonizing He La cells cytoskeleton behavior by multi TiO2 phased nanostructure synthesized through ultra short pulsed laser. Scientific Report 5: 15294.
-
Mironava T, Hadjiargyrou M, Simon M, Jurukovaski, Rafilovich MH (2010) Silver nanoparticlescellular toxicity and recovery: effect of size, concentration and exposure time. Nan toxicology 4(1): 120-137.
-
Xu F, Piett L, Farkas S, Qazzaz M, Syed NI (2013) Silver nanoparticles cause degeneration of cytoskeleton and disrupt synaptic machinery of cultured cortical neuron. Mob Brain 6:29.
-
Liu YX, Karsai A, Anderson DS, Silva RM, Uyeminami DL (2015) Single cell mechanism provides an effective means to probe in vivo interactions between alveolar macrophages and Ag NPs. J Physical Chem- B 119(49): 15118-15129.
-
Cooper RJ, Spitzer N (2015) Silver nanoparticles disrupt cytoskeleton and neurotic dynamics in cultured adult Neural stem cells. Neurotoxicity 48:231-238.
-
Katsumiti A, Gilliland D, Arostegui I, Cajaraville MP (2015) Mechanism of toxicity of silver nanoparticles in comparison to bulk and ionic silver on Mussel hemocytes and gill cells. 10(6): 0129039.
-
Bohmert L, Nieman B, Lichtenstein D, Juing S, Lampen A (2015) Molecular mechanism of silver nanoparticles in intestinal cells. Nanotoxicology 9(7).
-
Pampaloni F, Florin E (2008) Microtubule architecture: inspiration for novel Carbon nanotubes based biomimetic materials. Trends in Biotechnology 26(6): 302-310.
-
Holt BD, Short PA, Rape AD, Wang YL, Islam MF, et al. (2010) Carbon nanotubes recognize actin structures in cells and ex vivo. ACS Nano 4(8): 4872-4878.
-
Ying D, Sun H, Li X, Li X, Zhao L (2016) Impact of carbon nanotubes on actin Polymerization. J Nanosci Nanotech 16(3):2408-2417.
-
Boyer AD, Ganesh S, Qin Z, Holt BD, Buehler MJ, et al (2016) Delivering SWCNTs to nucleus using nuclear protein domains, ACS Appl Materials and Interfaces. 8(5):3524-3534.
-
Lahir YK, (2012) Apoptosis – A Biological Phenomenon. Biochem Cell Arch 12(2): 237-248.
-
Youle RJ, Karbowski M (2005) mitochondrial fission in apoptosis. Nat Rev Mol Cell Biol, 6(8):657-663.
-
Johnston HJ, Hutchison GR, Christensen FM, Aschberger K, Stone V (2010) The biological Mechanism and physicochemical characteristics responsible for driving fullerene toxicity. Toxicol Sci 114(2):162-182.
-
Ratnikoval TA, Govindan PN, Salonene E, Ke PC (2011) In vitro polymerization of Microtubules with fullerene derivatives. ACS Nano 5(8): 6305-6314.
-
Guiliano V (2012) Bucky balls, health and longevity- state of knowledge.
-
Grebowski J, Krokos A, Puchala M, (2013) Fullerenol C-60(OH) 36 could associate to Band 3-proteins of human erythrocyte membrane. Biochimica et Biophysica Acta (BBA)- Biomembrane 1828(9): 2007- 2014.
-
Bressan E, Ferroni L, Gardin C, Sbricoli L, Gubato L, et al. (2014) Graphene based scaffolds effects on the stem cells. Commitment, J Translation Medicine 12:296
-
Hongwei D, Shuming N (2007) Cell penetrating quantum dots based on multivalent and endosomal disrupting surface coating, J Am Chem Soc 129 (11): 3333-3338.
-
Rivolta I, Panariti A, Collini M, Le-Hiero B, D’ Alfonso L, et al. (2011) biophysical model of intracellular distribution and perinuclear accumulation of particulate matter. Biophys Chem 158(2-3):134-140.
-
Panariti A, Miserocchi G, Rivolta I (2012) The effects of nanoparticles uptake on cellular behavior: disrupting and enabling functions. Nan Techno Sic Apple 5:87-100.
-
Katsumiti A, Gilliland D, Arostegui I, Cajaraville MP (2014) Cytotoxicity and cellular Mechanism involved in the toxicity of CdS quantum dots in hemocytes and gill cells of Mytilus galloprovincialis, Aquatic Toxicol 153:39-52.
-
Soenen SJ, Nuytten N, DeMeyer SF, De Smedt SC, De Cuyper M (2010) High intracellular iron oxide nanoparticles concentrations affected cellular cytoskeleton and focal adhesion kinase-mediated signaling, Small 6(7): 832-842.
-
Lahir YK (2015) Role and adverse effects of nanomaterials in food technology. Journal of Toxicology and Health 2.
- Evaluation of Skin Aging Preventive Effects of Cherry Blossom Petal Extracts Through Antioxidant and Anti-Glycation Activities
- Is Cell Death Responsible for False Positive Results of In Vivo Comet Assay?
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism