Interactive Correlation between Exosomes and Nanomaterials
The extracellular vesicular entities, also called as plasma dusts, are present in all biological fluids, cell lines and cultures, are fascinating the researchers. Investigations related to their structure, formation, biological, physiological, and cellular status reveal that exosomes are biostable, and morphologically resemble nanomaterials, specifically, those from mesenchymal stem-cells. As the exosomes are multi-utility cellular products, their cellular yield and quantification seems to be tedious. Exosomes as nanostructures enhance the efficacies of extracellular vesicles or exosomes by fusing with lateral endosomes or multivesicular bodies, and later bud off from plasma membrane in a similar manner as during endocytosis. These cellular vesicles are the functional backbone of most of inter and intracellular transport mechanisms. It becomes imperative to understand their characterization, factors affecting their behavior within and outside the cell. Ubiquitously, nanomaterials are used in biological, medical, pharmaceutical, and biomolecular fields. The combined use of exosomes and nanomaterials may act as useful tools for clinical and diagnostic applications as they reflect the physiological and pathological status of a system. The molecular crowding is a physiological process and controls dissipation of molecular structures that facilitate the effective functions, and determination of cellular physiochemical status. Therefore, it essentiates to appraise the implications of exosomes along with nanomaterials in relation to cellular, biomolecular, physicochemical aspects of interactions and their applications in the biomedical fields. In this review, an effort is made to explore the mechanism of their biogenesis, exosomes functions in association with nanomaterials, molecular crowding, and their structure and functional relationship
Overview
Professor Wolf noticed extracellular vesicles in plasma and named them as “plasma dust” in 1967. Later, these vesicular entities were detected in all biological body fluids and the cell lines and cultures. The exosomes are the products of cellular vesicles; this process is similar to endocytosis [1]. These membrane bound and membrane derived vesicles are the cellular product and mammalian cells like immune cells like macrophages, dendritic cells, T-cells, B-cells, mesenchymal stem cells, epithelial, endothelial cells, and cancer cells. Other cells such as intestinal epithelial cells, fibroblast, and antigen presenting cells, platelets, hepatocytes, and lymphocytes also produce exosomes (Figure 1). One can observe them in body fluids like human saliva, serum, breast milk, urine and even in semen [2, 3, 4, 5].
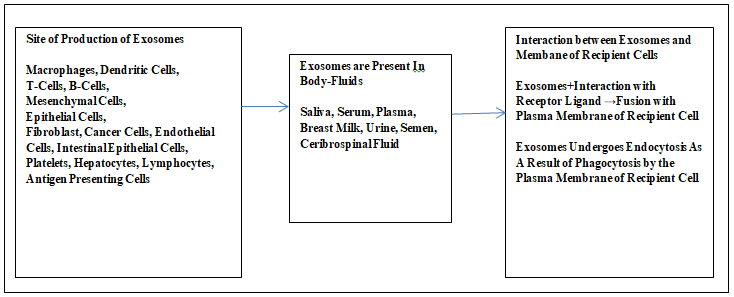
Exosomes are extracellular vesicular bodies that are delivered by the cell; before their release these fuse with the intermediate endocytic compartment and multivesicular entity involving the plasma membrane.
There exist microvesicles related to apoptosis (apoptotic bodies). These are the derivatives of the plasma membrane and separate as a result of shedding from it. The ectosomes, exosomes, and apoptotic bodies exhibit a similar range of sizes typically within 40-100 nm. These structures gulp some of the cytosol [6]. The exosomes are present in the body fluids like blood, urine, cerebrospinal fluid, and the matrix of the tissues of vertebrates and other multicellular organisms. The exosomes are also present in the matrix of the tissue and these are Matrix Bound Nanovesicles (MBV). The size of exosomes depends on the parent matrix-bound nanovesicles. Most of the exosomes exhibit a smaller size than the extracellular vesicles ranging within 30 to 150 nm diameters. The size of lipoproteins is much less than the cell but it is similar to that of the most extracellular vesicles [7, 8, 9, 10].
Endosomes are the primary molecular players within the cell and participate in cellular communication, growth, regulatory activities, defense, normal functionalities of cell, and pathogenesis. The endosomes are the integral component of the classical endocytic pathway that results in the formation of late endosomes and/or multivesicular cell organelles and exosomes (endosomes when are outside the cell are exosome). This process exhibits a well-organized transformation of such dynamic cellular structures in which their disconnection occurs from the parent membrane and finally becomes spatial and functional components of the feeder lysosomal pathway [11]. The exosomes play many roles like carrying waste or garbage of cells, biologically active materials, and exhibit immunomodulatory features. These take part in endocytic membrane transport pathway involving endosomes within the Trans Golgi network. This pathway transports the internalized molecules or ligands through the plasma membrane up to lysosomes. The lysosomes are the site where the transported molecules are degraded and or reprocessed and put back in the endocytic cycle or Trans Golgi network or Golgi apparatus within the plasma membrane. There is a heterogeneous group of vesicles which include exosomes and microvesicles. These vesicles originate from the endosomal system and exist in the biological body fluids of the biosystem and play significant physiological roles in the pathogenesis [12].
The exosomes are the membrane bound extracellular vesicles and act as intracellular sorting organelles in eukaryotic cells and these entities contain cellular or unwanted mater to be expelled out of the cell [6, 13, 14]. The exosomes participate in the normal cellular physiological functions including immune response, presentation of antigen, signal transduction, prognostic information during pathogenesis like chronic inflammation, diseases based on lipid metabolism, neurodegenerative disorders, cardiovascular, renal disorders, and tumor [15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25]. The exosomes are the products from different types of cells but exhibit basic common structural and functional proteins.
These proteins are Rab GTPase, SNARESs, annexins, Alix, Tsg101 and tetraspanins (Figure 2).
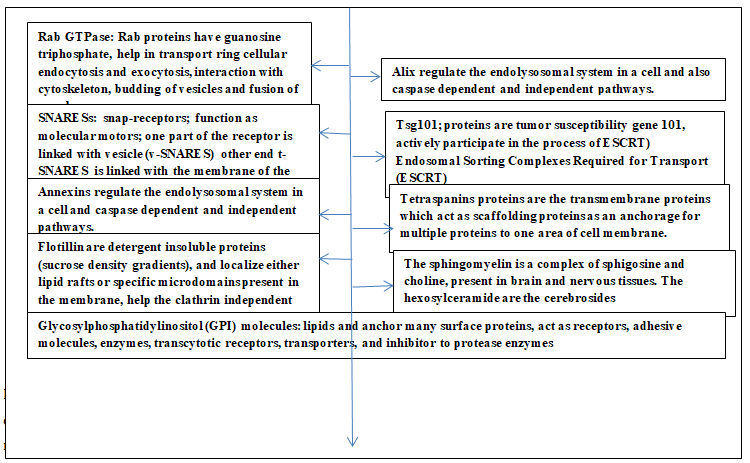
Rab proteins are guanosine triphosphatases and bring about protein transport in cellular endocytic and exocytic processes. These proteins also help the processes like interaction with cytoskeleton, fusion of membrane and budding of vesicles. SNAREs are the snap-receptors present on the membrane and function as molecular motors; these enable the required fusion between the two membranes. One part of this protein is located on the vesicle membrane (v-SNAREs) while the corresponding part (t-SNAREs) is on the target membrane. The annexins regulate the endolysosomal system in a cell and also caspase dependent and independent pathways. The Alix proteins regulate the endolysosomal system in a cell and also caspase dependent and independent pathways. The Tsg101 proteins are tumor susceptibility gene 101, and actively participate in the process of ‘Endosomal Sorting Complexes Required for Transport (ESCRT)’. The tetraspanins proteins are the transmembrane proteins which act as scaffolding proteins as an anchorage for multiple proteins to one area of cell membrane. Other biomolecules present are glycosylphosphatidylinositol- anchored molecules, flotillin, cholesterol, sphingomyelin and hexosylceramides. The glycosylphosphatidylinositol (GPI) molecules are lipids that anchor many surface proteins, and also act as receptors, adhesive molecules, enzymes, transcytotic receptors, transporters, and inhibitor to protease enzymes. The flotillin are detergent insoluble proteins (sucrose density gradients), and localize either lipid rafts or specific microdomains present in the membrane. These help the clathrin independent endocytosis pathway, the Src-family tyrosine kinase regulate this pathway. The sphingomyelin is a complex of sphigosine and choline, present in brain and nervous tissues. The hexosylceramide are the cerebrosides. The ATP-mediated activation of purinergic, receptors, activation of thrombin, and lipopolysaccharides, elevated intracellular Ca++ concentration, stimulate the release of extracellular vesicles [23, 24, 25, 26, 27, 28, 29, 30]. The exosomes are nanosized entities bounded with lipid bilayer ensheathing a fluid core and these components readily integrate with hydrophilic and hydrophobic compounds. As a result, the exosomes act as suitable cargo vehicles for genetic matter, proteins, and drugs. These exosomes carry their cargos to the specific target using one of the modes among interaction between receptor-ligand, endocytosis during phagocytosis, and direct fusion between exosomes and plasma membrane of recipient cell that enables the release of cargo into the cytoplasm of the target cell [5]. Further, these nanosized biocompatible entities are suitable theranostics, imaging probes, targeting ligands and in forming covalent linkage [31]. Due to some of the above features the emerging role of exosomes suggests their clinical implications as these are one of the most suitable, biocompatible, and biodispersible entities unlike some of the synthesized/designed structures which have toxicity and biocompatible issues [6, 32].
Characterization of Exosomes
Investigations related to the study, separation, and behavior of exosomes present in the biological fluids like blood, cerebrospinal fluid, breast milk, urine, amniotic fluid, seminal fluid, saliva, and malignant effusion, involve the physical and biophysical features of exosomes. The exosomes exhibit physical and biological features like size, surface charge, density, the cargo they carry, and membrane- associated antigen. Some of the popular techniques for their investigation include electron microscopy, dynamic light scattering, tunable resistive pulse sensing, and nanoparticle tracking analysis. The zeta potential of exosomes indicates the charge on its surface and acts as a significant parameter during the investigations concerning their dispersion in the emulsion, suspension, and colloidal nature in the testing samples. The zeta potential also indicates measuring the charge stability and influences the interactions of the affected particles. The exosomes attain surface electric charge spontaneously when they come in contact with a polar medium like a hydrophilic buffer. The surface charge is negative on exosomes as one finds on the plasma membrane of a cell specifically in hydrophilic buffers. The electron affinity differences between two phases, ionization of exosome membrane surface groups, and differential ion absorption due to the electrolytes solutions [33, 34, 35].
The Biochemical Aspects of Endosomal Membrane
It is very essential to review the biochemical and biophysical aspects of the endosomal membrane because this membrane is the structural and functional origin of exosomes. This role of the endodermal membrane is crucial during the structural and compositional changes that occur when the exosomes are developing and maturing. The biophysical properties and the protein and lipid components play a significant role during the endocytic and biosynthesis of cell organelles. Among these organelles, exosomes are of cellular functional importance because these entities are the derivatives of the endosomal membrane. Primarily, phosphatidylcholine and lysobisphosphatidic acid are present in the endosomal membrane and contribute to its microheterogeneity. The lysobisphosphatidic acid manifests strong fusogenic features that are pH-dependent and functional dynamic nature of the endosomal membrane. The morphological and biophysical investigations reflect the presence of a fluid region rich in glycerophospholipid within the bilayer and a liquid region which is plentiful with cholesterol and glycosphingolipids. This region is the site of the preferential portion of glycosylphosphatidylinositol- anchored proteins and double-acylated proteins. Cholesterol, glycosphingolipids, and glycosylphosphatidylinositol (GPI) anchored proteins constitute raft-lipid microdomains; these are not soluble in lower concentrations of non-ionic detergents. The lipid rafts are among the components that behave as cell surface platforms and act as functional bases during internalization or uptake, signaling, and pathogenic infection. These also help in sorting out proteins during biosynthetic processes [36]. The biomolecules participating in the endocytic pathway internalize during the early phase of development and later either recycle at the plasma membrane or degrade at lysosomes. There is a possibility that most of the participant biomolecules distribute in a mosaic pattern but some lipids do not follow this mode. The lipid rafts are higher in number among recycling endosomes, and as per the preference lipid analog shift to the late endosomes. The phosphatidylinositol 3-phosphate interacts with those proteins which contain FYVE or a PX domain, and these are prevalent during the early phase of the endocytic pathway. [The FYVE is a zinc-finger domain and includes proteins like Fab-1, YOTB, and EEA-1; these are cystic proteins and readily bind phosphatidylinositol 3-phosphate. The metal ion present in the domain plays a significant role while and coordinating and basic amino acids [37]. The PX domain binds with the phosphoinositide-binding component domain which targets cell membrane proteins. This domain is present in the P40phox and p74phox domain of the NADPH oxidase enzyme [38]. The lysobisphosphatidic acid is present relatively in a higher amount during the late endosomal phase. The quantity of lysobisphosphatidic acid is also more within the internal membrane of multivesicular organelles and multiwalled cellular structures [36, 39, 40, 41].
The Biophysical Aspects of Endosomal Membrane and Formation of Exosomes
The biological membrane, non-lamellar lipid phases, and lipid vesicles have a specific bending elasticity. This means curvature moduli (Kc) of lipid bilayers, the monolayer of a leaflet of inverted hexagonal phases of lipids, and the Gaussian curvature mode play a significant role during the formation of endosome. The endocytic pathway is an important biophysical functional aspect of cellular functions and provides inputs for intracellular trafficking. These inputs facilitate the internalization of nutrients, compartmentalization, and termination of signaling processes. The biochemical complexity and physical or biophysical forces result in the formation of an endocytic pit (initiation of formation of endosome). This process includes asymmetrical trans-bilayer stress, scaffolding, line tension, crowding, and passive mechanism. The active mechanism related to mechanochemical enzymes and cytoskeleton also participates during the formation of endosomes from the endosomal membrane [42].
Every biological membrane exhibits biomechanical characteristics and required energy that results in its bending. Bending is one of the prime processes during the formation of the endosomal membrane. The mode of bending in the case of the biological membrane depends on its biochemical aspects and its lipids composition. A biomembrane shows a lower degree of bending when unsaturated acyl chains are present in the lipids. The membranes having lipids with saturated acyl chains show a higher degree of bending; the amount of cholesterol present elevates the degree of bending in a given membrane. In the case of the fluid membrane the range of bending lies between 10 to 100Kt; k represents Boltzmann constant and T represents absolute temperature. [kT = 4X¯21 N/m at room temperature. The factor-like capacity of the lipid membrane is an important aspect in deciding the trafficking via biomembrane and this process needs energy. This required energy depends on the mode of bending rigidity (k) of the membrane and is based on the characteristic composition of the concerned membrane. The curvature and phase transition act as one of the important parameters during the sorting of lipids and fission of membrane tubules. The tubes and vesicles formed in a ternary mixture having sphingomyelin, phosphatidylcholine, and cholesterol show two clear significant difference in lipid composition in two nanostructures formed. The concentration and the temperature affect the formation of these nanostructures in this ternary mixture. The phase separation takes place during the tube fission. This feature reflects on the occurrences of lipid sorting at the time of tube fission and also this phenomenon critically relates to the membrane curvature and phase separation. The process of phase separation is significant during the membrane fission in tubes pulled out of the huge liposomes and/or purified membranes of the Golgi Complex [43, 44]. A lipid bilayer having both similar leaflets shows flat geometry when and when this symmetry becomes asymmetrical, the bending of the membrane takes place. Many different lipids are present on the membrane, and many proteins can be introduced during the state of asymmetry and this introduction causes an asymmetrical state in a given biological membrane.
Let, C represents an imposed curvature, C0 is an immediate membrane curvature; (it indicates the relaxed equilibrium curvature caused on the membrane specifically when external mechanical action is absent), and k is the energy following the mode of bending rigidity. Then Fbending =k/2 X (C-Co)2 [45]. Biological and biomimetic membranes have a fluid nature, and show bending and stretching elastic deformations. The elastic parameters like spontaneous (or unplanned) and selected curvature (m) and its bending rigidity (k) regulate the membrane behavior. Both these variables are under the intrinsic tension scale. The factor 2km2 represents the spontaneous (unplanned) tension. The mechanical tension causes stretching and compression to a membrane. This behavior of the membrane is helpful in preparation of experimental model of membrane, membrane biogenesis and related studies.
In the case of the membrane of the vesicle, the elastic distortion relates to the enclosed volume even if the mechanical forces are absent and the combined bending and stretching energy are the minima with the area of membrane for the fixed volume of the vesicle. These functional conditions cause mechanical tension within a membrane of the vesicle which is proportional to the spontaneous curvature and bending rigidity of the membrane. The factors like morphology of vesicle and the magnitude of mechanical tension may be very less as compared to the spontaneous tensions [46]. The surface density of curvature-generating proteins plays a significant role when spontaneous curvature results in a biological membrane. The protein having Bin/Amphiphysin/Rvs domains regulate the formation of curvature in a lipid membrane. These proteins are also functionally essential during cellular functions like endocytosis, cellular trafficking, motility, and other cellular functions that sustain cellular survival. These proteins also constitute three-dimensional structures of 3D molecular scaffolds that facilitate change in the mechanical features and reshaping of the membrane [47]. Conventionally, the invagination of membrane takes place towards proteins that facilitate curvature during positive curvature, and when invagination occurs away from curvature generating proteins it results in the negative curvature [46]. The direction of membrane deformation is of functional significance in the biological field. Generally, whenever cytosolic protein interacts with the cytoplasmic leaflet resulting in the membrane bending in the direction of cytosol; it causes positive curvature. When the protein binds with exoplasmic leaflet inducing curvature towards cytosol it results in negative curvature. A good number of cytosolic proteins bind with bilayers and form any type of curvature. Most of the pathogens utilize the negative curvature at the site of the entrance on the plasma membrane and also during their budding out of the host cells. Generally, toxins interact with glycosphingolipids which cause negative curvature. The protein-lipid complexes like these have a cone geometrical shape and this shape results in compaction involving lipid head groups. The acyl chain in the presence of toxin-bound lipid head groups creates mechanical strain and the bending minimizes this strain [48, 49, 50, 51]. This feature possibly illustrates the mechanism during the physicochemical interactions between membrane and the interacting toxins.
The mechanism of membrane bending may be symmetric or asymmetric, and involve phenomena like chirality, scaffolding, and crowding. The asymmetric trans-bilayer stress and lipid or protein compaction act as structural and functional bases during the bending mechanism of a membrane. There may be an asymmetric insertion of the helix or asymmetric transmembrane proteins into one of the leaflets of the membrane. There may be either lipid or protein compaction into the membrane. When amphipathic or hydrophobic proteins interpose from one side of the membrane there forms a bend. The parameters like shape and density of protein on the membrane and the depth of interposition determine the overall interposition. When there is a low density of protein in a given membrane the fractional density of bound or inserted protein influence the spontaneous membrane curvature. The type and pattern of distribution cluster of transmembrane proteins also act as a deciding parameter in the spontaneous membrane curvature bud [52, 53, 54].
The chirality among molecules like membrane components, proteins, and lipids specifically glycolipids represents the intrinsic asymmetry and this feature can cause a deformity in the membrane. The topological defects occur whenever chiral molecules get involved as structural components and cause constrain in a uniform membrane.
The formation of tubules or buds release such constrain in a constrained membrane. The toxins released by cholera incept abnormal lipid phase as a result multiple bindings take place with glycosphingolipid GM1, the resultant structure is the textured phase. The proteins attached to glycolipids or glycophosphatidylinositol (GP1) are likely to induce new structural orientation in the membrane, and this change is probably because of their coupling with the dynamic cortical cytoskeleton. The mechanism of this process appears to be ambiguous, and possibly the transmembrane proteins are likely to associate with the mechanism occurring utilizing cytosolic tails [55, 56].
The process of scaffolding facilitates the molecular interactions that take place during cellular functions structurally. Scaffolds, in molecular biology, are the specific biomolecular frameworks that help to assemble to help desirable cellular interaction/s so that results in new functional structures, and the process is scaffolding. This process occurs within cells and recruits varieties of adaptor proteins, some factors, kinases, phosphates, and enzymes. The rigid and curved backbone of the membrane participates in formation of spontaneous membrane curve. The localized deformation on the membrane causes curvature tension between the surface of the protein and the membrane. This change acts as a scaffold and it is appropriate for developing membrane curvature [57].
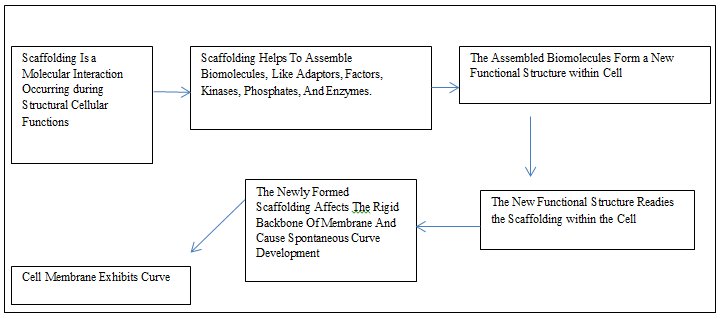
The BAR-domain protein belongs to a superfamily whose members have a backbone consisting of dimers that have different intrinsic curvature and a specific orientation; these features depend on the specific member of the family. The N-BAR proteins possess amphipathic helices and BAR- domains. Both of these components and their density at the site on the membrane affect the formation of curvature. The BAR domain superfamily negotiates the biogenesis of cell organelles, cellular division, migration, and cellular trafficking [58]. The scaffold-based molecular models form new desired substructure or scaffolds. This phenomenon involves sequentially adding or arranging new atoms and bonds following the mold of the scaffold. These scaffolds exist in naturally generated molecules. The molecules produced involving scaffold influence the properties present and revamp the set features [59]. This aspect may explain the mechanisms involved in cell migration and organogenesis during embryonic development.
In general, the actin mechanism contributes to the bending and vesicle formation from the endocytic membrane. The bending of the membrane initiates when the actin network gets physically connected to the plasma membrane, at this time the cross-linking should be optimum. The nucleation and polymerization of the recently formed actin filaments derive the initial invagination of the membrane. When the rate of nucleation of filament declines the key transition takes place. Thereafter, the invagination grows and cleavage of vesicle progresses even if the proportional elevation of actin is not optimum. The amount of filamentous actin and the degree of cross-linking both these factors affect the growth of invagination and vesicle [60]. The processes of budding and tubulation in the membrane occur in the absence of the processes like scaffolding and bilayer insertion in the membrane involved. The spontaneous bending in the membrane is the result of lateral pressure induced by the steric interactions between the involved proteins; this lateral pressure is one of the impacts of molecular crowding [42, 61, 62].
The presence of some of the self-assembled proteins induce membrane curvature and form either spherical, tubular orientations. The proteins like clathrin, dynamin, F-BAR-domain proteins, come under this group of self- assembling proteins. The spherical assembled structures exhibit the radii range between 50 to 120 nm while tubular formations have 10 nm radii. These natural cytological formulations are regarded as coats. The cytological spherical structures show a strong tendency to self-assemble and this tendency is strongly associated with membrane resulting in the rigid structure; hence, these are assigned the term coat/s. The shape of these structures is following the degree of lateral pressure and the shape of the participating components. Generally, in human cells, the clathrin, COP-I, and COP-II give rise to spherical shape while dynamin and F-BAR-domain proteins attain tubular form [63, 64, 65, 66, 67]. The COP-I and COP-II are the protein complexes that help in the formation of walls of vesicles of the coats, hence, named as coatomers. These vesicles specifically carry proteins from the cis- end of the Golgi complex to the rough endoplasmic reticulum. The term COPI represents the specific coat protein complex and it inaugurates the formation of vesicles [68].
The local pulling force is other biophysical factor that plays functional role. The bending rigidity of a membrane and its lateral tension influence the local pulling force of a membrane. The amplitude of such forces varies within tens of pico-newtons. Such forces are common during the movements of molecular motors. The molecular motors associate with the membrane and these move along the cytoskeleton filaments; the microtubules couple with kinesins and myosins hang-around with actin [42].
The cell membrane and surface tension have correlation. The constituents of cell membrane like phospholipids, glycolipids and proteins play significant role in maintaining specific cell membrane surface tension. The phospholipids are plentiful in comparison to the proteins and glycolipids. The phospholipids result in the hydrophobic and hydrophilic interactions between tails and head respectively. These interactions cause a specific state of tension on the membrane. The in-plane membrane tension is stable along the complete cell membrane. The two factors of cell membrane, namely lipid composition and surface area, play major role in maintaining the specific membrane tension is a cell. Any stress or pull on the membrane (in-plane) disturbs the hydrophobic and hydrophilic interactions within the phospholipids of a membrane. A change in the in-plane membrane tension counter acts any deformation occurring in membrane. Any move to increase the area of cell membrane without changing the phospholipids results in elevation in membrane tension. When a cell is subjected to osmosis or hypertonic state, there is a swelling and increase in the surface area of the cell but no change in the phospholipid composition. The cell membrane interconnects with actin, a component of cytoskeleton present underneath which provide structural and functional support to the membrane (the inner surface of cell membrane) [69]. The surface tension of a membrane along with the adhesion between membrane and cytoskeleton maintain a specific level of cohesive force that retains the structural, spatial, and functional integrity of the membrane. A force or stretch or pull which is more than surface tension of a cell membrane causes deformation of a cell under investigation. There are many membrane domains having different molecular composition and properties because a cell membrane is made of different cholesterol, saturated and unsaturated phospholipids, and glycosylated lipids. These exhibit interaction resulting in ordered specific region on membrane and these regions utilize different lipids and proteins to carry out variety of functions. Thus, there are variety of domains on the cell membrane having different molecular composition and features. There exists phase separation phenomenon in the cell membrane due to lateral separation of lipids and this aspect relates with temperature, pressure, and structural properties like length of hydrocarbon tails, extent of unsaturation, and the constitution of the head of different lipids present in the membrane. The domain boundary or length of phase participates in the process of lipid phase separation, even at the rim of the membrane domain. This process gets the required energy for the steric interaction due to the high mismatch of phospholipids and it is related to the domain boundary or length of phase.
The line tension in a membrane is because of the interfacial energy present at the length of phase and or domain boundary in the membrane under consideration. The membrane domain attain circular orientation, this minimizes the degree of line tension. The decline in the free energy due to the formation of membrane domain recompenses the loss of free energy of line tension (if there is). Membrane tension (surface tension) and line tension provide structural and functional aspects in cellular activities like cellular motility, endocytosis, exocytosis, even during infection due to virus [69, 70, 71, 72, 73, 74, 75, 76]. The line tension is a force that causes bending or curves at specific locus; this happens due to misalignment or height mismatching. This morphological maladjustment exposes the hydrophobic acyl chain segment to bulk water molecule or hydrophobic head groups present in the ambient lipids in the membrane [77]. The bacterial toxins induce clustering of acyl chain glycosphingolipids resulting in the formation of domain which is more in size than the membrane [78]. The higher stretching force (lysis tension) ranging from 10ˉ2 to 10ˉ2 N/m is able to disrupt and cleave a lipid membrane. One can severe a transport carrier in the form of bud like or tube like structure from a biological membrane in a “non-leaky” manner either using squeezing or pinching mechanism [42, 79]. The protein dynamin or similarly structured protein is able to interchange the chemical energy of GTP (guanosine triphosphate) into the mechanical squeezing force on the membrane under study [80, 81, 82, 83]. The helical conformation of dynamin induces “local constriction” on a membrane. Further, thus local constriction enhances the elastic energy at the edge of the dynamin polymer which in turn diminishes the barrier of energy needed for fission [42, 83]. The membrane tension is a functional parameter that influences the membrane morphological maladjustment and this may reflect on the membrane tension experienced by the cell membrane of cells when subjected to cultural and testing experimental in cell-line culture.
General Structural and Functional Aspects of Exosomes
Exosomes are multi-vesicular bodies. The endocytic process helps the formation of exosomes in eukaryotic cells involving the endosomal membrane. A bilipid layer acts as its limiting membrane, endodermal in origin, and it is a derivative of the parent endoderm. These vesicular bodies are formed within the cell and exocytosis helps their release in the ambient environment. Some proteins, lipids, DNA, m-RNA, and mi-RNA from the parent cell get engulfed in these exosomes during their intracellular formation. The primary function of exosomes is to separate the recyclable or degradable to the sites like lysosomes. The multi- vesicular bodies are categorized depending on their sites of aggregation. Those which accumulate within the cytoplasm are “intraluminal vesicles” and the one released in the intercellular medium is “exosomes” [84, 85, 86, 87].
Generalized Mode of Formation of Exosomes Packaging of Crago
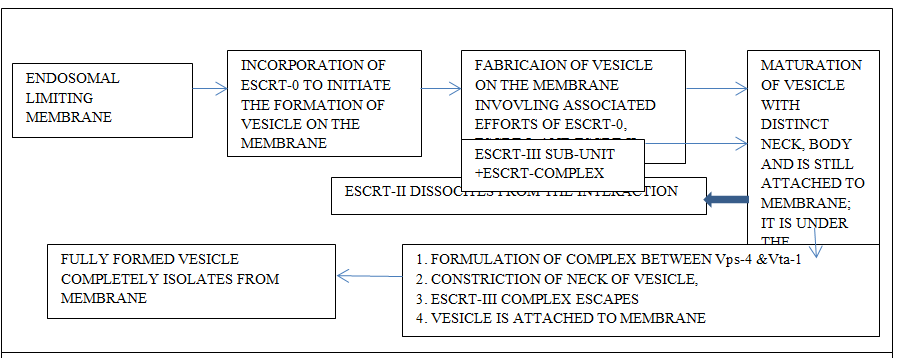
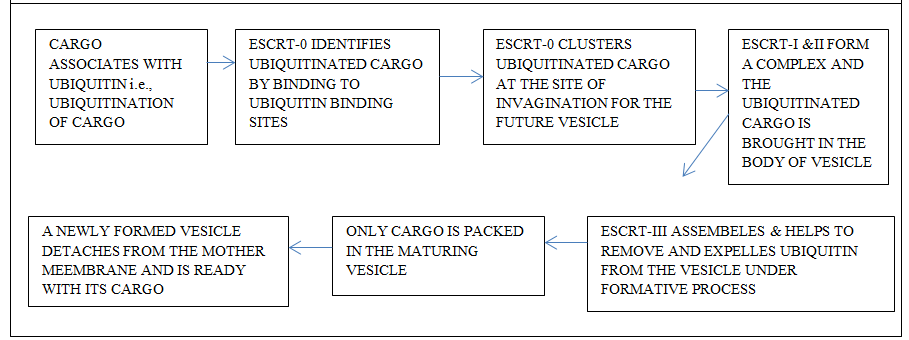
Most of the intraluminal vesicles develop from the endosomal limiting membrane. This process is either endosomal sorting complexes required for transport (ESCRT) mechanism dependent or occurs independently of endosomal sorting complexes required for transport mechanism [6]. There exists ambiguity concerning the mechanism of the production of exosomes [6]. The overall understanding reflects that the multi-vesicular bodies are the product of the intraluminal endosomal membrane. The vesicles develop as a bud and develop into a full vesicular body. Later, these fuse with lysosomes where their degradation takes place or later undergo exocytosis and come into the extracellular environment [84, 85, 86, 87]. Intraluminal vesicles are the products of the invagination of the endosomal membrane. The intraluminal vesicles aggregate within the lumen of matured endosomes. In due course of time, these provide the required metabolites which are needed for the biogenesis of melanosomes, Weibel-Palade bodies, and azurophilic particles; these structures are the lysosome concerned organelles. These products mingle with lysosomes or with the plasma membrane. The liberated intraluminal vesicles at this stage are exosomes [6].
The mechanism related to the endosomal sorting complex mode of formation of exosomes involves protein complexes. The endosomes engage these proteins after their cytosolic domains are tagged with ubiquitin. The ESCRT-O a component of endosomal sorting complexes required for transport (ESCRT) complexes identifies the ubiquitinated domain on the protein recruited by the endosomal membrane and conveys the ubiquitinated cargo onto ESCRT-1; its Tsg-101 of ESCRT-1 recognizes ubiquitin (Figures 4 & 5). Endosomal sorting complex (ESCRT) plays a key role in the formation of multi vesicular bodies and it also helps in loading the cargo. The ESCRT complex is of vital importance during retroviral budding and cytokinetic abscission. Basically this complex is essential during the recognition of cargo and membrane-sculpting device which show distinct behavior as ESCRT proteins, sorting of cargo, and the membrane they deform [88]. The involvement of ESCRT mode helps to conglomerate the ubiquitinated cargo proteins on the endosome. This also plays role in the formation of intraluminal vesicles by generating curvature of the endosomal membrane (Edgar 2016). It is of common observation that there is a formation of intraluminal vesicles even when endosomal sorting complexes required for transport (ESCRT) are absent and the process is independent of endosomal sorting complexes required for transport (ESCRT). The mechanism concerning this mode of formation of intraluminal vesicle needs more investigation. The basic understanding indicates that there is a need for a protein CD63; this protein is present in abundance on intraluminal vesicles but its significance is ambiguous. There seems to be a possibility that cone-shaped bending features of lipids like ceramide play some role in this process [89, 90, 91].
Exosomes act as Indicators of Pathogenesis
Most of the cells in human body and the general experimental models release the extracellular vesicles (exosomes), and these get appropriately biodistributed attributing to their specific functionalities. The biomolecules like tetraspanins, surface receptors, proteins, are present on the surface of exosomes. The exosomes derived from oligodendrocytes and those having ICAM-1 get readily internalized because of these biomolecules on the surface of exosomes. The exosomes produced by platelets, melanomal, and glioblastosomal cells conveniently fuse with the receptor cells specifically the targeted tumor cell; the acidic pH, intratumoral cellular physiological conditions further enhance such fusions [92, 93, 94]. Specifically, all types of nervous system produce extracellular vesicles (exosomes) during their normal developmental, physiological, and as behavioral responses to the pathogenic states. The composition of exosomes depends on the type and the physiological status of cells producing them. These exosomes display significant role while exhibiting phenotypic behavior and cellular fate [95]. The related investigations suggest that there are specific levels of these exosomes under normal and healthy statues of an individual. The normal levels of exosomes deviate in the body fluids during pathogenesis, progression of infection or disease, therapy success or failure, the systemic responses, and stressful conditions existing. Extracellular vesicles are a potential cellular products and that act as a possible biomarkers to diagnose a physiological and pathological conditions; more so is the case of neurological disorders [95, 96]. Intercellular communication is an important aspect while investigating functioning of nervous tissue pt nervous system, with respect to sensing behavior, synaptic communication, nerve regeneration, nerve degeneration and brain tumor or neuropathological conditions [96]. The presence of exosomes in biopsies samples are the probable biomarkers or indications that facilitates the clinical findings [96, 97, 98, 99]. It is very convenient to analyze exosomes as compared to circulating tumor cells, circulating free DNA, circulating free RNA, biopsies samples. This is because the exosomes are more in numbers, better intratumoral heterogeneity, more of molecular signature present in the related exosomes in comparison to the usual circulating cellular components. The exosomes enclose DNA and exhibit higher degree of sensitive and specificity regarding their detection because of the cargo, like KRAS (KARS gene provide instructions to prepare K-ras protein), EGFR (epidermal growth factor receptor), BRAF (BRAF gene mutation concerned with colorectal cancer and its spreading), and miRNA (microRNA), in exosome present in the biopsies fluid samples [99, 100, 101, 102]. The techniques like nanoparticles tracking analysis (NTA), flow cytometry, dynamic light scattering (DLS), Western blotting, immunoassay are some of the common modes to analyze the exosomes in a test samples. These techniques are helpful in analyzing presence of proteins, their molecular weight, quantification of surface proteins markers, concentration, phenotypes, size, of extracellular (exosomes) [99].
Interactive Interface between Exosomes and Nanoparticles
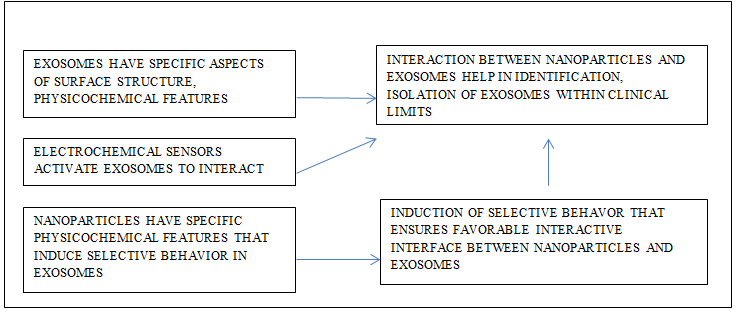
Exosomes exhibit specific surface markers; these markers interact with metal nanoparticles, thus, helping the detection and isolation of the exosomes from the test sample. These detections are within the level of clinical sensitivity. The electrochemical sensors are most suitable for this purpose. Specifically microfabricated microchip having multiplexed gold sensors is able to catch the exosomes and even microsomes during electrochemical analysis. The metal nanoparticles like silver, copper, nanoparticles successfully arrest epithelial cell adhesion molecules while copper nanoparticles capture prostate specific membrane antigen. Epithelial cell adhesion molecules are common markers found on epithelial cancerous cell/tissues. Such features of the exosomes reflect on their interactive behavior. This technique is fast, cost effective, and has higher level of clinical sensitivity [103, 104]. The physicochemical features of exosomes surface and nanomaterials exhibit selective behavior in response to the interaction between the two. There is selective uptake of exosomes associated with nanomaterials in tissues as it helps the regulated interaction between tissues and nanoparticles associated with exosomes. The exosomes are the cellular products that have antigens, proteins, and biomolecules that help their internalization in the target cells. This feature of exosomes accomplishes the cellular communication (Figure 4). The nanoparticles associated with exosomes readily escape the immunological recognition thereby enhancing the chances of degree of better biodistribution [105]. Thus, there seems to be favorable interactive interface between exosomes and nanomaterials. Metal-organic frameworks functionalized with extracellular vesicles elevate cargo loading efficiency, internalization, and biodistribution of the cargo in the test model and selective target homotypic tumor sites [106].
Carbon nanostructures such as graphene quantum dots, graphene oxide and functionalized as luminescent form are useful to detect biological molecules like microRNA, and short oligonucleotides. This detection is based on the specific features like high µ-electron density of sp2 graphene-based nanostructure system facilitate the ability of these systems to act as suitable carriers for biomolecules. Other properties like structural size, surface carboxyl, concentration, to help the influence degree of solubility, luminescent properties. All these efforts ensure the elevated the efficacies related to emittance, loading and carrying capacity, quenching, and sensitivity of these carbon nanomaterials [107].
Do Nanomaterials Impact Exosomes (Extracellular Vesicles)?
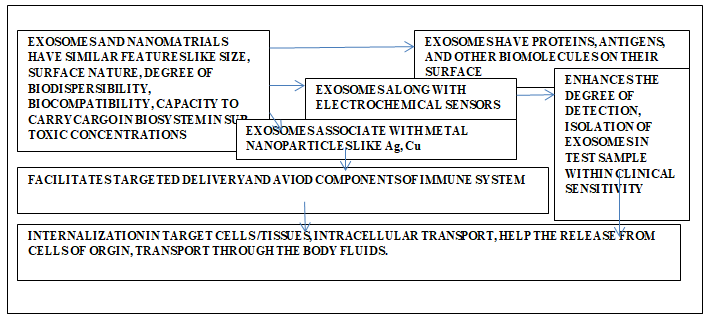
Nanoparticles have potential to elevate the degree of sensitivity of devices suitable in the field of sensing. The detection using lab-on-a-chip technology enables analysis involving very small amount of sample. Thus, this technique renders the clinical tests to be cost effective. In addition to this chip technique, there are many detection modes like colorimetry, fluorescence, surface-enhanced Raman spectroscopy, electrochemistry, nuclear magnetic resonance, etc., [99, 108, 109, 110]. Nanomaterials and exosomes have some similar features like size, surface properties, biodispersibility, biocompatibility, ability to carry cargo within biosystems etc. There is a possibility that nanomaterials in their sub-toxic concentration may influence the formation, intracellular transport, release from cell of origin, transport with the biofluids like blood, lymph, or inter cellular fluid. In such functionalities nanomaterials may participate in their pristine or functionalized forms. There seems to be very limited references on such aspects.
The nanoparticles functionalized with PLGA show features which make it suitable candidate for the biomedical application, like its ability of higher degree of biodegradability and biocompatibility, its formulation and production is suitable to carry micro and macro, hydrophilic and hydrophobic molecular drugs, its ability to protect the drug in biosystem from degradation and provide sustained release of the cargo. Such PLGA formulated nanomaterials can modify the surface features to ensure stealthness and improved interaction with the biomolecules and are better agents to target the specific tissue or cell [111]. The paracrine impacts of mesenchymal stem cells are useful for therapeutical applications because these cells secrete many useful neuroprotective and anti-inflammatory biomolecules.
The administration of pretreated mesenchymal stem cells with super paramagnetic iron oxide nanoparticle fluid MAG-D, in the experimental model, the levels of glial-derived neurotrophic factors, ciliary neurotrophic factors, hepatocyte growth factor and interlukin-10 in the dystrophic retina of rat. The degree of secretion of these four biomolecules gets affects under the different concentrations of fluid MAG-D (0, 7, 120, 274 pg iron oxide per cell respectively) in the culture medium in vitro experiments.
Retinoschisin is an extracellular protein and it plays crucial effective impact in the organization of retinal cells. This protein associates with plasma membrane of the variety of retinal cells which helps the self-sustenance of structural aspects of retina. The amount of iron oxide nanoparticle fluid MAG-D influences the innate potential of mesenchymal stem cells to produce therapeutical molecules and even from the genetically modified cells. These molecules are enclosed in exosomes produced in mesenchymal stem cell [112, 113].
The change in the surface charge of nanoparticle influences their cellular uptake, nanoparticle with positive charge exhibit higher degree of uptake in comparison to those having negative surface charge [114]. The process of internalization of superparamagnetic iron oxide nanoparticles show three modes of metabolic pathways: the macrophages RAW264.7 cells during their mitosis allow the transfer of these nanoparticles in the daughter cells, lysosomes of these cells degrade these nanoparticles thereafter, release in the intracellular cytoplasmic metabolic pool, and the unaffected superparamagnetic iron nanoparticles undergo exocytosis and expelled out of cell. Whatever the case may be the internalized nanoparticle exhibit impact on iron metabolism but no cytodamage. The metabolic impacts include up-regulation of ferritin light chain at m-RNA and level of protein, even the level of ferroprotin-1 along with mRNA elevates [115]. The composition, yield, generation of exosomes increases with nanoparticles and these stimulate the multivesicular bodies to release the exosomes. The mesenchymal stem cells show ideal uptake of magnetic nanoparticles solution as polymeric clustered superparamagnetic nanoparticles having 5 µg/ ml concentrations. The higher concentration of these nanoparticles results in decline in their viability but degree of internalization increases and form clusters. This feature depends on surface charge [113]. The study related to the interaction between exosomes and nanomaterials can pave path for understanding the mechanism involved.
Nanoparticles Help Detection and Characterization of Microsomes and Exosomes
Metal nanoparticles help the detection and characterization of microsomes and exosomes. The metal nanoparticles are in the form of chip-technology and after their electro-oxidation; in this form these identify the surface proteins on the vesicles and thus, help in their detection and characterization. For example, the chips have metal nanoparticles and undergone electro-oxidation readily and then identify the surface proteins like EpCAM and PSMA that are present on the surface of microsomes and exosomes originating in the prostate cancer cells [104]. Exosomes are potentiated agents for therapeutic uses. These agents after functionalized with supramagnetic iron oxide elevate the degree of regeneration of nervous tissue. Further, the manipulated extracellular vesicles modified with magnetic supramagnetic iron oxide nanoparticles stimulate growth factors after convenient uptake in neural tissue. These manipulated nanoparticles along with exosomes activate inter and intracellular communication, regulating the gene expression, that all enhance the out growth in the neural, thereafter undergo easy degradation [96]. The iron oxide nanoparticles induce the mesenchymal stem cells exhibit low rate of productivity of exosomes under the guidance of magnetic field. These nanoparticles induced mesenchymal stem cells induce enhance the repairing, healing process and decline in degree of apoptosis and fibrosis. These iron oxide nanoparticles induced mesenchymal stem cells elevate the angiogenesis and recovery of cardiac functionality [116]. Nanomaterials based optical biosensors facilitate detection of exosomal biomarkers (Figure 4) [117].
Nanoparticles Participate In Cellular Release of Exosomes
The exosomes are endogenous nanovesicles and play vital roles during intercellular transmission of information. The nanoparticles like magnetic iron oxide enhance the level of exosomes in the bronchoalveolar lavage fluid (BAL- Fluid) of the lungs of mice. These nanoparticles circulate comprehensively and show immunostimulatory impact on dendritic cells and sensitized T-cells in vitro studies while in vivo these magnetic iron oxide nanoparticles increased lung inflammation and induction of antibodies presenting cells to prompt Th-1 responses in the case mice [118, 119, 120]. The studies on two types of cells, namely, human peripheral blood mononuclear cells (PBMC) and monocytes derived dendric cells (MDDC) reveal no major differences in the phenotypes or liberation of exosomes after sub-toxic exposure of TiO2 and zinc oxide (ZnO) nanoparticles. It was very difficult to detect any interference during the trafficking or transporting, and release of exosomes because of their nanosized. Techniques, like the flow cytometry and total protein contents analysis of the sample of the generated exosomes, could not help the evaluation of generation of exosomes bound onto micro- sized beads and very less quantity of yield of exosomes [120].
The iron oxide nanoparticles functionalized with poly (lactic-co-glycolic acid) (PLGA) are relatively more biodispersible; hence, exhibit better bioavailability in biosystems. The physicochemical features like surface charge, of nanoparticles stimulate the production of exosomes from stem cells. Pluripotent stromal cells like mesenchymal stem cells produce extracellular vesicles and are functionally important because these vesicles carry metabolites like growth factors and antioxidants. These cargos are helpful to the ambient cells. These vesicles have nano size, and are biocompatible and biostable structures. Under natural conditions, their yield is relatively low, the same enhances with the help of specifically modified nanoparticles. The surface modifications of nanoparticles stimulate the cellular release of exosomes [113]. The amphiphilic cationic (cCHP) nanogel formulated based on the surface polymer engineering technique [121]. This technique helps to modify the physical, chemical, electrical, magnetic, optical, mechanical, wear-resistant, corrosion-resistant features of the surface. Specifically, the surface morphology, mechanical capability, and chemistry, play significant role during adhesion, the process of film formation and finalizing functionality of the film [122]. The amphiphilic cationic (cCHP) nanogel forms a complex with exosome as a result the delivery of exosome becomes better. The mouse macrophage cell mixed with cCHP nanogel show that the HeLa cells internalize the resultant hybrid exosome effectively due to endocytosis even in the presence of endocytic inhibitors. The nanogel system facilitates the release of functional exosome into cells. During this successful release of exosomes neuron- like differentiation has been seen in the recipient cells [121].
Molecular Crowding, Exosomes, and Nanomaterials
The cytoplasm of a living cell is a suitable example of a highly crowded environment that maintains homeostasis. It is a site of interactions regulated by the principles of kinetics and thermodynamics. The molecular crowding affects the rates and equilibria constants of the biochemical and biomolecular reaction occurring therein. These effects are due to the mobility of reactants, reactions that cause the attraction between them, resultant depletion interactions, and non-specific biochemical interactions amongst crowding molecules excluding the effect of volume-as this factor is mostly constant [123, 124, 125, 126, 127, 128, 129, 130]. Molecular crowding plays a very significant functional role in cytophysiology. Usually, nucleic acids, proteins, polysaccharides, soluble and non- soluble materials are present in the cytoplasm. These components form around 20 to 40 percent of the cellular volume. During molecular crowding, the concentration of its constituents is about 400 g/L(-1). Molecular crowding occurs during molecular, biochemical, medical, pharmaceutical, biotechnological, and nanotechnological interactions and the related applications to the biological system [131].
The cellular functions ranging from the emergence of life up to cell size regulation is related to the crowding of molecules. The cytoplasmic viscosity depends on the length scale concept, interaction related to depletion, non- specific binding, and the non-specific biomolecules present in the cytoplasm [132, 133, 134]. Molecular crowding influences the mobility, the interactions of biomolecules. As a result, parameters like diffusion constants, reaction kinetics get influenced. This aspect affects inter and intramolecular interactions, the tendency of molecules to retain their original stereoscopic, functional, and resultant interactive states. There are physicochemical variations at a molecular level in the cytoplasm of prokaryotes and eukaryotes. These may govern inter and intracytoplasmic and nuclear diffusion constants in these biosystems. In such cases, the kinetics of interactions includes protein-protein, protein-DNA or RNA, and gene expression reactions.
The viscosity of the synthetic or biological systems depends on the phenomenological pattern of diffusion. The diffusion coefficient of the probe particle depends on the structure of the fluid; it may be like polymeric, micellar, or colloidal structure, understudy. The protein and colloidal solutions exhibit long time self-diffusion and macroscopic viscosity that describes the properties of transportation. The phenomenological concept controls this process. This concept is based on the implicational aspect of theoretical physics to experimental data and predicts the consequences. This concept does not stress the experimentation based on scientific methodology to evaluate the scientific hypothesis. Thus, phenomenology is a philosophical notion and anticipates real behavior. It is one of the most suitable techniques to investigate the behavior of particles present in body fluids based on the principle of particle physics [135]. This concept depends on the defined measurable parameters such as the hydrodynamic radius of the particles, and the volume of the fraction. This technique is suitable for investigating viscosity and diffusion occurring in the cytoplasm. The concentration and size of crowding particles influence this technique [136, 137, 138]. The polymer systems are the suitable mimicking model that helps the study of the rate of diffusion, and responses related to viscosity in the case of the biological system even though, at times, the cytoplasm is regarded as poroelastic matter [139].
In a fluid medium, the mobility of its constituents corresponds to the size, viscosity of the fluid involving the Einstein equation, and the collision concept. The kinetics of the intermolecular interaction, and their intrinsic action, affects the Brownian movements and the rate of particle collision. Extracellular vesicles, their biodistribution, and their intracellular interactions are ambiguous in a biosystem because of the differences in surface charges on the particles. Nanoparticles and nonfunctionalized exosomes have a net negative charge depending on the nature of their surface molecules. Body fluids are complex, dynamic, and homeostatic phenomena, specifically in the case of vertebrates. These fluids behave like a solution, colloidal, and suspension as these permit dissolution of minerals/salts, distribution of colloidal particles, and suspension media for the insoluble matter. The parameters like zeta potential, size, and nature of the materials to be distributed within the biosystem, control the behavior of the components of body fluids. Zeta potential provides the degree of stability of colloidal particles; the dispersibility, stability of components of the suspension, colloidal mixture in disperse system, and relate to the electrophoretic mobility, electrical charge on particles, particle-particles interaction, particle-medium interaction, degree of aggregation among the constitutional particles. These parameters play a significant behavioral role in extracellular vesicles or exosomes in the body fluids as these fluids behave like a solution, colloidal, and suspension media. This behavior becomes evident during cellular uptake and cytotoxic investigations.
The equation of van Smoluchowski describes the distribution of particle size depending on time and per interaction among the particles. The electrophoretic mobility of charged particles depends on the physicochemical features of the dispersing particles. Overall, the surface of the biological plasma membrane has a negative charge due to the glycosylated proteins which intercalate the lipid bilayer. On suspension, cellular plasma membrane and exosomes show negative surface charge. The surface charge of exosomes interrelates with the degree of ionization of the groups present on the surface, protonated states, bindings present or formed within a molecule and different molecules, hydrogen bonds, adsorption of electrolytes, and chemical nature of the grafted chain. Exosomes associated with nanoparticles are among the suitable agents that may maintain stability in a body fluid. These act as a carrier for drugs, biomolecules, pharmacokinetics, formulated nanomedicines [110, 140, 141, 142, 143, 144, 145, 146].
The exosomes play a major role in eliminating nanoparticles. The exosomes perform double function; first function is to trap nanoparticles in the biosystem and second function is to eliminate them from the body. The primary human macrophages scavenge the nanoparticles and expel them via exosomes, thereby, enhancing natural shuttling of nanomaterials [147]. The exosomes are the products of endosomal pathway and play significant roles in the paracrine and autocrine cellular communications. The malicious cells, cancer cells, and other dysfunctioning cells produce exosome; these exosomes participate in progression of malicious micro and ambient cellular environment in the affected tissues. The nanomaterials have specific physicochemical properties like, high surface volume ratio, and ease to get functionalized as per the need. These properties become more effective when they associate with exosomes and act as multiparametric molecules. These molecules open wide horizon for pondering and delving in biological, clinical, and diagnostic fields. There exists appreciable compatibility between exosomes and nanomaterials. This aspect plays major role during multiparametric molecular profiling and also of as vesicles in biosystem. This helps in evaluation of biophysical and biomolecular configuration of vesicles that too in native biofluids.
The nanotags consisting of gold and other nanomaterials increase the degree of sensing, imaging, and quantitative evaluation of exosomes markers [148, 149, 150]. Light causes oscillations in the electrons present on the surface of nanopalsmonic nanomaterials. Such nanomaterials in the form of optical antenna play role at interface, and find suitable applications in translational medical field related to biosystem. This aspect is more suitable to explore intracellular molecular activities, delivery of gene, and precisely diagnose molecular interactions in vivo and in vitro. It is feasible to investigate biological processes like electron tunneling, electron transfer, and quantum mechanism related biological processes. The encapsulated biocompatible nanoparticles act as efficient agents for theranostics application specifically for cancer, and metabolic disorders. This is possible only because the exosomes are very effective, and target oriented successful vehicles in biosystem. Such associated nanomaterials open wide range of possibilities of applications in the therapies like photothermic, magnetic hyperthermia, and gene silencing and drug delivery systems. These exosome associated nanomaterials show favorable degree of penetration, appropriate biodistribution, biostability, lower level of immunogenicity, and ability to move across most of the barriers in biosystems. The templated plasmonics technique is applicable to exosomes, and involves the formation of gold nanoshells. These nanoshells get assembled and piled up on vesicles within biosystem. This assembled templated formulation acts as exosomal markers, and helps their scanning and examination. These targets bound nanoshells exhibit biomolecular selectivity which is useful in expressing specific plasmonic signature quencher for fluorescence probes. The optical and absorbance signals produced facilitate the analytical studies of vesicles. This feature is useful in determining the exosomal biomarkers in the medical field.
Conclusion
Exosomes being cellular product exhibit higher degree of biostability, biodistribution, and have morphological dimensions similar to nanomaterials. These cellular products pose difficulties during the estimation of their yield and quantification. Exosome and nanomaterials exhibit similar features like size, surface nature, degree of biodispersibility, biocompatibility, cargo capacity, release of cargo at predetermined target, in sub-toxic concentrations. Their impact on cellular molecular crowding seems to be within physiological limits. When exosomes get associated with nanomaterials, there is an additional enhancement in the multi-utility features and other functional efficacies of both. Such efficiencies include their cargo capacity, biodistribution, release of the cargo at the predetermined targets. Some of the nanomaterials, like TiO2 and ZnO, can influence their cellular production under sub-toxic concentrations. It is relatively convenient to trace them in biosystem because these nanomaterials show appropriate cellular and tissue real-time imaging ability. The extracellular vesicles and microvesicles are effective agents for intra and intercellular communications. This feature provides a scope of their applications to understand the mechanism involved during such transmissions. These vesicles have feasible molecular self-assembly nature, ability to provide suitable protections and intracellular delivery of biofunctional molecules. The exosomes and nanomaterials, together offer a wide horizon for investigation, experimentation, innovation, and applications in the fields of biochemistry, medicine, pharmaceuticals, molecular, and cell biology.
Conflicts of Interest
The authors confirm that there are no conflicts of interest.
Funding
No funding is involved for this presentation from any source.
References
-
Hessvik NP, Llorente A (2018) Current knowledge on exosome biogenesis and release. C Cell Mol Life Sci 75(2): 192-208.
-
Dear JW, Street JM, Bailey MA (2013) Urinary exosomes: a reservoir for biomarker discovery and potential mediators of intra renal signaling. Proteomics 13(10- 11): 1572-1580.
-
Song J, Chen X, Wang M, Xing Y, Zheng Z, et al. (2014) Cardiac endothelial cell-derived exosomes induce specific regulatory B-cells. Sci Rep 4: 7583.
-
Madison MN, Roller RJ, Okeoma CM (2014) Human semen contains exosomes with potent anti-HIV-1 activity. Retrovirology 11: 102.
-
Oves M, Qari HA, Felemban NM, Khan AAP, Rehan M, et al. (2018) Exosomes: A paradigm in drug development against cancer and infectious diseases. J Nanomaterials.
-
Edgar JR (2016) What are exosomes, exactly? BMC Biology 14: 46.
-
Keller S, Sanderson MP, Stoeck A, Alvogt P (2006) Exosomes: from biogenesis to secretion to biological functions. Immunological Letters 107(2): 102-108.
-
van der Pol E, Boing AN, Harrison P, Sturk A, Niewland R (2012) Classification, functions and clinical relevance of extracellular vesicles. Pharmacol Rev 64(3): 676-705.
-
Huleihel L, Hussey GS, Naranjo JD, Zhang L, Dziki JL, et al. (2016) Matrix bound nanovesicles within extracellular matrix bioscaffolds. Science Adv 2(6): e1600502.
-
Spaull R, McPherson B, Gialeli A, Clayton A, Uney J, et al. (2019) Exosome populate cerebrospinal fluid of preterm infants with post-hemorrhage hydrocephalus. Int J Dev Neurosci 73(1): 59-65.
-
Huotari J, Helenius A (2011) Endosome maturation. EMBO J 30(17): 3481-3500.
-
Von Bartheld CS, Altick AL (2011) Multivesicular bodies in neurons: distribution, protein contents and trafficking functions. Prog Neurobiol 93(3): 313-340.
-
Harding C, Heuser J, Stahl P (1983) Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J Cell Biol 97(2): 329-339.
-
Pan BT, Teng K, Wu C, Adam M, Johnstone RM (1985) Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J Cell Biol 101(3): 942-948.
-
Mittelbrunn M, Gutierrez Vazquez C, Villarroya Beltri C, Gonzalez S, Sanchez Cabo F, et al. (2011) Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nature Communications 2: 282.
-
Gonzalez Calero L, Martin Lorenzo M, Alvarez Llamas G (2014) Exosomes: a potential key target in cardio-renal syndrome, Frontiers in Immunology 5: 465.
-
Record M, Poirot M, Silvente Poirot S (2014) Emerging concepts on the role of exosomes in lipid metabolic diseases. Biochimie 96: 67-74.
-
Greening DW, Gopal SK, Xu R, Simpson RJ, Chen W (2015) Exosomes and their roles in immune regulation and cancer. Semin Cell Dev Biol 40: 72-81.
-
Gangoda L, Boukouris S, Liem M, Kalra H, Mathivanan S (2015) Extracellular vesicles including exosomes are mediators of signal transduction: are they protective or pathogenic?. Proteomics 15(2-3): 260-271.
-
Lasser C, O Neil SE, Shelke GV, Sihlbom C, Hansson SF, et al. (2016) Exosomes in the nose induce immune cell trafficking and harbor an altered protein cargo in chronic airway inflammation. J Transl Med 14(1): 181.
-
Minciacchi VR, Freeman MR, Di Vizio D (2015) Extracellular vesicles in cancer: exosomes, microvesicles and the emerging role of large oncosomes. Semin Cell Dev Biol 40: 41-51.
-
Howitt J, Hill AF (2016) Exosomes in the pathology of neurodegenerative diseases. J Biol Chem 291(52): 26589-26597.
-
Kishore R, Garikipati VN, Gumpert A (2016) Tiny shuttles for information transfer: exosomes in cardiac health and disease. J Cardiovasc Transl Res 9(3): 169-175.
-
Salem KZ, Moschetta M, Sacco A, Imberti L, Rossi G, et al. (2016) Exosomes in tumor angiogenesis. Methods Mol Biol 1464: 25-34.
-
Zhang Y, Liu Y, Liu H, Tang WH (2019) Exosomes: biogenesis, biologic function and clinical potential. Cell and Biosciences 9: 19.
-
Raposo G, Tenza D, Mecheri S, Peronet R, Bonnerot C, et al. (1997) Accumulation of major histocompatibility complex class II molecules in mast cell secretory granules and their release upon degranulation. Mol Biol Cell 8: 2631-2645.
-
Thery C, Regnault A, Garin J, Wolfers J, Zitvogel L, et al. (1999) Molecular characterization of dendritic cell- derived exosomes, Selective accumulation of the heat shock protein hsc73. J Cell Biol 147(3): 599-610.
-
Savina A, Fader CM, Damiani MT, Colombo MI (2005) Rab11 promotes docking and fusion of multivesicular bodies in a calcium-dependent manner. Traffic 6(2): 131-143.
-
van Niel G, Porto Carreiro I, Simoes S, Raposo G (2006) Exosomes: A common pathway for a specialized function. J Biochem 140(1): 13-22.
-
Gurunathan S, Kang MH, Jeyaraj M, Qasim M, Kim JH (2019) Review of the isolation, characterization, biological function, and multifarious therapeutic approaches of exosomes. Cells 8(4): 307.
-
Luan X, Sansanaphongpricha K, Myers I, Chen H, Yuvan F, et al. (2017) Engineering exosomes as refined biological nanoplatforms for drug delivery. Acta Pharmacologica Sinica 38: 754-763.
-
Kim MS, Haney MJ, Zhao Y, Mahajan V, Deygen I, et al. (2015) Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine 12(3): 655-664.
-
Hunter RJ (2013) Zeta potential in colloid science. Academic Press, London, New York, Toronto, Sydney.
-
Xu R, Greening DW, Zhu HJ, Takahashi N, Simpson RJ (2016) Extracellular vesicles isolation and characterization towards clinical application. J Clin Invest 126(4): 1152-1162.
-
Beit Yannai E, Tabak S, Stamer WD (2018) Physical exosomes: exosomes interactions. J Cell Mol Med 22(3): 2001-2006.
-
Kobayashi T, Marie Helen B, Chaverllier J, Makino A, Mayran Nb, et al. (2002) Separation and characterization of late endosomal membrane domains. J Biol Chem 277(35): 32157-32167.
-
Gaullier JM, Simonsen A, D Arrigo A, Bremnes B, Stenmark H, et al. (1998) FYVE fingers bind PtdIns (3)P. Nature 394(6692): 432-433.
-
Wishart MJ, Taylor GS, Dixon JE (2001) Phoxy lipids: revealing PX domains as phosphoinositide Binding modules. Cell 105 (7): 817-820.
-
Gillooly DJ, Morrow IC, Lindsay M, Gould R, Bryant NJ, et al. (2000) Localization of phosphatidylinositol 3-phosphate in yeast and mammalian cells. EMBO J 19(17): 4577-4588.
-
Gillooly DJ, Simonsen A, Stenmark H (2001) Cellular functions of phosphatidylinositol 3-phosphate and FYVE domain proteins. Biochem J 355(Pt2): 249-258.
-
Simonsen A, Stenmark H (2001) PX domains: attracted by phosphoinositides. Nature Cell Biology 3: E179-E181.
-
Johannes L, Wunder C, Bassereau P (2014) Bending on the rocks, A cocktail of biophysical modules to build endocytic pathways. Cold Spring Harb Perspect Biol 6(1): a016741.
-
Rawicz W, Olirich KC, McIntosh T, Needham D, Evans E (2000) Effects of chain length and unsaturation on elasticity of lipid bilayers. Biophysics Journal 79(1): 328- 339.
-
Roux A, Cavalier P, Prost J, Bassereau P, Goud B (2005) Role of curvature and phase transition in lipid sorting and fission of membrane tubules. EMBO J 24(8): 1537- 1545.
-
Helfrich W (1973) Elastic properties of a lipid bilayer: theory and problem experiments. Z Naturforsch C 28(11): 693-703.
-
Lipowsky R (2014) Coupling of bending and stretching deformations in vesicle membrane. Advances in Colloid and Interface Science 208: 14-24.
-
Simunovic M, Evergren E, Golushko I, Prevost C, Renard HF, et al. (2016) How curvature generating proteins build a scaffold on Membrane nanotubes. Proceedings of the National Academy of Science of USA 113(40): 11226- 11231.
-
Compelo F, McMahon HT, Kozlov MM (2008) The hydrophobic insertion mechanism of membrane curvature generated by proteins. Biophys J 95(5): 2325- 2339.
-
Sens P, Johannes L, Bassereau P (2008) Biophysical approaches to protein induce 3D membrane deformation in trafficking. Curr Opin Cell Biol 20(4): 476-482.
-
Rossman JS, Lamb AR (2011) Influenza virus assembly and budding. Virology 411(2): 229-236.
-
Gautreau A, Ognievetskaia K, Ungermann C (2014) Function and regulation of endosomal fusion and mechanics. Cold Spring Harb Perspect Biol 6(3): a16832.
-
Hu J, Shibata Y, Voss C, Shemesh T, Li Z, et al. (2008) Membrane protein of endoplasmic reticulum induces high curvature tubes. Science 319(5867): 1247-1250.
-
Anthonny B (2011) Mechanism of membrane curvature sensing. Annu Rev Biochem 80: 101-123.
-
Aimon S, Callen Jones A, Berthand A, Pinot M, Toombes GES, et al. (2014) Membrane shape modulates transmembrane protein distribution. Dev Cell 28(2): 212-218.
-
Watkins EB, Miller CE, Majewski J, Kuhl TL (2011) Membrane texture induced by specific protein binding and receptor clustering: Active roles for lipids in cellular function. Proceedings of National Academy of Science of the USA 108(17): 6975-6980.
-
Gowrishankar K, Ghosh S, Saha SCR, Mayor S, Rao M (2012) Active remodeling of cortical actin regulates the spatiotemporal organization of cell surface molecules. Cell 149(6): 1353-1367.
-
Mim C, Unger VM (2012) Membrane curvature and its generation by BAR proteins. Trends Biochem Sci 37(12): 526-533.
-
Frost A, Unger VM, De Camilli P (2009) The BAR domain superfamily: Membrane-molding macromolecules. Cell 137(2): 191-196.
-
Lim J, Hwang SY, Moon S, Kim S, Kim WY (2020) Scaffold- based molecular design with graph generative model. Chemical Science 11(4): 1154-1164.
-
Lemmon S, Kukulski W, Hetty EM, Specht T, Briggs JAG, et al. (2018) The Contributions the actin machinery to endocytic membrane bending and vesicle formation. Mol Biol Cell 29(11): 1346-1358.
-
Stachowiak JC, Hayden CC, Sasaki DY (2010) Steric confinement of proteins on lipid membranes can drive curvature and tabulation. Proceedings of National Academy of Science of the USA 107(17): 7781-7786.
-
Stachowiak JC, Schmid EM, Ryan CJ, Ann HS, Sasaki DY, et al. (2012) Membrane bending by protein-protein crowding. Nat Cell Biol 14(9): 944-949.
-
McMahon HT, Mills IG (2004) COP and clathrin- coated vesicle budding: Different pathways, common approaches. Curr Opin Cell Biol 16(4): 379-391.
-
Stagg SM, La Pointe P, Balch WE (2007) Structural design of cage and coat scaffolds that direct membrane traffic. Curr Opin Struct Biol 17(2): 221-228.
-
Masuda M, Mochizuki N (2010) Structural characteristics of BAR domain superfamily to sculpt the Membrane. Semin Cell Dev Biol 21(4): 391-398.
-
Schmid SL, Frolov VA (2011) Dynamin: Functional design of a membrane fission catalyst. Annu Rev Cell Dev Biol 27: 79-105.
-
Ferguson SM, De Camilli P (2012) Dynamin, a membrane- remodeling GTPase. Nature Review Molecular and Cell Biology 13(2): 75-88.
-
Schekman R, Orci L (1996) Coat proteins and vesicle budding. Science 271(5255): 1526-1533.
-
Lahir YK (2016) Some aspects of interactions between nanomaterials and the cytoskeleton of eukaryotic cells. Adv Clin Toxicolo 1(2): 1-10.
-
Sheetz M (2001) Cell control by membrane-cytoskeleton adhesion. Nature Reviews Molecular Cell Biology 2: 392- 396.
-
Kranenburg M, Smit B (2005) Phase Behavior of Model Lipid Bilayers. J Phys Chem B 109(14): 6553-6563.
-
Salbreux G, Charras G, Paluch E (2012) Actin cortex mechanics and cellular morphogenesis. Trends Cell Biology 22(10): 536-545.
-
Diz-Muñoz A, Fletcher DA, Weiner OD (2013) Use the force: membrane tension as an organizer of cell shape and motility. Trends Cell Biol 23(2): 47-53.
-
Lieber AD, Yehudai Resheff S, Barnhart EL, Theriot JA, Keren K (2013) Membrane Tension in rapidly moving cells is determined by cytoskeletal forces. Curr Biol 23(15): 1409-1417.
-
Sonnino S, Prinettim A (2013) Membrane Domains and the “Lipid Raft” Concept. Curr Med Chem 20(1): 4-21.
-
Sezgin E, Lavental I, Mayor S, Eggeling C (2017) The mystery of membrane organization: Composition, regulation and role of lipid rafts. Nat Rev Mol Cell Biol 18(6): 361-374.
-
Johannes L, Mayer S, (2010) Induced domain formation in endocytic invagination, lipid sorting, and scission. Cell 142(4): 507-510.
-
Romer W, Berland L, Chambon V, Gaus K, Windchiegl B, et al. (2007) Shiga toxins induce tubular membrane invagination for its uptake into cells. Nature 450(7170): 670-675.
-
Evans E, Smith BA (2011) Kinetics of Hole Nucleation in Biomembrane Rupture. New J Phys 13: 095010.
-
Chappei JS, Mears JA, Fang S, Leonard M, Schmid SL, et al. (2011) A pseudoatomic model of the dynamin polymer identifies a hydrolysis dependent power stoke. Cell 147(1): 209-222.
-
Campelo F, Malhotra V (2012) Membrane fission: The biogenesis of transport carriers. Annu Rev Biochem 81: 407-427.
-
Morlot S, Galli V, Klein M, Chiaruttini N, Manzi J, et al. (2012) Membrane shape at the edge of the dynamin helix sets location and duration of the fission reaction. Cell 151(3): 619-629.
-
Shnyrova AV, Bashkirov PV, Akimov SA, Pucadyil TJ, Zimmerberg J, et al. (2013) Geometric catalysis of membrane fission driven by flexible dynamin rings. Science 339(6126): 1433-1436.
-
Gruenberg J, Stenmark H (2004) The biogenesis of multivesicular endosomes. Nat Rev Mol Cell Biol 5(4): 317-323.
-
Piper RC, Katzmann DJ (2007) Biogenesis and Function of Multivesicular Bodies. Annu Rev Cell Dev Biol 23(1): 519-547.
-
Isola A, Chen S (2017) Exosomes: The Messengers of Health and Disease. Curr Neuropharmacol 15(1):157- 165.
-
Brennan K, Martin K, Fitz Gerald SP, Sullivan JO, Wu Y, et al. (2020) A comparison of methods for the isolation and separation of extracellular vesicles from protein and lipid particles in human serum. Scientific Reports 10(1039).
-
Henne WM, Buchkovich NJ, Emr SD (2011) The ESCRT pathway. Dev Cell 19 21(1): 77-91.
-
Trajkovic K, Hsu C, Chiantia S, Rajendran L, Wenzel D, et al. (2008) Ceramide triggers the budding of exosome vesicles into multivesicular endosomes. Science 319(5867): 1244-1247.
-
Stuffers S, Wegner CS, Stenmark H, Andreas Brech A (2009) Multivesicular Endosome Biogenesis in the Absence of ESCRTs. Traffic 10(7): 925-937.
-
Edgar JR, Eden ER, Futter CE (2014) Hrs-and CD63- dependent competing mechanisms make different sized endosomal intraluminal vesicles. Traffic 15(2): 197-211.
-
Parolini I, Federici C, Raggi C, Lugini L, Palleschi S, et al. (2009) Microenvironmental pH is a key factor for exosome traffic in tumor cells. J Biol Chem 284(49): 34211-34222.
-
Meldolesi J (2018) Exosomes and ectosomes in intercellular communication. Curr Biol 28(8): 435-444.
-
Bebelman MP, Bun P, Huveneers S, van Niel G, Pegtel DM, et al. (2020) Real-time imaging of multivesicular body plasma membrane fusion to quantify exosome release from single cells, Nat Protoc 15(1): 102-121.
-
Rajendran L, Jitin Bali J, Barr MM, Court FA, Kramer Albers EM, et al. (2014) Emerging roles of extracellular vesicles in nervous system. J Neurosci 34(46): 15482- 15489.
-
Neubert J, Glumm J (2016) Promoting neural regeneration using extracellular vesicles loaded with supramagnetic iron oxide nanoparticles. Neural Regen Res 11(1): 61-63.
-
Logozzi M, De Milito A, Lugini L, Borghi M, Calabrò L, et al. (2009) High Levels of Exosomes Expressing CD63 and Caveolin-1 in Plasma of Melanoma Patients. PLoS One 4(4): e5219.
-
Duijvesz D, Versluis CYL, Van Der Fels CAM, Vredenbregt Van Den BMS, Leivo MS, et al. (2015) Immuno-based detection of extracellular vesicles in urine as diagnostic marker for prostate cancer. Int J Cancer 137(12): 2869- 2878.
-
Vasconcelos MH, Caires HR, Ābols A, Xavier CPR, Linē A (2019) Extracellular vesicles as a novel source of biomarkers in liquid biopsies for monitoring cancer progression and drug resistance. Drug Resist Updat 47: 100647.
-
Martın Gracia B, Martin Barreiro A, Cuestas Ayllon C, Grazu V, Line A, et al. (2020) Nanoparticle-based biosensors for detection of extracellular vesicles in liquid biopsies. J Materials Chemistry B 8(31): 6710-6738.
-
Sadovska L, Eglitis IJ, Line A (2015) Extracellular vesicles as biomarkers and therapeutic targets in breast cancer. Anticancer Res 35(12): 6379-6390.
-
Castellanos Rizaldos E, Grimm DG, Tadigotla V, Hurley J, Healy J, et al. (2018) Exosome-Based Detection of EGFR T790M in plasma from non-small cell lung cancer patients. Clin Cancer Res 24(12): 2944-2950.
-
Vaidyanathan R, Naghibosadat M, Rauf S, Korbie D, Carrascosa LG, et al. (2014) Detecting exosomes specifically: a multiplexed device based on alternating current electrohydrodynamic induced nanoshearing. Anal Chem 86(22): 11125-11132.
-
Zhaou YG, Mohamadi RM, Poudineli M, Kermanshah L, Ahmed SU, et al. (2015) Interrogating circulatory microsomes and exosomes metal nanoparticles. Nano- Micro Small 12(6): 727-732.
-
Fathi P, Rao L, Chen X (2020) Extracellular vesicle- coated nanoparticles. View 2(2): 20200187.
-
Cheng G, Li W, Ha L, Han X, Hao S, et al. (2018) Self-assembly of extracellular vesicle-like metal– organic framework nanoparticles for protection and intracellular delivery of biofunctional proteins. J Am Chem Soc 140(23): 7282-7291.
-
Goryacheva OV, Vostrikova AM, Kokorina AA, Mordovina EA, Tsyupka DV, et al. (2019) Luminescent carbon nanostructure for microRNA detection TrAC. Trends in analytical Chemistry 119: 115613.
-
Issadore D, Shao H, Chung J, Newton A, Pittet M, et al. (2011) Self-assembled magnetic filter for highly efficient immunomagnetic separation. Lab on a Chip 11(1): 147- 151.
-
Jeong J, Park D, Pathania CM, Castro Weissleder R, Lee H (2016) Integrated Magneto-Electrochemical Sensor for Exosome Analysis. ACS Nano 10(2):1802- 1809.
-
Lahir YK, Avti P (2020) Nanomaterials and their interactive behavior with biomolecules, Cells and Tissues. Bentham Science Publishers, Singapore.
-
Danheir F, Ansovena E, Silva JM, Coco R, Breton AL, et al. (2012) PLGA based nanoparticles: An over view of biomedical applications. J Control Release 161(2): 505- 522.
-
Bashar AE, Metcalfe A, Yanai A, Lavor C, Hafeli UO, et al. (2013) Influence of iron oxide nanoparticles on innate and genetically modified secretion of mesenchymal stem cells, IEEE Trans Magn 49(1): 380-393.
-
Park DJ, Yun WS, Kim WC, Park JE, Lee SH, et al. (2020) Improvement of stem cells derived exosome release efficiency by surface-modified nanoparticles. J Nanobiotechnology 18: 178.
-
Jiang XE, Musyanovych A, Rocker C, Landfester K, Mailander V, et al. (2011) Specific effects of surface carboxyl groups on anionic polystyrene particles in their interactions with mesenchymal stem cells. Nanoscale 3(5): 2028-2035.
-
Gu J, Xu H, Han Y, Dai W, Hao W, et al. (2011) The internalization path way, metabolic fate and biological effect of superparamagnetic iron oxide nanoparticles in the macrophage like RAW264.7 cell. Sci China Life Sci 54(9): 793-805.
-
Lee JR, Park BW, Kim J, Choo YW, Kim HY, et al. (2020) Nanovesicles derived from iron oxide nanoparticles– incorporated mesenchymal stem cells for cardiac repair. Sci Adv 6(18): eaaz0952.
-
Shao B, Xiao X (2020) Recent achievements in exosomal detection by nanomaterial based optical Biosensors- A review. Analytica Chimica Acta 1114: 74- 84.
-
Zhu M, Li Y, Li J, Feng W, Nie G, Zhao Y (2012) Exosomes as extra-pulmonary Signalling conveyors for nanoparticle-induced systemic immune activation. Small 8(3): 404-412.
-
Zhu M, Tian X, Song X, Li Y, Tian Y, et al. (2012) Nanoparticle-induced exosomes target antigen- presenting cells to initiate Th1-type immune activation. Small 8(18): 2841-2848.
-
Anderson Willman B, Gehermann U, Cansu Z, Buerki Thurnberr T, Krug HP, et al. (2012) Effects of sub-toxic concentrations of TiO2 and ZnO nanoparticles on human lymphocytes, dendritic cells and exosome production. Toxicol Appl Pharmacol 264(1): 94-103.
-
Sawada SI, Sato YF, Kawasaki R, Yasuoka JI, Mizuta R, et al. (2019) Nanogel hybrid assembly for exosome intracellular delivery: effects on endocytosis and fusion by exosome surface polymer engineering. Biomaterials Science 8(2): 619-630.
-
Petkoska AT, Nasov I (2014) Surface engineering of polymers-case study: PVD coating on polymers. Zastta Materijala 55: 1-8.
-
Berg OG (1990) The influence of macromolecular crowding on thermodynamic activity: solubility and dimerization constants for spherical and dumbbell- shaped molecules in a hard sphere mixture. Biopolymers 30(11-12): 1027-1037.
-
Minton AP (2005) Influence of macromolecular crowding upon the stability and state of association of proteins: predictions and observations. Journal Pharm Sci 94(8): 1668-1675.
-
Marenduzzo D, Finan K, Cook PR (2006) The depletion attraction: an underappreciated force driving cellular organization. J Cell Biol 175(5): 681-686.
-
Zhou HX, Rivas G, Minton AP (2008) Macromolecular crowding and confinement: biochemical, biophysical, and potential physiological consequences. Annu Rev Biophys 37: 375-397.
-
Minton AP (2013) Quantitative assessment of the relative contributions of steric repulsion and chemical interactions to macromolecular crowding. Biopolymers 99(4): 239-244.
-
Sarkar M, Li C, Pielak GJ (2013) Soft interactions and crowding. Biophys Rev 5(2): 187-194.
-
Sarkar M, Smith AE, Pielak GJ (2013) Impact of reconstituted cytosol on protein stability, Proceedings of National Academic Science, USA 110(48): 19342-19347.
-
Zhou EH, Martinez FD, Fredberg JJ (2013) Cell rheology: mush rather than machine. Nat Mater 12(3): 184-185.
-
Miyoshi D, Sugimoto N (2008) Molecular crowding effects on structure and stability of DNA. Biochimie 90(7): 1040-1051.
-
Kalodimos CG, Biris N, Bonvin AM, Levandoski MM, Guennuegues M, et al. (2004) Structure and flexibility adaptation in nonspecific and specific protein-DNA complexes. Science 305(5682): 386-389.
-
Spitzer J, Poolman B (2009) The role of biomacromolecular crowding, ionic strength, and physicochemical gradients in the complexities of life’s emergence. Microbiol Mol Biol Rev 73(2): 371-388.
-
Soh S, Banaszak M, Kandere Grzybowska K, Grzybowski BA (2013) Why cells are microscopic: a transport-time perspective. J Phys Chem Lett 4(6): 861- 865.
-
Marder MP (2010) Condensed matter physics, 2nd (Edn.), John Wiley and sons.
-
Holyst R, Bielejewska A, Szymański J, Wilk A, Patkowski A, et al. (2009) Scaling form of viscosity at all length-scales in poly (ethylene glycol) solutions studied by fluorescence correlation spectroscopy and capillary electrophoresis. Physical Chemistry and Chemical Physics 11(40): 9025-9032.
-
Kalwarczyk T, Ziebacz N, Bielejewska A, Zaboklicka E, Koynov K, et al. (2011) Comparative analysis of viscosity of complex liquids and cytoplasm of mammalian cells at the nanoscale. Nano Letters 11(5): 2157-2163.
-
Kalwarczyk T, Sozanski K, Jakiela S, Wisniewska A, Kalwarczyk E, et al. (2014) Length-scale dependent transport properties of colloidal and protein solutions for prediction of crystal nucleation rates. Nanoscale 6(17): 10340-10346.
-
Zhou HX (2013) Influence of crowded cellular environments on protein folding, binding, and oligomerization: biological consequences and potentials of atomistic modeling. FEBS Lett 587(8): 1053-1061.
-
Delgado AV, González Caballero F, Hunter RJ, Koopal LK (2007) Measurement and interpretation of electrokinetic phenomena. Journal of Colloid Interface Science 309(2): 194-224.
-
Akagi T, Ichiki T (2008) Cell electrophoresis on a chip: what can we know from the changes in electrophoretic mobility? Anal Bioanal Chem 391(7): 2433-2441.
-
Kaszuba M, Corbett J, Watson FM, Jones A (2010) High-concentration zeta potential measurements using light-scattering techniques. Philos Trans A Math Phys Eng Sci 368(1927): 4439-4451.
-
Frohlich E (2012) The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int J Nanomedicine 7: 5577-5591.
-
Bhattacharjee S, Rietjens IMC, Singh MP, Atkins TM, Purkait TK, et al. (2013) Cytotoxicity of surface- functionalized silicon and germanium nanoparticles: the dominant role of surface charges. Nanoscale 5(11): 4870-4883.
-
Bhattacharjee S (2016) DLS and zeta potential-what they are and what they are not?. Journal of Controlled Release 235: 337-351.
-
Midekessa G, Godakumara K, Ord J, Viil J, Lättekivi F, et al. (2020) Zeta Potential of Extracellular Vesicles: Toward Understanding the Attributes that Determine Colloidal Stability. ACS Omega 5(27): 16701-16710.
-
Lagozzi M, Mizzoni D, Bocca B, Di Raimo R, Petrucci F, et al. (2019) Human primary macrophages scavenge AuNPs and eliminate it through exosomes, A natural shuttling for nanomaterials. Eur J Pharm Biopharm 137: 23-36.
-
Sanchez Purra M, Roig Solvas B, Rodriguez Quijada C, Leonardo BM, Hamad Schifferli K (2018) Reporter selection for nanotags in multiplexed surface enhanced Raman Spectroscopy assays. ACS Omega 3(9): 10733- 10742.
-
Muller L (2020) Exosomes: nanodust? HNO 68: 56- 59.
-
Shao HJ, Chung J, Issadore D (2016) Diagnostic technologies for circulating tumor cells and exosomes. Biosci Rep 36(1): e00292.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells