Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3 Alpha/Beta in the in-vitro Amyloid-beta Toxicity Model
The neurodegenerative diseases such as Alzheimer’s disease (AD) become more common globally due to great effort on providing longer lifespans for human beings. Besides, the researches related to amelioration of age- related diseases by epigenetic influences gain acceleration to solve the molecular mechanism of these diseases or to improve care and opportunities of the patients. Herein, we aimed to investigate the neuroprotective and/or therapeutic effects of both SIRT1 activation and inhibition as an epigenetic influence on the exosome releasing amount and exosomal protein content in the human neuroblastoma cell line under Aβ toxicity. Cells were treated with SIRT1 activator (CAY10602, 20 μM) and SIRT1 inhibitor (Ex527, 80μM) after or before Aβ toxicity. Addition to LDH cytotoxicity tests, cell lysates and immunoprecipitated exosome samples were analyzed by western blot with respect to proteins in the PI3K/AKT/GSK3/mTOR pathway. According to the western blot analysis, SIRT1 inhibitor increased the level of phosphorylated AKT, and inhibits the GSK3α and GSK3β by increasing their phosphorylated levels; also, it decreased the amount of phosphorylated mTOR. Therefore, SIRT1 inhibitor might have protective and therapeutic effect on amyloid beta toxicity. On the other hand, SIRT1 activator increased the level of SIRT1 protein and PTEN phosphorylation. In the exosomal content, Aβ toxicity increased the level of GSK3β and decreased its phosphorylation. Interestingly, both SIRT1 activator and inhibitor produced similar neuroprotective and therapeutic effect on the Aβ toxicity in the exosomes. These results indicate that SIRT1 inhibitor and activator may be neuroprotective in the pathophysiology of AD in particular points through different molecular pathways.
Introduction
With an increase in the lifespan of human beings worldwide, the diseases related to senility such as Alzheimer’s disease (AD), which constitutes 60-80% of all dementia cases, have been started to affect more people. According to the World Health Organization’s report (2016), AD and other dementias are the 5th leading causes of death globally, which were not on the list in the year 2000. AD is a progressive neurodegenerative disorder which causes cognitive function and memory impairments in elderly. Due to the high cost of treatment expenses and complete dependence of patients to their caregivers, AD adversely affects the life of people themselves, their caregivers and the economy of the governments [1, 2, 3]. Nowadays, the investigators focus on the researches in the area of neurodegenerative disease either to solve the molecular mechanism of disease or to improve care and opportunities of the patients.
Main pathological hallmarks of AD are intercellular senile plaques mainly composed of amyloid beta, and intracellular neurofibrillary tangles produced by hyperphosphorylated tau. In clinical perspective, synapse and neuron loss; brain atrophy; decrease in particular neurotransmitters and correspondingly memory impairments, and problems in motor functions such as speaking and walking are the AD symptoms which are getting worse by time [4, 5, 6, 7, 8, 9]. Investigations showed that environmental factors such as hypertension, obesity, smoking, depression, low level of physical activity, and low education level could increase the risk of AD, and if these factors are eliminated the risk could be decreased as much as 30 percent [4, 8]. Therefore, modifying the epigenetic influences in the daily life may affect gene expression of the diseased-state. Sirtuins are one of these molecules which have been demonstrated to have important epigenetic roles in most of the pathological conditions.
Certain in-vivo and in-vitro studies showed that overexpression and enzyme activation of SIRT1 which is one of the sirtuins having an NAD+dependent enzyme activity to deacetylase histone and non-histone proteins from their lysine residues increase α-secretase expression. Therefore, SIRT1 decreases Aβ production, increases neuronal survival and provides neuroprotection by supporting non-amyloidogenic pathway in the brain. According to the previous studies, SIRT1 activates BDNF and improve memory and cognitive functions. However, some other studies done by nicotinamide which is one of the SIRT1 inhibitors enhance cognitive functions and Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
memory, also decreased phosphorylated tau (Thr231) amount [10, 11, 12, 13, 14, 15, 16, 17, 18, 19].
In addition to epigenetic influences, the circulating molecule in the blood varies depending on the health of the cell and pathological state. It was also shown that the metabolites and molecules in the blood are carried with different mechanisms. Exosomes are one of these transport systems which can be also used as a biomarker for diagnosis in the neuro degenerative diseases such as AD [20, 21].
Exosomes can carry RNA, protein, lipid and their cargo content can change according to the cell they are originated. In physiological conditions, they have role in cell to cell communication, discarding unwanted molecules, neuronal development and protection, nerve regeneration, and synaptic plasticity [20, 22, 23, 24, 25]. In AD, it is shown that exosomes carry Aβ, tau and disease-related miRNAs and exosomal marker proteins such as Alix and Flotillin-1 are found in amyloid beta plaque regions [22, 23, 26, 27, 28, 29, 30, 31, 32, 33]. There are different perspectives about role of exosomes in AD. For instance, Abdullah, et al. Proposed that exosomes help clearance of Aβ and the level of exosome was decreased in astrocyte cells in response to Aβ treatment [34], where other studies claim that exosomes aid the spread of disease via carrying toxic proteins from cell to cell and their levels may increase in the case of AD [26, 35]. Therefore, in the current study, we aimed to investigate the neuroprotective and/or therapeutic effects of both SIRT1 activation and inhibition on the exosome releasing amount and exosomal protein content in the human neuroblastoma cell line with Aβ toxicity.
Materials and Methods
Cell Culture
SH-SY5Y human neuroblastoma cell line (ATCC CRL- 2266, USA) was used for the experiments. SH- SH5Y cells were cultured and passaged using DMEM medium (Biosera, Cat. No: LM-D1099, France) containing 10% (v/v) heat activated fetal bovine serum (Biosera, Cat. No: FB-1001H/100, France) and 1% (v/v) penicillin- streptomycin (Biosera, Cat. No: LM-A4118/20, France).
Aβ1-42, SIRT1 Activator and SIRT1 Inhibitor Preparation
221.5 μM stock solution of Aβ1-42 (Abcam, Cat. No: ab120301, USA) was prepared by dissolving in 1% (v/v) NH4OH (Sigma-Aldrich, Germany) and incubated for 48 hours at 37oC before used in experiments. 10 mg of activator (CAY10602; 3-(Benzenesulfonyl)-1-(4- fluorophenyl)pyrrolo[4,5-b]quinoxalin-2-amine, SIRT1 activator II; Calbiochem, Cat No: 566313, Germany) and 5 mg of inhibitor (Ex527; 6-Chloro-2,3,4,9-tetrahydro- 1H- carbazole-1-carboxamide;Calbiochem, Cat No: 566322, Germany) were dissolved in 400μl and 750 μl of DMSO, respectively and aliquots were stored at -20oC.
Lactate Dehydrogenase (LDH) Assay
The assay was done according to the manufacturer’s instructions with little modifications (Pierce, Cat. No: 88953, USA). Briefly, cells were seeded with a density of 10.000 cells/well in a 96-well plate, and after 24 hours, 6 different SIRT1 activator (5μM, 10μM, 20μM, 40μM, 60μM, 80μM) and inhibitor (98nm, 10μM, 20μM, 40μM, 80μM, 100μM) doses were applied as triplicates to find nontoxic doses. After 24-hour incubation, 50 μl of culture media from all wells were transferred to new 96-well plate, 50 μl of reaction mix was added and incubated for 30 minutes at room temperature in the dark. Then, 50 μl of stop solution was added. The absorbance values were measured by using microplate reader (Chromate Manager 4300, Palm City/ USA) at 492 nm as primary filter and at
630 nm as secondary filter.
Establishing In-vitro Aβ1-42 Model
Establishment of in-vitro Aβ1-42 model was done by LDH assay, according to manufacturer’s instructions (Roche, Cat No: 11644793001, Germany). Shortly, cells were seeded in a 96-well plate with 10.000 cell/well density. After 24-hour, four different Aβ1-42 doses (1.25μM, 2.5μM, 5μM, 10μM) were applied as triplicate and incubated for 24 hours at 37oC. Working solution of the kit was prepared and incubated for 15 min in the dark at room temperature. After addition 100μl of LDH working solution in another 96-well plate the absorbance values were measured using microplate reader (Chromate Manager 4300, Palm City/ USA) at 492 nm.
Treatment of Cells with SIRT1 Activator/ SIRT1 Inhibitor and Aβ1-42
This experiment is consisted of 10 groups which were normalized according to their vehicle data (1) Control (only cell); (2) Aβ1-42; (3) only activator; (4)(activator + Aβ1-42); (5) Aβ1-42 + activator; (6) Activator + Aβ1-42; (7) only inhibitor; (8) (inhibitor+ Aβ1-42); (9) Aβ1-42 + inhibitor; (10) inhibitor + Aβ1-42(Table 1).
| Groups | 48th Hour | 72nd Hour | 96th Hour | ||
| 1 | Control | Cell Seeding | - | - | |
| 2 | Aβ | Cell Seeding | Aβ | - | Cell Harvesting |
| 3 | CAY10602 (activator) | Cell Seeding | CAY10602 | - | Cell Harvesting |
| 4 | (CAY10602+Aβ1-42) | Cell Seeding | CAY10602+Aβ1-42 | - | Cell Harvesting |
| 5 | Aβ1-42+CAY10602 | Cell Seeding | Aβ1-42 | CAY10602 | Cell Harvesting |
| 6 | CAY10602+Aβ1-42 | Cell Seeding | CAY10602 | Aβ1-42 | Cell Harvesting |
| 7 | Ex527 (Inhibitor) | Cell Seeding | Ex527 | - | Cell Harvesting |
| 8 | (Ex527+Aβ1-42) | Cell Seeding | Ex527+Aβ1-42 | - | Cell Harvesting |
| 9 | Aβ1-42+Ex527 | Cell Seeding | Aβ1-42 | Ex527 | Cell Harvesting |
| 10 | Ex527+Aβ1-42 | Cell Seeding | Ex527 | Aβ1-42 | Cell Harvesting |
Table 1: Aβ/SIRT1 activator (CAY10602) and inhibitor (Ex527) treatment groups according to the time of substance Table 1: Aβ/SIRT
Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
Exosome Isolation
Exosome isolation from cell culture medium was done according to the manufacturer’s instructions (Norgen, Cat. No: 60400, Canada). Basically; medium collected from each group was centrifuged at 1000 rpm for 15 minutes to remove cellular debris. Supernatants were mixed with 2.75μl of ExoC buffer and 200μl of Slurry E. After 5 minutes incubation at room temperature, tubes were centrifuged at 2000 rpm for 2 minutes, supernatants were discarded. Pellets were re-suspended with 200 μl of Exo R buffer and centrifuged at 500 rpm and at 6000 rpm for 1 minute. Supernatants which include exosomes were collected and stored at -20°C.
Exosome Immunoprecipitation
Exosome immunoprecipitation was done by using Pure Proteome Protein G Magnetic Beads (Millipore, Cat No: LSKMAGG02, USA) according to manufacturer’s indirect immunoprecipitation protocol with little modifications. Briefly, 100μl of exosome solution and 2μl of Alix antibody (Cell Signaling, Cat. No: 2171s, USA) were mixed and incubated overnight at 4oC with continuous mixing. After incubation, 50 μl G-protein magnetic beads were added to the exosome antibody mix and incubated at room temperature for 10 minutes with continuous mixing. After the incubation samples were removed and the beads were made ready for denaturation.
Western Blot
Cell lysates and immunoprecipitated exosome samples were denatured. Protein concentration of cell lysates were measured spectrophotometrically (Implen, Germany). Exosomes and 40 μg of total protein from each lysate were separated by NuPAGE 4-12% Bis-Tris gel and then proteins were transferred to PVDF membrane using iBlot Dry Blotting System (Invitrogen, USA). Membranes were blocked using 5% non-fat milk in 50 mM Tris- buffered saline containing 0.1% Tween-20 (TBS-T; blocking solution) for 1 hour at room temperature and incubated overnight in primary antibody solutions as follows: SIRT1 (1:1000; Cell Signaling, Cat. No: 9475s, USA); phospho-AKT (1:1000; Cell Signaling, Cat. No: 9271s, USA); AKT (1:1000; Cell Signaling, Cat. No: 4685s, USA); phospho-GSK3α/β (1:1000; Cell Signaling, Cat. No: 8566s, USA); GSK3α/β (1:1000; Cell Signaling, Cat. No: 5676s, USA); phospho-mTOR (1:500; Cell Signaling, Cat. No: 5536s, USA); mTOR (1:500; Cell Signaling, Cat. No: 2983s, USA); phospho-PTEN (1:1000; Cell Signaling, Cat. No: 9554s, USA); PTEN (1:1000; Cell Signaling, Cat. No: 9559s, USA); Tsg101 (1:500; Santa Cruz, Cat. No: sc-7964, Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
USA); antibodies were diluted in blocking solution. Next day, membranes were washed with TBS-T for 3 times and incubated with HRP- conjugated anti-mouse (1:2500; Cell Signaling, Cat. No: 7076s, USA); and anti-rabbit (1:2500; Cell Signaling, Cat. No: 7074s, USA) secondary antibodies within blocking solution for 1 hour at room temperature. β-actin (1:1000; Cell Signaling, Cat. No: 3700s, USA) was used as loading control. Blots were developed using Western Bright Sirius detection kit (Advansta, USA) and visualized by Bio-Rad ChemiDoc XRS System (Bio-Rad Laboratories Inc., USA). Protein levels were analyzed densitometrically with ImageJ software (Image J; National Institute of Health, Bethesda, MD, USA) and total protein levels are corrected with values obtained by β-actin blots where phosphorylated protein levels are corrected with values obtained by total protein blots.
Total Antioxidants Status (TAS)-Total Oxidant Status (TOS) Assay
The TAS and TOS levels of cell culture media were detected by automated measurement method using automated analyzer (Chromate Manager 4300, Palm City/USA) as described previously according to manufacturer’s protocol (Rel Assay Diagnostics, Gaziantep/TURKEY). Briefly, while measuring the TAS level, the colored dianisidyl radicals’ absorbance was monitored to detect Fenton reaction. The samples’ antioxidative effects were expressed in mmolequiv/L Trolox. In the measurement of TOS, the oxidant levels in the samples were determined by measuring the absorbance level resulted from colored reaction of oxidation of ferrous-ionodianisidine complex to the ferric ion. The TOS level is expressed in μmol H2O2equiv/L. Oxidative index status was calculated by the following formula:
OSI = [(TOS)/(TAS × 1000)] × 100
Statistical Analysis
Statistical analyses were performed by one-way ANOVA followed by Tukey’s LSD test using SPSS for Windows 18. All values are given as mean ±SD and p values ≤ 0.05 is considered as significant.
Results
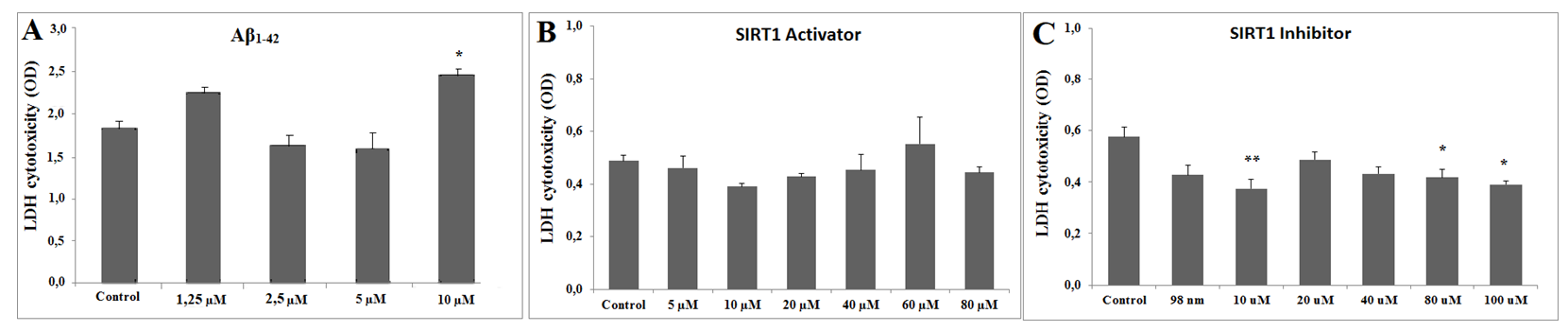
Cytotoxicity Assay
According to the LDH cytotoxicity assay, the toxic dose of Aβ concentration was determined as 10 µM (Figure 1A). In order to find the non-toxic dose of SIRT1 activator (CAY 10602) and inhibitor (Ex527), six different doses of the substances were applied to the SH-SY5Y cells. For the SIRT1 activator, there was no statistically significant difference between the control group and activator groups in the LDH cytotoxicity assay. Therefore, 20µM concentrations for the SIRT1 activator were chosen according to the previous literature [36]. For the SIRT1 inhibitor, 10µM, 80µM and 100µM concentrations for the SIRT1 inhibitor were significantly reduced the cytotoxicity, according to the LDH assay results and previous literature [37], 80 µM concentration of SIRT1 inhibitor was chosen for further experiments (Figure 1B & 1C).
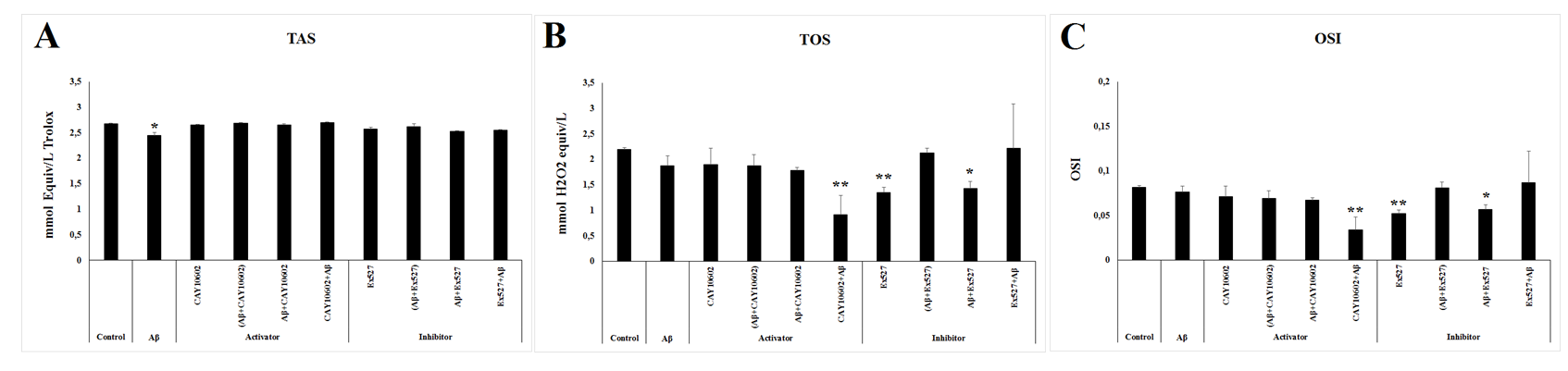
Levels of TAS, TOS and OSI in SIRT1 Activator (CAY 10602) /Aβ and SIRT1 Inhibitor (Ex527)/Aβ Groups
The TAS level were similar in all experimental groups except in Aβ group whose level significantly decreased compared to the control group (p≤0.05) (Figure 2A). On the other hand, both TOS and OSI level were significantly low in the CAY10602+Aβ, Ex527, and Aβ+Ex527groups compared to the levels in the control group (p≤0.05) (Figure 2B & 2C).
Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
Protein Expression Levels in Response to Aβ/SIRT1 Activator and Aβ/SIRT1 Inhibitor Treatment
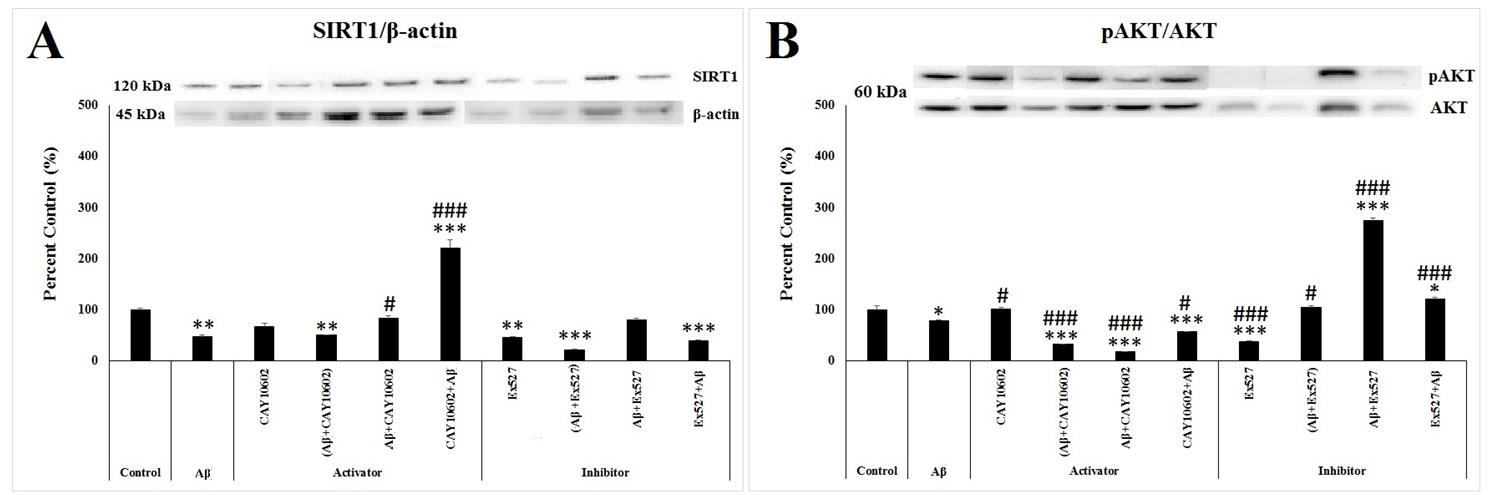
According to one way ANOVA, there was a significant between-group differences in the SIRT1 level (F(9;29)=53.735, p<0.001). As shown in Figure 3A, the SIRT1 protein level in the Aβ (p=0.003), (Aβ+CAY10602) (p=0.006), Ex527 (p=0.002), (Aβ+Ex527) (p≤0.001), and Ex527+Aβ (p=0.001) groups decreased significantly, while its level significantly increased in the CAY10602+Aβ group (p≤0.001) compared to the control group. Meantime, a significant increase in the SIRT1 level was observed in the Aβ+CAY10602 (p=0.005) and CAY10602+Aβ (p≤0.001) groups compared to the Aβ group (Figure 3A).
According to one way ANOVA, there was a significant between-group differences in the activity of AKT protein which was calculated from the ratio of phosphorylated AKT to the total AKT (F(9;29)=280.769, p≤0.001). As a result of only Aβ treatment, p-AKT level marginally decreased compared to the control group (p=0.064). In addition, it was seen that there was a further significant decrease in the (Aβ+CAY10602), Aβ+CAY10602, CAY10602+Aβ, and Ex527 groups (p≤0.001), while there was a significant increase in the Aβ+Ex527 (p≤0.001) and Ex527+Aβ (p=0.05) groups compared to the control group (Figure 3B). Compared to the Aβ group, the ratio of p-AKT to the AKT significantly decreased in the (Aβ+CAY10602) (p≤0.001), Aβ+CAY10602 (p≤0.001), CAY10602+Aβ (p=0.039), and Ex527 (p≤0.001) groups and it was significantly increased in the CAY10602 (p=0.05), (Aβ+Ex527) (p=0.012), Aβ+Ex527 (p≤0.001), Ex527+Aβ (p≤0.001) groups. (Figure 3B).
According to one way ANOVA, there was a significant between-group differences in the activity of PTEN protein which was calculated from the ratio of phosphorylated PTEN to the total PTEN (F(9;29)=49.576, p≤0.001). In the Aβ group, the activity of PTEN decreased significantly compared to the control group (p≤0.001). In addition, the activity of PTEN decreased in the (Aβ+CAY10602) (p≤0.001), Aβ+Ex527 (p=0.001), and Ex527+Aβ (p=0.019) groups, while there was a significant increase in the (Aβ+Ex527) (p≤0.001) group compared to the control group (Figure 3C). Compared to the Aβ group, the ratio of p-PTEN to the PTEN significantly increased in the CAY10602 (p=0.001), Aβ+CAY10602 (p=0.002), CAY10602+Aβ (p≤0.001), Ex527 (p≤0.001) and (Aβ+Ex527) (p≤0.001) groups (Figure 3C).
According to one way ANOVA, there was a significant between-group differences in the activity of mTOR protein which was calculated from the ratio of phosphorylated mTOR to the total mTOR (F(9;29)=12.637, p<0.001). According to post hoc Tukey’s test, the activity of mTOR decreased in the Ex527 (p=0.008), (Aβ+Ex527) (p=0.004), and Ex527+Aβ (p=0.015) groups (Figure 3D). Compared to the Aβ group, the ratio of p- mTOR to the mTOR significantly decreased in the Ex527 (p≤0.001), (Aβ+Ex527) (p≤0.001), Aβ+Ex527 (p=0.036), and Ex527+Aβ (p=0.001) groups (Figure 3D).
Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
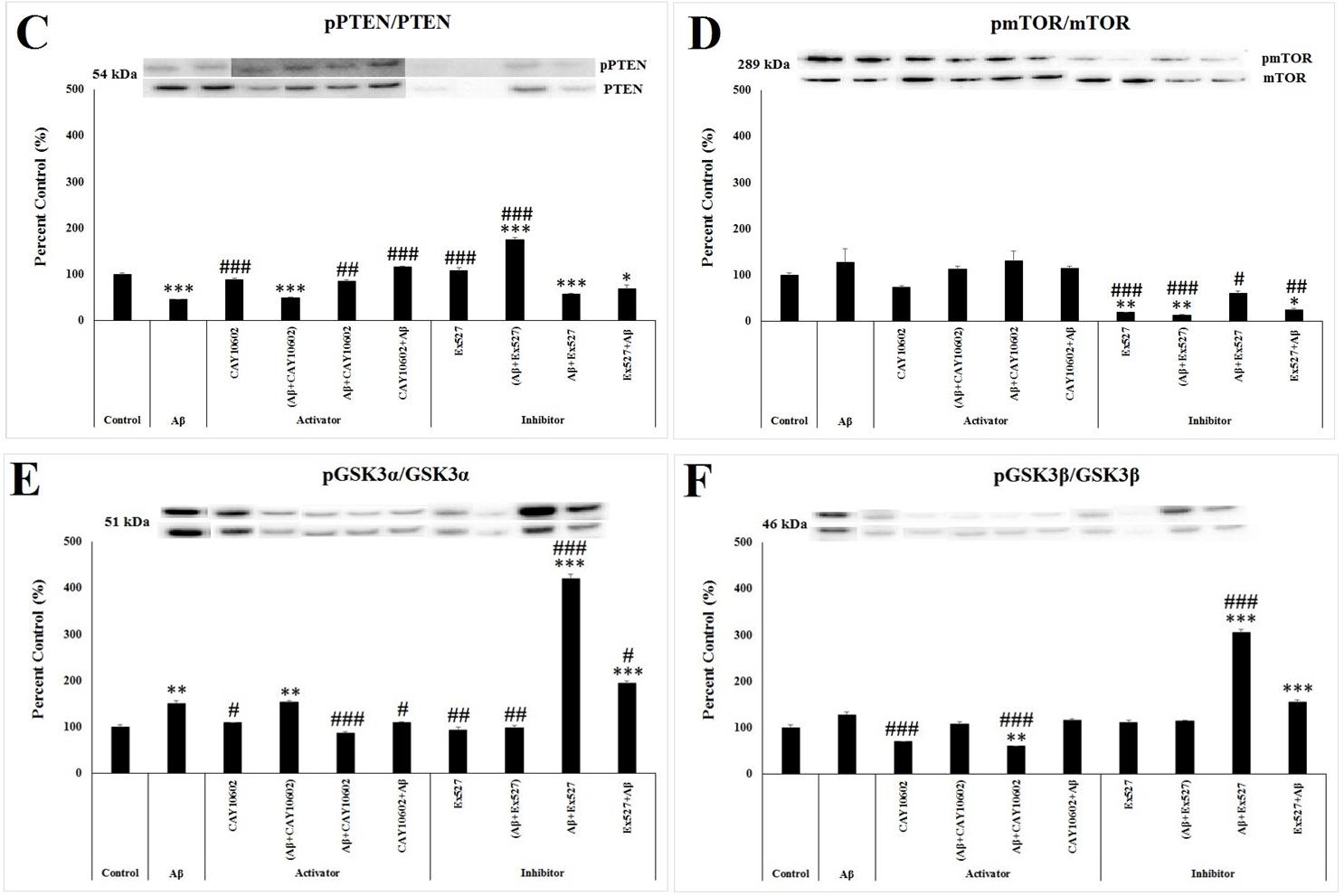
According to one way ANOVA, there was a significant between-group differences in the ratio of phosphorylated GSK3α/β to the total GSK3α/β (F(9;29)=159.707, p≤0.001; F(9;29)=110.242, p≤0.001, respectively). According to post hoc Tukey’s test, the level of phosphorylated GSK3α increased significantly in the Aβ (p=0.007) and (Aβ+CAY10602) (p=0.004) groups (Figure 3E) while the level of phosphorylated GSK3β decreased in the Aβ+CAY10602 (p=0.01) (Figure 3F) compared to the control group. In addition, the level of both phosphorylated GSK3α/β protein significantly increased in the Aβ+Ex527 and Ex527+Aβ groups compared to the control group (p≤0.001) (Figure 3E & 3F). Compared to the Aβ group, the ratio of p-GSK3α to the GSK3α significantly decreased in the CAY10602 (p=0.039), Aβ+CAY10602 (p=0.001), CAY10602+Aβ (p=0.049), Ex527 (p=0.002), and (Aβ+Ex527) (p=0.005) groups Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
while its ratio significantly increased in the Aβ+Ex527 and Ex527+Aβ groups (p≤0.001) (Figure 3E). Compared to the Aβ group, the ratio of p-GSK3β to the GSK3β significantly decreased in the CAY10602 (p<0.001) and Aβ+CAY10602 (p<0.001) groups while its ratio significantly increased in the Aβ+Ex527 group (p<0.001) (Figure 3F).
Exosome Releasing Amount in Aβ/SIRT1 Activator and Aβ/SIRT1 Inhibitor Groups
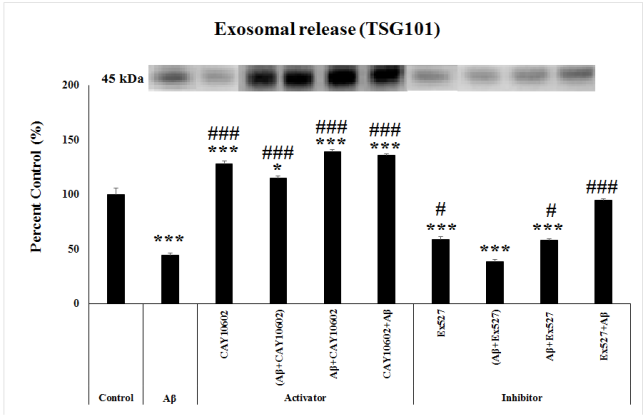
After immunoprecipitation of exosomes, western blot assay for TSG101 performed and the amount of exosomes were found as follows. According to one way ANOVA, there was a significant between-group differences in the exosome releasing amount determined by the level of TSG101 protein (F(9;29)=104.704, p≤0.001)Post hoc Tukey’s test showed that the level of TSG101 significantly decreased in the Aβ, Ex527, (Aβ+Ex527) and Aβ+Ex527 groups (p≤0.001), while there was a significant increase in the CAY10602, Aβ+CAY10602, CAY10602+Aβ (p≤0.001), and (Aβ+CAY10602) (p=0.012) compared to the control group. Compared to the Aβ group, there was a significant increase in the CAY10602, (Aβ+CAY10602), Aβ+CAY10602, CAY10602+Aβ, Ex527+Aβ (p≤0.001), Ex527 (p=0.012) and Aβ+Ex527 (p=0.019) groups (Figure 4).
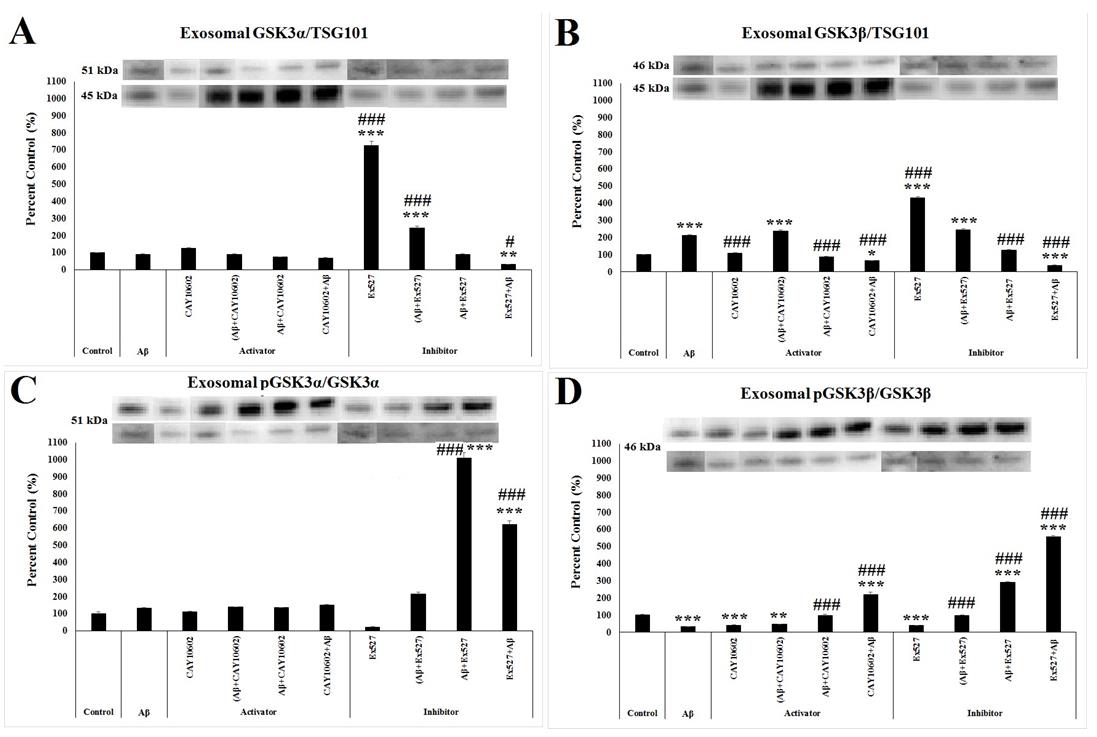
Exosomal p-GSK3α/β content in Aβ/SIRT1 Activator and Aβ/SIRT1 Inhibitor Groups
After immunoprecipitation of exosomes, while western blot assay for SIRT1, AKT, PTEN, GSK3α/β and mTOR was performed, + only GSK3α/β was detected in exosomal content. According to one way ANOVA, there was a significant between-group differences in the total expression of GSK3α/β protein in response to TSG101, which is an exosomal marker found at the surface of exosomes and whose expression level indicates the amount of exosomes released by cells (F(9;29)=391.572, p≤0.001; F(9;29)=273.062, p≤0.001, respectively). According to post hoc Tukey’s test, the level of exosomal GSK3α significantly increased in the Ex527 and (Aβ+Ex527) groups (p≤0.001) while its level decreased in the Ex527+Aβ group (p=0.005) compared to the control group (Figure 5A). While there was no significant change in the expression of exosomal GSK3α between the control and Aβ group, exosomal GSK3β increased in the Aβ group compared to the control group (p≤0.001) (Figure 5B). In addition, its level also increased in the (Aβ+CAY10602) (p≤0.001), (Aβ+Ex527) (p≤0.001) and Ex527 (p≤0.001) groups, and it decreased in the CAY10602+Aβ (p=0.049) and Ex527+Aβ (p≤0.001) groups compared to the control group (Figure 5B). Compared to the Aβ group, the amount Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
of GSK3β decreased in the CAY10602, Aβ+CAY10602, CAY1060 2Aβ, Aβ+Ex527, and Ex527+Aβ groups and its amount increased in the Ex527 group (p≤0.001) (Figure 5B).
In addition, we also determined the activity of GSK3α/β in the exosome by calculating the ratio of phosphorylated GSK3α/β to the total GSK3α/β and we found a significant between-group differences (F(9;29)=122.513, p<0.001; F(9;29)=440.357, p<0.001, respectively). While the activity of GSK3α did not change due to only Aβ administration, its activity significantly increased in the Aβ+Ex527and Ex527+Aβ groups (p<0.001) (Figure 5C). However, Aβ administration significantly decreased the activation of GSK3β compared to the control group (p<0.001). Its activity also decreased in the CAY10602 (p=0.001), (Aβ+CAY10602) (p=0.004), and Ex527 (p=0.001) groups and the activity of GSK3β increased in the CAY10602+Aβ (p<0.001), Aβ+Ex527 (p<0.001), and Ex527+Aβ (p<0.001) groups compared to the control group (Figure 5D). Compared to Aβ group, the ratio of p-GSK3β to the GSKβ significantly increased in the Aβ+CAY10602, CAY10602+Aβ, (Aβ+Ex527), Aβ+Ex527, and Ex527+Aβ groups (p<0.001) (Figure 5D).
Discussion
The expression of SIRT1 protein decreased in neuropathological conditions and particularly in the elderly [12, 18]. According to the previous studies, both activation/overexpression and inhibition of SIRT1 have some positive impacts on AD pathology [12, 13, 15, 16, 18, 19]. Therefore, in the present study, we aimed to investigate the detailed molecular mechanism of this relation through analyzing both SIRT1 activation and inhibition on the exosome releasing amount and exosomal protein content in Aβ toxicity. Previous studies performed in animals and cell culture has shown that Aβ toxicity may cause oxidative stress which is a fundamental feature of neurodegenerative diseases like AD [38, 39, 40]. In the current study, a significant decrease in Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
the amount of antioxidants with Aβ toxicity was noted as stated previous studie s showing the relation between Aβ toxicity and antioxidant deficiency [41, 42]. Interestingly, imbalance in the level of antioxidant status due to Aβ administration ameliorated by addition of both SIRT1 activator and inhibitor. This defensive response of alteration in SIRT1 activity against to free radicals produced by Aβ toxicity may be related to SIRT1’s antioxidative properties in addition to cell’s compensatory action against to inhibition of SIRT1 activity. For example, activation of SIRT1 by resveratrol, a natural SIRT1 activator, prevents Aβ-induced oxidative stress [43] and inhibition of SIRT1 by sirtinol, a class III HDAC inhibitor, restores alpha-synuclein toxicity-induced oxidative stress [44] suggesting the presence of SIRT1 in different antioxidant pathways.
The presence of SIRT1 in the several key pathways linked to progression of AD such as deacetylation of tau, promotion of the non-amyloidogenic APP processing pathway, protection against to oxidative stress, points out that decreased SIRT1 level results AD pathology due to attenuation of SIRT1’s neuroprotective role [15, 45, 46, 47]. However, some of the previous studies also showed that inhibition of SIRT1 may restore cognitive functions and decrease the amount of phosphorylated tau [16, 18, 19]. In the present study, we also noted a decrease in the SIRT1 expression due to addition of Aβ to the cells. In addition, studies proposed that SIRT1 enzyme activators and inhibitors work in a way that protein expression does not altered while the enzyme is activated or inhibited respectively [48, 49, 50]. However, in the present study, treatment of Aβ-induced cells with SIRT1 activator ameliorated the SIRT1 level. Interestingly, protective effect of SIRT1 activator was stronger in the stimulation of SIRT1 expression.
In experimental models and post-mortem patient brain tissues showed that AKT level and its activation decreased in AD [51, 52, 53]. In our study, we observed a significant decrease in the phosphorylation of AKT by Aβ toxicity. However, the addition of SIRT1 activator to the cells before or after Aβ toxicity cannot be effective in the activation of cell survival. Interestingly, SIRT1 inhibitor removed the Aβ-induced inhibition of AKT, especially when it was given therapeutically. Also, in our study, we observed a significant increase in the phosphorylation of both GSK3α/β suggesting an inhibition of GSK3α/β in the both therapeutic and protective treatment groups of SIRT1 inhibitor parallel to AKT phosphorylation. In a previous study, it is found that activation of GSK3α increase Aβ production [54] and activation of GSK3β cause tau hyperphosphorylation and eventually memory impairment and neuronal death [55]. Therefore, SIRT1 inhibitor might show neuroprotective and therapeutic effect by increasing cell survival via activating AKT/GSK3 axis. In addition to this, inhibition of SIRT1 decreased the phosphorylation of mTOR which has been regulated by AKT. It was suggested that mTOR regulates Aβ generation or Aβ clearance by regulating several keys signaling pathways, including AKT [56]. Also, it was shown that in postmortem AD brains the level of phosphorylated mTOR elevated which was related the inhibition of autophagy and increase in tau hyperphosphorylation [57], in our study SIRT1 activator did not altered the mTOR level while SIRT1 inhibitor decreased the phosphorylated mTOR level. This result may suggest that SIRT1 inhibitor might activate autophagy to help Aβ clearance. On the contrary SIRT1 activator did not provide increase in the Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
level of phosphorylated AKT and did not cause GSK3 inhibition. However, when compared to amyloid beta group, it increased the PTEN level. Majority of the studies shows that the level of PTEN decreased in postmortem brain tissues of AD patients [58, 59, 60, 61], therefore SIRT1 activator might show protective effect through PTEN.
Exosome releasing in AD pathology is a debating issue among researchers. Certain scientist argued that exosome release is a mechanism which helps clearing of toxic proteins while others decline that exosomes causes toxic protein propagation between cells [26, 34, 35]. In the present study the amount of exosomes decreased in response to amyloid beta toxicity. However, the exosome releasing amount increased significantly with SIRT1 activator treatment compared to both control and amyloid beta groups. Exosome releasing amount decreased compared to the control, when the inhibitor was given alone and as therapeutically. However, when compared to the amyloid beta group the amount of exosome increased in inhibitor groups. In the present study, only GSK3α/β was detected in exosomal content and only the expression and activation of GSK3β was altered with Aβ toxicity. In our results, SIRT1 inhibitor caused GSK3α and GSK3β inhibiton in the exosomes which is parallel to the cell lysate results. Interestingly SIRT1 activator also caused GSK3β inhibition. Therefore, both SIRT1 activator and inhibitor produced same neuroprotective effect in exosomal content. One of the previous studies showed that the exosomes carrying a therapeutic molecule can treat AD symptoms through activating the AKT/GSK3β pathway in which both the level of phosphorylated AKT and the level phosphorylated GSK3β increased [62]. The pharmacological modulation of GSK3β by SIRT1 activator and inhibitor have shown promise in the treatment of AD by increasing exosome amount and therefore toxic protein clearance and by increasing phosphorylation levels of the GSK3β when compared to amyloid beta group.
Conclusion
This study was done to detail the molecular mechanism of SIRT1 activator and inhibitor effects on in- vitro amyloid beta toxicity model. When SIRT1 was stimulated by activator CAY10602, the antioxidant status was ameliorated and also activator shown potential protective effect by increasing PTEN amount. When inhibitor Ex527 was applied to the cells, it altered the phosphorylated levels of AKT, GSK3 and mTOR in the opposite way of AD. In the exosomal content, it was observed that both SIRT1 activator and inhibitor may have protective effect by increasing exosome amount and therefore toxic protein clearance and by inhibiting GSK3α and GSK3β, while that was parallel with the cell lysates results for the SIRT1 inhibitor.
Acknowledgment
The authors would like to thank Asst. Prof. Mehmet Ozansoy and M.Sc. Mehmet Ozgen Altıntas for their help in this study.
Funding
This research received funding from the Science Foundation BAP of the University of Health Sciences (BAP number 2017/055).
References
-
Polanco JC, Li C, Bodea LG, Martinez-Marmol R, et al. (2018) Amyloid-β and tau complexity Towards improved biomarkers and targeted therapies. Nat Rev Neurol 14(1): 22-40.
-
Chen GF, Xu TH, Yan Y, Zhou YR, Jiang Y, et al. (2017) Amyloid beta: Structure, biology and structure-based therapeutic development. Acta Pharmacol Sin 38(9): 1205-1235.
-
(2016) WHO Fact Sheets, The Top 10 Causes of Death, statistics.
-
Masters CL, Bateman R, Blennow K, Rowe CC, Sperling RA, et al. (2015) Alzheimer’s disease. Nat Rev Dis Prim 1(1): 1-18.
-
Meyer-Luehmann M, Stalder M, Herzig MC, Kaeser SA, Kohler E, et al. (2003) Extracellular amyloid formation and associated pathology in neural grafts. Nat Neurosci 6(4): 370-37.
-
Demuro A, Mina E, Kayed R, Milton SC, Parker I, et al. (2005) Calcium dysregulation and membrane disruption as a ubiquitous neurotoxic mechanism of soluble amyloid oligomers. J Biol Chem 280(17): 17294-17300.
-
Thathiah A, De Strooper B (2011) The role of G protein-coupled receptors in the pathology of Alzheimer’s disease. Nat Rev Neurosci 12(2): 73-87. Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
-
Calissano P, Matrone C, Amadoro G (2009) Apoptosis and in vitro Alzheimer’s disease neuronal models. Commun Integr Biol 2(2): 163-169.
-
Carolindah MN, Rosli R, Adam A, Nordin N (2013) An Overview of in Vitro Research Models for Alzheimer’S Disease (AD). Regen Res 2(2): 8-13.
-
Donmez G (2012) The neurobiology of sirtuins and their role in neurodegeneration. Trends Pharmacol Sci 33(9): 494-501.
-
Hubbard BP, Sinclair DA (2014) Small molecule SIRT1 activators for the treatment of aging and age- related diseases. Trends Pharmacol Sci 35(3): 146-154.
-
Donmez G (2012) The effects of SIRT1 on Alzheimer’s disease models. Int J Alzheimers Dis 2012: 4-6.
-
Lalla R, Donmez G (2013) The role of sirtuins in Alzheimer’s disease. Front Aging Neurosci 5: 1-4.
-
Carafa V, Nebbioso A, Altucci L (2012) Sirtuins and disease: The road ahead. Front Pharmacol 3: 1-6.
-
Qin W, Yang T, Ho L, Zhao Z, Wang J, et al. (2006) Neuronal SIRT1activation as a novel mechanism underlying the prevention of alzheimer disease amyloid neuropathology by calorie restriction. J Biol Chem 281(31): 21745-21754.
-
Lavu S, Boss O, Elliott PJ, Lambert PD (2008) Sirtuins - Novel therapeutic targets to treat age-associated diseases. Nat Rev Drug Discov 7(10): 841-853.
-
Bonda DJ, Lee HG, Camins A, Pallas M, Casadesus G, et al. (2011) The sirtuin pathway in ageing and Alzheimer disease: Mechanistic and therapeutic considerations. Lancet Neurol 10(3): 275- 279.
-
Ng F, Wijaya L, Tang BL (2015) SIRT1 in the brain- connections with aging-associated disorders and lifespan. Front Cell Neurosci 9: 1-13.
-
Green KN, Steffan JS, Martinez-Coria H, Sun X, Schreiber SS, et al. (2008) Nicotinamide Restores Cognition in Alzheimer’s Disease Transgenic Mice via a Mechanism Involving Sirtuin Inhibition and Selective Reduction of Thr231-Phosphotau. J Neurosci 28(45): 11500-11510.
-
Thery C, Zitvogel L, Amigorena S (2002) Exosomes: composition, biogenesis and function. Nat Rev Immunol 2(8): 569-579.
-
Fevrier B, Raposo G (2004) Exosomes: Endosomal- derived vesicles shipping extracellular messages. Curr Opin Cell Biol 16(4): 415-421.
-
Sarko DK, McKinney CE (2017) Exosomes: Origins and therapeutic potential for neurodegenerative disease. Front Neurosci 11: 1-7.
-
Bellingham SA, Guo B, Coleman B, Hill AF (2012) Exosomes: vehicles for the transfer of toxic proteins associated with neurodegenerative diseases? Front Physiol 3: 1-12.
-
Chia BS, Low YP, Wang Q, Li P, Gao Z (2017) Advances in exosome quantification techniques. TrAC, Trends Anal Chem 86: 93-10.
-
Properzi F, Ferroni E, Poleggi A, Vinci R (2015) The regulation of exosome function in the CNS: implications for neurodegeneration. Swiss Med Wkly 145:1-14.
-
Malm T, Loppi S, Kanninen KM (2016) Exosomes in Alzheimer’s disease. Neurochem Int 97: 193-199.
-
Yuyama K, Sun H, Usuki S, Sakai S, Hanamatsu H, et al. (2015) A potential function for neuronal exosomes: Sequestering intracerebral amyloid‐β peptide. FEBS Lett 589(1): 84-88.
-
Winston CN, Goetzlb EJ, Akersd JC, Carterd BS, Rockensteina EM, et al. (2016) Prediction of conversion from mild cognitive impairment to dementia with neuronally derived blood exosome protein profile. Alzheimers Dement (Amst) 3: 63-72.
-
Polanco JC, Scicluna BJ, Hill AF, Gotz J (2016) Extracellular vesicles isolated from brains of rTg4510 mice seed tau aggregation in a threshold-dependent manner. J Biol Chem 291(24): 12445-12446.
-
Howitt J, Hill AF (2016) Exosomes in the pathology of neurodegenerative diseases. J Biol Chem 291(52): 26589-26597.
-
Yuyama K, Sun H, Mitsutake S, Igarashi Y (2012) Sphingolipid-modulated exosome secretion promotes clearance of amyloid-β by microglia. J Biol Chem 287(14): 10977-10989.
-
Goetzl EJ, Boxer A, Schwartz JB, Abner EL, Petersen RC, et al. (2015) Low neural exosomal levels of cellular Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175. survival factors in Alzheimer's disease. Ann Clin Transl Neurol 2(7): 769-773.
-
Fiandaca MS, Kapogiannis D, Mapstone M, Boxer A, Eitan E, et al. (2015) Identification of preclinical Alzheimer's disease by a profile of pathogenic proteins in neurally derived blood exosomes: a case-control study. Alzheimers Dement 11(6): 600-607.
-
Abdullah M, Takase H, Nunome M, Enomoto H, Ito J (2016) Amyloid-B Reduces Exosome Release from Astrocytes by Enhancing JNK Phosphorylation. J Alzheimer Dis 53(4): 1433-1441.
-
Dinkins MB, Dasgupta S, Wang G, Zhu G, He Q, et al. (2015) The 5XFAD Mouse Model of Alzheimer’ s Disease Exhibits an Age-Dependent Increase in Anti- Ceramide IgG and Exogenous Administration of Ceramide Further Increases Anti-Ceramide Titers and Amyloid Plaque Burden. J Alzheimer Dis 46(1): 55-61.
-
Nayagam VM, Wang X, Tan YC, Poulsen A, Goh KC, et al. (2006) SIRT1 modulating compounds from high- throughput screening as anti-inflammatory and insulin-sensitizing agents. J Biomol Screen 11(8): 959- 967.
-
Shah SA, Yoon Gh, Chung SS, Abid MN, Kim TH, et al. (2017) Novel osmotin inhibits SREBP2 via the AdipoR1/AMPK/SIRT1 pathway to improve Alzheimer’s disease neuropathological deficits. Mol Psychiatry 22(3): 407-416.
-
Mattson MP (2004) Pathways Towards and Away from Alzheimer’s Disease. Nature 430(7000) 631- 639.
-
Perry G, Cash AD, Smith MA (2002) Alzheimer disease and oxidative stress. J Biomed Biotechnol 2(3): 120- 123.
-
Huang H, Yan P, Sun T, Mo X, Yin J, et al. (2018) Procyanidins extracted from lotus seedpod ameliorate amyloid-β-induced toxicity in rat pheochromocytoma cells. Oxid Med Cell Longev 2018: 1-14.
-
Carrillo-Mora P, Luna R, Colín-Barenque L (2014) Amyloid beta: Multiple mechanisms of toxicity and only some protective effects? Oxid Med Cell Longev 2014: 1-15.
-
Atwood CS, Obrenovich ME, Liu T, Chan H, Perry G, et al. (2003) Amyloid-β: A chameleon walking in two worlds: A review of the trophic and toxic properties of amyloid-β. Brain Res Rev 43(1): 1-16.
-
Quadros BAQ, Silva JPS, Romeiro CFR, dos Santos SV, Rodrigues CA, et al. (2018) Neuroprotective mechanisms of resveratrol in Alzheimer’s disease: Role of SIRT1. Oxid Med Cell Longev 2018: 1-15.
-
Albani D, Polito L, Batelli S, De Mauro S, Fracasso C, et al.(2009) The SIRT1 activator resveratrol protects SK- N-BE cells from oxidative stress and against toxicity caused by α-synuclein or amyloid-β (1-42) peptide. J Neurochem 110(5): 1445- 1456.
-
Lutz MI, Milenkovic I, Regelsberger G, Kovacs GG (2014) Distinct patterns of sirtuin expression during progression of Alzheimer’s disease. Neuromolecular Med 16(2): 405-414.
-
Min SW, Cho SH, Zhou Y, Schroeder S, Haroutunian V, et al. (2011) Acetylation of Tau Inhibits Its Degradation and Contributes to Tauopathy. Neuron 67(6): 953-966.
-
Chen J, Zhou Y, Mueller-Steiner S, Chen LF, Kwon H, et al. (2005) SIRT1 protects against microglia-dependent amyloid-β toxicity through inhibiting NF-κB signaling. J Biol Chem 280(48): 40364-40374.
-
Solomon JM, Pasupuleti R, Xu L, McDonagh T, Curtis R, et al. (2006) Inhibition of SIRT1 Catalytic Activity Increases p53 Acetylation but Does Not Alter Cell Survival following DNA Damage Inhibition of SIRT1 Catalytic Activity Increases p53 Acetylation but Does Not Alter Cell Survival following DNA Damage. Mol Cell Biol 26(1): 28-38.
-
Oon CE, Strell C, Yeong KY, Ostman A, Prakash J (2015) SIRT1 inhibition in pancreatic cancer models: Contrasting effects _in vitro_ and _in vivo_. Eur J Pharmacol 757: 59-67.
-
Dai H, Perni RB, Kustigian L, Carney D, Case A, et al. (2010) SIRT1 activation by small molecules: Kinetic and biophysical evidence for direct interaction of enzyme and activator. J Biol Chem 285(43): 32695- 32703.
-
Yi JH, Baek SJ, Heo S, Park HJ, Kwon H, et al. (2018) Direct pharmacological Akt activation rescues Alzheimer’s disease like memory impairments and aberrant synaptic plasticity. Neuropharmacology 128(2018): 282-292. Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175.
-
Zhang B, Wang Y, Li H, Xiong R, Zhao Z, et al. (2016) Neuroprotective effects of salidroside through PI3K/Akt pathway activation. Drug Des Devel Ther 2016(10): 1335-1343.
-
Baki L, Shioi J, Wen P, Shao Z, Schwarzman A, et al. (2004) PS1 activates PI3K thus inhibiting GSK-3 activity and tau overphosphorylation: Effects of FAD mutations. EMBO J 23(13): 2586-2596.
-
Phiel CJ, Wilson CA, Lee VM, Klein PS (2003) GSK- 3a regulates production of Alzheimer’s disease amyloid-b peptides. Nature 423(6938): 435-439.
-
Jimenez S, Torres M, Vizuete M, Sanchez-Varo R, Carmona-Cuenca I, et al. (2011) Age-dependent accumulation of soluble amyloid β (Aβ) oligomers reverses the neuroprotective effect of soluble amyloid precursor protein-α (sAPPα) by modulating phosphatidylinositol 3-kinase (PI3K)/Akt-GSK- 3β pathway in Alzheimer mouse model. J Biol Chem 286(21): 18414-18425.
-
Cai Z, Chen G, He W, Xiao M, Yan LJ (2015) Activation of mTOR: A culprit of Alzheimer’s disease? Neuropsychiatr Dis Treat 11: 1015-1030.
-
Tang Z, Ioja E, Bereczki E, Hultenby K, Li C, et al. (2015) mTor mediates tau localization and secretion: Implication for Alzheimer’s disease. Biochim Biophys Acta - Mol Cell Res 1853(7): 1646- 1657.
-
Tramutola A, Triplett JC, Domenico FD, Niedowicz DM, Murphy MP, et al. (2015) Alteration of mTOR signaling occurs early in the progression of Alzheimer disease (AD): Analysis of brain from subjects with pre-clinical AD, amnestic mild cognitive impairment and late-stage AD. J Neurochem 133(5): 739-749.
-
Matsuda S, Nakagawa Y, Tsuji A, Kitagishi Y, Nakanishi A, et al. (2018) Implications of PI3K/AKT/PTEN Signaling on Superoxide Dismutases Expression and in the Pathogenesis of Alzheimer’s Disease. Diseases (Basel, Switzerland) 6(2): 1-13.
-
O’Neill C (2013) PI3-kinase/Akt/mTOR signaling: Impaired on/off switches in aging, cognitive decline and Alzheimer’s disease. Exp Gerontol 48(7): 647-653.
-
Heras-sandoval D, Perez-rojas JM, Hernandez-damian J, Pedraza-chaverri J (2014) The role of PI3K/AKT/mTOR pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration. Cell Signal 26(12): 2694-2701.
-
Wang H, Sui H, Zheng Y, Jiang Y, Shi Y, et al.(2019) Curcumin-primed exosomes potently ameliorate Kilic U, et al. Both Activation and Inhibition of SIRT1 May Act via Exosomal GSK3α/βin the in-vitro Amyloid-beta Toxicity Model. Adv Clin Toxicol 2019, 4(4): 000175. cognitive function in AD mice by inhibiting hyperphosphorylation of the Tau protein through the AKT/GSK-3β pathway. Nanoscale 11(15): 7481-7496.
- Evaluation of Skin Aging Preventive Effects of Cherry Blossom Petal Extracts Through Antioxidant and Anti-Glycation Activities
- Is Cell Death Responsible for False Positive Results of In Vivo Comet Assay?
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism