Effect of 17α-Ethinylestradiol on the Intestinal Calcium Carbonate Excretion Rate of Yellow Fin Sea Bream (Acanthopagrus latus)
Marine fish maintain their hypotonic body fluids by drinking seawater, excreting excess salt, and retaining water. Calcium in seawater is precipitated as CaCO3 in marine fish gut to limit the Ca2+ availability for absorption from the ingested water to maintain Ca2+ homeostasis; this process is controlled by endocrine functions. This study was aimed to investigate the effect of the exposure to synthetic estrogen; ethinylestradiol (EE2), on the osmoregulation process (CaCO3 excretion rate from the intestine) and reproduction in the native fish species of Kuwait. The fish were exposed in a semi-static system to three treatments (control and 5 and 10 ng EE2/l of seawater) for two weeks, and then the terminal sampling was carried out. Blood samples were taken for vitellogenin (Vtg) analysis using Western blotting, dot blot, and immunohistochemistry (IHC). Calcium carbonate samples were analysed for the excretion rate and showed no significant difference in treatments indicating that no effect on osmoregulation was observed. Water samples were analyzed by the enzyme-linked immunosorbent assay (ELISA) for the concentration of EE2 in the exposure tanks, showing a 50% decrease in the nominal concentration after 24 h of exposure. Fish gonads and liver were dissected for histological examination, showing no obvious intersex in control and lowdose exposed group, whereas the high-dose exposed group showed signs of intersex. No Vtg was induced in the plasma and that could be due to many factors such as low concentrations received in the exposure tanks (2.3 and 4.9 ng/l), fish species, and exposure period. The hepatosomatic index showed no significant difference between treatments and the gonadosomatic index showed a significant difference between the control and high-dose group. Interesting results were obtained from IHC, where Vtg was localized in both treatments, although more Vtg was detected in higher-dose exposed fish for both tissues (liver and gonads).
Al-Jandal N* and Azad IS
Environment and Life Sciences Research Center, Kuwait Institute for Scientific Research, Kuwait *Corresponding author: N Al-Jandal, Environment and Life Sciences Research Center, Kuwait Institute for Scientific Research, Kuwait; Email: njandal@kisr.edu.kw
Introduction
The endocrine-disrupting chemicals are known as environmental estrogens, estrogenic chemicals, estrogen mimics, or xenoestrogens and are defined by the EU Commission as “an exogenous substance that causes adverse health effects in an intact organism, or its progeny, consequent to changes in endocrine functions” Jones L, et al. [1]. They are thought to be largely derived from those naturally excreted by women, and/or synthetic estrogen-like molecules such as 17α–ethinylestradiol (EE2) from the contraceptive pill Tyler CR, et al. [2].
Environmental estrogens originate from a variety of sources including sewage treatment plant (STP) effluents and adverse physiological effects (endocrine disruption) have been observed in several fish species sampled downstream of STP discharges. It is well known that the presence of estrogenic compounds such as 17α–estradiol (E2) and the synthetic EE2 in surface waters is the consequence of municipal sewage discharges Garriz A, et al. [3]. In the treatment plant, bacterial may activate EE2 into its free form Al-Ansari AM, et al. [4]; Atkinson SK, et al. [5]. During the activated sludge process, EE2 persists stable and avoids breakdown and elimination Forrez I, et al. [6]; Zuo Y, et al. [7]. It is considered as the primary contaminant that contributes to the estrogenic activity in surface waters in the United Kingdom [8]. Furthermore, it is one of the most potent and significant xenoestrogens that has been detected in the domestic sewage ranging from 0.2 to 7ng/l, and up to 5ng/l in surface waters Desbrow C, et al. [8]; Belfroid AC, et al. [9]; Larsson DGJ, et al. [10]; Ternes TA, et al. [11].
One of the most widely used biomarkers of exposure to estrogens and their mimics in fish is the induction of vitellogenin (Vtg) [12, 13]. Vitellogenin is a yolk precursor lipophospho protein in the oviparous vertebrates [14], which is produced by the female liver stimulated by estrogen from the ovaries, and it is considered as a high molecular weight (250 to 600 kD) glycophospholipoprotein [15]. Vitellogenin has repeatedly been reported to be normally present in the serum of mature females only. However, after artificial stimulation, both males and immature females are capable of producing Vtg [16], and injection or feeding of xenoestrogens as well as exposure via water results in Vtg production in male as well as in female oviparous vertebrates [17, 18, 19]. Although EE2 detected in the environment is present in nanograms per liter, these concentrations are high enough to induce plasma Vtg when exposed to marine organisms under laboratory conditions [20, 21]. Fish feminization is induced by EE2 concentrations as low as 0.1 ng/l in male rainbow trout [16]. In a full life cycle of zebrafish (Brachydanio rerio), exposure via water at 1.67 ng/l EE2 caused Vtg induction Segner H, et al. [17]. It has been reported that long-term exposure of newly hatched roach (Rutilus rutilus) to 4 ng EE2/l resulted in the sex reversal of the male population [22]. Indeed, in laboratory and field studies, EE2 can cause estrogenic effects when present in the range of <1 to 5 ng/l [23, 24]. However, higher levels have been documented in Canadian sewage outflows [11] and once discharged from WWTPs, they may come in contact with aquatic life revealed that E2 and EE2, at environmentally relevant concentrations [3, 10], did not affect sperm quality and fertilization percentage in pejerrey (Odontesthes bonariensis) when tested separately. However, when these two estrogens were combined, they had the potential to reduce sperm motility thus negatively affecting the percentage of fertilization of pejerrey fish.
In fact, the presence of Vtg in liver and serum of male fish may be used as a biomarker of exposure to xenoestrogenic chemicals Pelissero C, et al. [25]; Heppell SA, et al. [26]; Palmer BD, et al. [27]; Bieberstein U, et al. [28] reported that the induction of Vtg in the liver of estrogen-induced male and female rainbow trout (Oncorhynchus mykiss) was visualized by immunohistochemical and immunocytochemical techniques in both frozen and paraffin-embedded tissues. The detection of Vtg served as a positive control. Localization was accomplished using an indirect immunofluorescence method.
The potential for natural estrogen to control the production of intestinal calcium carbonate (CaCO3) was investigated in seawater-acclimated rainbow trout (Oncorhynchus mykiss) following intraperitoneal implantation of E2 at two doses (0.1 and 10 µg E2g–1), the results revealed that the intestinal excretion of calcium carbonate was reduced Al-Jandal N, et al. [2]. The results obtained in the previous study led to further investigation on the effect of EDCs on fish osmoregulation. In the present study, the experiment was conducted in the native fish species of Kuwait to show the effect of the exposure to synthetic estrogen EE2 on the excretion rate of CaCO3 and on the reproductive process. The fish were exposed in a semi-static system to two doses, 5 and 10 ng EE2/l seawater, along with controls, for two weeks and various effects parameters were analyzed.
Materials and Methods
Exposure Regime and Terminal Sampling
Immature male yellow finned seabream (Acanthopagrus latus) (n=36; 227.6±3 g; 21.7±0.1 cm fork length) were obtained from the Aquaculture Program facilities at Kuwait Institute for Scientific Research (KISR). Fish were kept in aerated seawater (pH 7.5; 32.0±0.2 °C), then randomly allocated to one of the three treatments (12 fish per treatment) and maintained in their individual chambers during the experiment. Fish were exposed to waterborne EE2 (control, 5 ng/L, and 10 ng/L) in a semi-static system for two weeks, where 50 % of the water in each chamber was renewed every 48h and re-dosed with the required volume of seawater and EE2 concentration. Fish were starved for two days pre- and during exposure, and blood samples were taken via caudal puncture prior to exposure for the analysis of plasma Vtg. After two weeks of exposure, all fish were terminated for analysis.
Determination of 17α-ethinylestradiol in Exposure Tanks
Water samples were taken from the exposure tanks at 0 h and after 24 h to measure the nominal and actual concentration of EE2. Water samples were stored at 4ºC until analysis using two different methods, the liquid chromatography/mass spectrometry (LC-MS) and enzyme- linked immunosorbent assay (ELISA).
Water Sample Extraction Method
Water samples were extracted prior to analysis using the SPE–DEX 4790 Automated Extraction System (Horizon Technology SPE–DEX), according to Kelly C, et al. [29]. The Disc was preconditioned by rinsing with 5 ml of methanol followed by 10 ml distilled water to remove any potentially interfering compounds. The filtrate was poured through the system, followed by washing with 5 ml distilled water. Each disk was then allowed to dry, prior to washing with 5ml hexane, by suctioning for about one minute to dry the cartridge. EE2 was then eluted by the addition of 5 ml dichloromethane, and the aliquots were collected and combined in a salinized glass tube. All samples were evaporated using a Turbovap evaporator (Zymark, Warrington, UK), where nitrogen gas was supplied through Pasteur pipettes into 10ml tubes. Temperature was controlled at 40-50°C with water bath to accelerate the evaporation process. The residue of 1l filtrate was suspended by the addition of 100 µl of 100 % methanol and the mixture was stirred very well with a vortex. The mixture was terminated by pouring 900 µl distilled water to adjust the content at 10% methanol (v/v).
Liquid Chromatography–Mass Spectrometry (LC-MS)
Samples were filtered through 0.45 µm nylon membrane filters (Whatman, UK), and 1 g sodium azide was added to each bottle as a preservative. An additional 25 mg/L of ascorbic acid was added to potable and seawater as a quenching agent. The samples were stored at 4oC prior to extraction. The samples were spiked with 13C–Ethinyl estradiol and extracted by offline phase extraction with an automated wastewater extraction unit (Horizon Technologies, USA) using an Oasis HLB cartridge (200 mg and 6 ml).
Mass spectrometry was performed using Waters ACQUITY TQ-S triple quadrupole mass spectrometer equipped with an electrospray source. Both labeled and unlabeled EE2 were infused into the mass spectrometer to optimize the multiple reaction monitoring transitions and to determine their retention times using various gradient programs. Subsequently, a set of runs with different gradient settings were carried out to optimize the chromatographic separation and peak shape and simplify detection in the chromatographic areas where potential superposition was expected. The linear dynamic range of the instrument response was then verified by injecting (at the optimized gradient conditions) a set of standards containing both labeled and unlabeled EE2 at the following concentrations: 10, 20, 40, 50, 60, 80, and 100 ppb. It was observed that the detector was about 100 ppb, which resulted in a loss of response linearity.
Water samples were analyzed also by using EE2 ELISA Kit (Ethinylestradiol Elisa Kit, Ecologiena, and TOKIWA Chemicals Industries). EE2 monoclonal antibody binds exclusively with EE2 and does not show a cross–reaction with other hormones or chemicals with similar structures. The quantitative analysis ranges from 0.05 μg/L to 3 μg/L, sensitive enough to detect EE2 in a specimen from river water, blood, etc. With ease of handling, the total time for measurement is only 2.5 h. The ELISA measurement is highly reproducible; the coefficient of variation (CV) is generally under 10%. The kit, in a 96–well microplate format, enables simultaneous measurement of multiple samples at a more reasonable cost. Water samples from the exposure tanks were pretreated after measuring the pH (7.5). According to the procedure, the pH of the filtrate should be between 5 and 8, and according to the Kit manufacturer’s protocol, the C18 cartridge was preconditioned by the addition of 5 ml of methanol and then 10 ml of distilled water. To save time, suctioning with a vacuum pump at a flow rate of 20 ml/min was recommended. The filtrate was poured through the C18 cartridge at the same flow rate. The cartridge then washed by 5 ml of distilled water for about a minute to dry, and then washed with 5 ml of hexane. The EE2 (analyte) was eluted with 5 ml of dichloromethane at a slow flow rate (3 ml/ min). The solvent was evaporated with compressed air at a temperature between 40–50 °C to accelerate the evaporation of the organic solvent. A Hundred percent methanol was added to the residue and stirred with a vortex. The residue was then suspended by 10% v/v, and the mixture was stirred with a vortex. Samples containing EE2 within the dynamic range (0.05-3 μg/L) can be directly applied to the assay after filtration. Samples with EE2 content below the range must be concentrated with solid phase extraction prior to the ensuing session. The CV was generally under 10% throughout the dynamic range.
Vitellogenin Analysis using Immuno histochemistry
Immunohistochemistry (IHC) was used to localize Vtg in the tissue samples. Paraffin- embedded tissues, gonad and liver, were sectioned and collected onto coated (1% lysine) slides, dried on a thermostat stage, and held overnight in an incubator at 35 °C. Dewaxed, hydrated sections of liver and gonad were used in immunofluorescence staining. Briefly, the sections were treated with trypsin (0.1% w/v in phosphate buffered saline (PBS), biochemical grade, Sigma Aldricht) for 10 min and washed twice with PBS–Tween (PBST) 20 (T-20, Fluka, Switzerland; 0.01% v/v with PBS). Nonspecific antibody binding sites were blocked for 30 min using 3% bovine serum albumin (in PBS, pH 7.4) and washed with PBST. The sections were treated with the first antibody (rabbit anti-sea bream vitellogenin, Cayman Norway) at a dilution of 1:1000 (sterile PBS, pH 7.4) for 1 h in a humid chamber, while the control sections were incubated with normal unimmunised rabbit serum of an identical dilution. Sections were washed with PBST before treating with goat anti-rabbit FITC conjugate (1:40 dilution with distilled water) for 1 h. The sections were again washed with PBST before lightly blotting and mounting using glycerol. Gel Electrophoresis Western blotting combined gel electrophoresis was used to separate the proteins in the microsome fractions with the specificity of immunological detection. Gel electrophoresis was carried using 8 % sodium dodecyl sulphate-potyacrylamide gels (SDS-PAGE) according to the method of Schägger H [30]. A rubber gasket was fixed on the edge of the glass plate. Spacers were then placed on the plate in which the ridge of the gasket was directed upwards. The second flat glass was placed on top and clipped together. The stock solutions prepared for gel electrophoresis are given in Table 1.
| Buffers | Tris* (M) | Tricine (M) | pH | SDS (%) |
|---|---|---|---|---|
| Anode buffer | 0.2 | - | 8.9 | - |
| Cathode buffer | 0.1 | 0.1 | 8.25 | 0.1 |
| Separating buffer | 3 | - | 8.45 | 0.3 |
| Stacking buffer | 1.64 | - | 7.5 | 0.3 |
Table 1: Stock Buffer Solution for SDS-PAGE. (*) adjusted with HCl (1 M); SDS-PAGE: sodium dodecyl sulphate-potyacrylamide gels.
All solutions were kept at room temperature with the exception of the bisacrylamide mixture, which was stored at 4 °C. The composition of separating and stacking gels is given in Table 2. The 8% acrylamide (29: 1 acrylamide to bisacrylamide) was used as a uniform separating gel. All the components of the gel were mixed together in a Buchner flask and degassed, except tertamethylethylenediamine (TEMED) and 10% ammonium persulfate (APS) (must be fresh), which were added at the end. Gently, the flask was swirled to mix and the gel was cast carefully (~ 18 cm × 22 cm plate) using a glass syringe connected to 1-mm diameter tube to prevent air bubble generation. Ethanol was added (gently) on the top of the separating gel to give a very sharp liquid interface (visible within 10 min). The separating gel was left to polymerize for at least 1 h. After the termination of the polymerization, the surface of the gel was rinsed with distilled water before the pouring of 3% stacking gel (1 to 2 cm). The plate was filled with stacking gel solution and a comb was inserted into the gel by taking care not to trap any bubbles below the teeth.
| Solutions | Separating Gel | Stacking Gel |
|---|---|---|
| 8 % bisacrylamide (ml) | 3 % bisacrylamide (ml) | |
| 40 % bisacrylamide | 7 | 1.125 |
| Gel buffer | 11.67 | 3.72 |
| TEMED | 0.015 | 0.01 |
| 10 % APS | 0.15 | 0.1 |
| H2O | 16.165 | 10.045 |
Table 2: Composition of Separating and Stacking Gel. TEMED: Tertamethylethylenediamine; APS: Ammonium Persulfate
The samples were diluted in a ratio of 1:3 volume of the sample buffer (4% SDS, 12% glycerol, 50 mM Tris, 2% mercaptoethanol, 0.01% bromophenol blue R adjusted with HCI to pH 6.8). The protein samples were then incubated for 30 min at 40 °C. The comb and rubber gasket spacer were removed from the gel gently and the wells were rinsed with distilled water and the water was drained off. The plate set was assembled in the vertical apparatus for running the electrophoresis (Sigma, Z37, 507-1). The upper chamber and the wells were filled with a cathode buffer (10 X Tris-tricine- SDS buffer). A sufficient quantity of anode buffer was poured into the lower chamber of the electrophoresis apparatus until the bottom of the gel was immersed in buffer taking care to eliminate bubbles from the bottom of the gel, and the top was covered, while the electrodes reached into the buffer of the upper chamber. The sample was loaded to the bottom of a well. The top of the electrophoresis apparatus was assembled and connected to the power source (cathode (+) is connected to the upper buffer chamber). The electrophoresis was performed at room temperature and the run was started at 60 V. After about 30 min, when the sample had completely entered the stacking gel (or left the sample well), the voltage was raised to 140 V for 2.5 h. The gel apparatus and the glass sandwich were disassembled, and the sandwich was placed carefully on paper towels and the clamps were removed from the sandwich. The bottom glass plate with the gel was lifted and the gel was transferred to an appropriate container filled with a fixing buffer.
The compositions of all the solutions are listed in Table 3. The protein bands were fixed in a fixing solution for 30 min before they were stained with a staining solution for at least 2 h (or overnight) by shaking with a rocker unit. A complete background of the gel was destained in two steps: by shaking the gel in the destain II solution for two hours, the destaining solution being exchanged every half an hour; followed by, destaining the gel in destain I solution, and then storing the gel in the preservation solution.
| Solutions | Acetic Acid (%) | Methanol (%) | H2O | Glycerol (%) | Brilliant Blue R (%) |
|---|---|---|---|---|---|
| Fixing | 7 | 40 | 530 | - | - |
| Staining | 10 | 45 | 450 | - | 1 |
| destain II | 10 | 10 | 800 | - | - |
| destain I | 10 | 50 | 400 | - | - |
| Preservation | 10 | - | 770 | 13 | - |
Table 3: Stock Solutions for Fixing, Staining, and Destaining.
Measurement of Vitellogenin by Western Blot
Western blot analysis was run with rabbit polyclonal antibodies against sea bream Vtg. The primary antibody was purchased from Biosense Laboratories AS, Bergen, Norway, to detect the Vtg protein in fish plasma. The immunological method requires the use of specific antibodies that specifically bind the target Vtg. To do this, the project team tried to measure Vtg- immunopositive protein using primary antibody apparently similar to the local fish (Acanthopagrus latus). The transfer process of the protein and antibody activity was checked by dot blot tests to check the efficiency of the Western blot procedure.
Two tests were carried out to determine if the antibodies and the detection system were effective. The first test was run to check the efficiency of the Western blot steps, in which the blotted membrane was stained with 0.5 % 3, 3’-Diaminobenzidine (DAB) solution for 1 min to check whether all protein bands have successfully transferred to the membrane. To check the binding of the primary antibody to the Vtg standard, another test was undertaken where 2 µl of seven Vtg standards, (16.67, 8.3, 4.2, 2.1, 1.0,
0.5, and 0.26 ng/PBS µl) were spotted onto the membrane and incubated for a few minutes at room temperature. The blot was allowed to dry (30 min) before going to the next step. Then, the nonspecific binding were blocked with 5% bovine serum albumin (BSA) in Tween20 buffer for 1 hour at room temperature and washed three times (10 minutes each) with Tween20 buffer to remove excess 5% BSA from the membrane. After washing the primary antibody (1:1000, diluted in 5% BSA) was added and incubated for 30 min. Again, the membrane was washed three times in Tween20 buffer before adding the secondary. Antibody (goat anti rabbit horseradish peroxidase (IgG- HRP) conjugate, cat HPO3/Lot 31125). The excess secondary antibody was also washed three times with Tween20 buffer after 30 min of incubation and the dot was detected by the DAB solution.
Hepatosomatic Index and Gonadosomatic Index
The total organ weight (liver and gonads) was taken for the hepatosomatic index (HSI) and the gonadosomatic index (GSI). The indices were calculated according to the following formula:
HSI = (Liver weight (g) / Fish weight (g)) × 100 (1) GSI = (Gonad weight (g) / Fish weight (g)) × 100 (2) As the liver is known to be the target organ for the metabolism in the fish body, the liver HSI is a useful biomarker to detect the hazardous effects of the environmental stressors. In this study, the weight of the liver and the gonads were taken from each fish in different treatments and related to the total body weight. GSI is a highly comprehensive index to assess the damage of the reproductive function at the tissue level. In general, the exposure to environmental estrogens always results in a pronounced decline in the GSI of adult male fish Jobling GP, et al. [12]. These significantly lower GSIs are likely to be a result of Vtg synthesis. Jobling, et al. reported the inverse correlation between plasma Vtg and GSI in male roaches (Rutilus rutllus). Considering that Vtg synthesis is an energy-wasting process, the reproduction and growth of exposed fish tend to be delayed.
Histology
Fish terminal sampling was carried out, all fish samples were dissected, and gonads and liver were removed for histology and sex confirmation. Gonads and liver were fixed in 10% formalin for 24 h and then in 50% propanol. Tissue samples were processed by Shandon processor, from 50% propanol to molten wax through a series of alcohol and xylene. After processing, tissues were and embedded with wax by histo-embedder (Lieca Biosystems). Tissue blocks were sectioned using a microtome, and sections were transferred onto the slides for staining. Slides were stained with haematoxilin and eosin and mounted using DPX.
Calcium Carbonate Excretion Rate
Intestinal precipitates of calcium carbonate excreted into the chamber were collected twice daily morning and night from each fish. The excretion rate of CaCO3 from dry weight was calculated as follows:
Dry weight = Dry wt of ppt (g) ∕ Fish mass (kg) / Time (h) Dry samples of CaCO3 were also analyzed at KISR to measure the _C_a2+% as CaCO3 content in the part per thousand (ppt) using Energy–dispersive X–ray Spectroscopy (EDX).
Statistical Analysis
Significant differences between treatments will be determined using one way Analysis of Variance (ANOVA), and differences between treatment groups will be considered to be significant when P< 0.05. All statistical analysis will be conducted using Sigmastat 3.5 (Systat Software, Inc.).
Results
Exposure Regime and Terminal Sampling
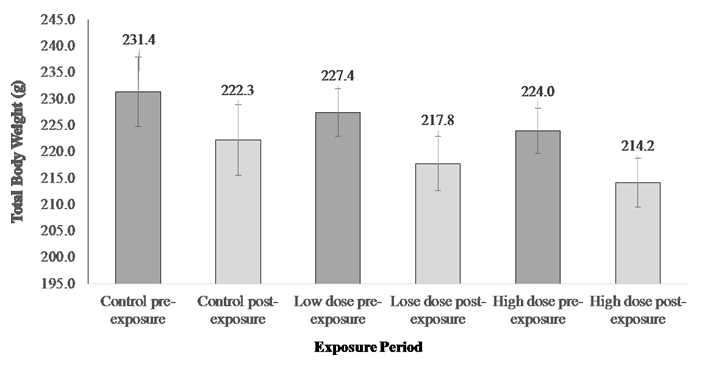
Using the semi-static exposure system or the renewal exposure system was a compromise between the flow through system and the static system in this study. After two weeks of exposure, the experiment was terminated and terminal sampling was carried out. The body measurements were taken pre- and post- the exposure. The results showed a trend of decrease in the total body weight (≈5%) after two weeks of exposure in all groups, although the difference in weight pre- and post- the exposure is not significant different (Figure 1).
Determination of 17α-ethinylestradiol in Exposure Tanks
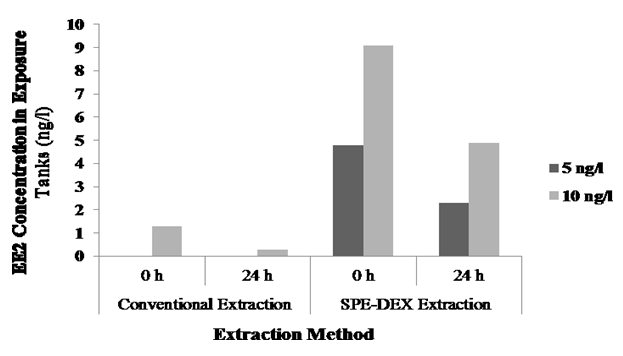
The exposure conditions were designed to stimulate Vtg induction in the local species of Kuwait (yellow-finned seabream) by exposing them to waterborne EE2 (5 and 10 ng/L) for two weeks. Measuring EE2 level in the water was done using the LC-MS technique; however, the levels in the samples recorded were below the detection limit. Therefore, ELISA technique was carried out for water samples at 0 h and after 24 h. The results of EE2 levels obtained by the conventional extraction method were considerably different from the expected, in which the reduction of EE2 was reported in all water samples tested (Table 4). Nevertheless, EE2 level was recorded only in the highest exposure level sample.
| Extraction Method | Initial Concentration (ng/l) | Actual Concentration (ng/l) | Recovery Percentage (%) |
|---|---|---|---|
| Conventional Extraction | 5 ng/l (0 h) | Below the detection limit | NA |
| 5 ng/l (24 h) | Below the detection limit | NA | |
| 10 ng/l (0 h) | 1.3 | 13 | |
| 10 ng/l (24 h) | 0.3 | 3 | |
| SPE–DEX Extraction | 5 ng/l (0 h) | 4.8 | 96 |
Table 4: Ethinylestradiol Recovery after the Extraction Method.
Standardization experiments showed that the extraction of EE2 from water sample using SPE–DEX 4790 extractor system results in a much better recovery of EE2 in the water samples (Table 3) than to the conventional method. The results indicated ~50% loss of EE2 after 24 h of exposure for both treatments (Figure 2).
Vitellogenin Analysis by using Immunohistochemistry
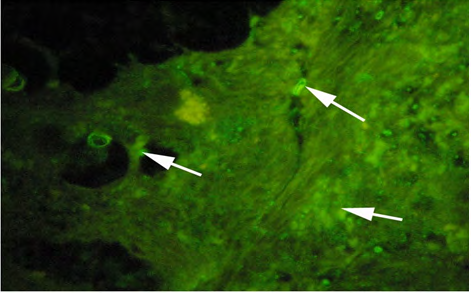
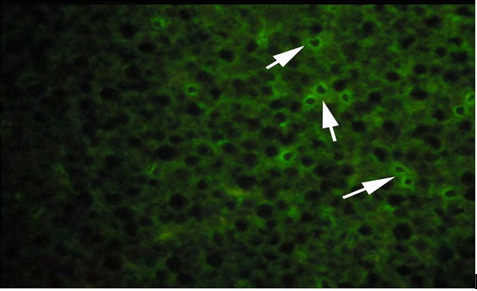
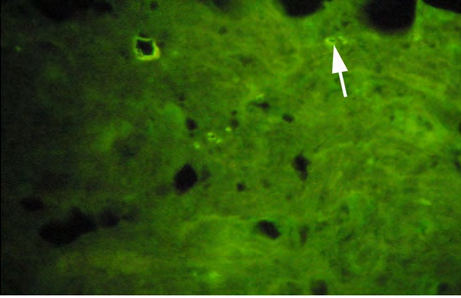
Vitellogenin in plasma samples were measured by immunolocalization using antibody- based technique (IHC) in the tissue (liver/gonad) sections revealed of yellow-finned seabream. Unexposed (control) and exposed to low dose (5 ng/l) and high dose (10 ng/l), showed a clear response to EE2 exposure by the fish. The control fish tissue showed weak or no localization of Vtg compared with a mild but consistent localization in the fish tissue exposed to the low dose, while there was a strong immunefluorescence in immmunohistochemicals labeling present in the liver tissue of fish exposed to the high dose. The response of the liver tissue was more consistent and intense compared with that of the gonad tissues. The results of immunofluorescence in the liver of control showed no signs of labeling or weak positive labeling only (Figure 3a) was noticed indicating the background levels of Vtg. Significant positive reaction was associated with the intra hepatocytic and perinuclear regions of the tissue of fish exposed to low-dose EE2 group (Figure 3b) and similar but highly reactive localization was noticed in the liver tissue (Figure 3c) of the fish exposed to high-dose EE2.
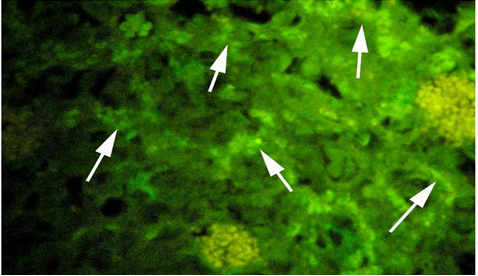
The gonad tissue from the control fish revealed extremely lower positive fluorescent labeling (Figure 4a) compared to the gonad from the fish exposed to EE2 (Figures 4b and 4c). However, the gonad sections of fish exposed to high-dose EE2 showed strong but diffused reactivity.
(A) Control fish
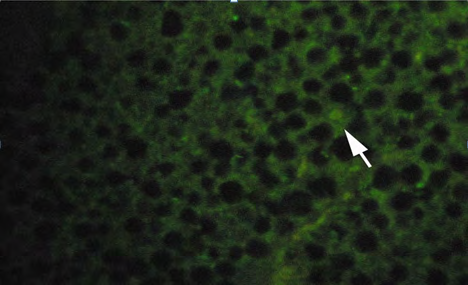
(B) Low dose (5 ng/l) exposed fish.
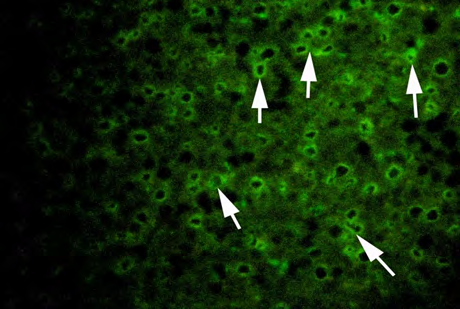
(C) High dose (10 ng/l) exposed fish Figure 3: Plates showing immunohistochemistry technique used in the liver for Vtg localization.
(A) Control fish.
(B) Low dose (5 ng/l) exposed fish.
(C) High dose (10 ng/l) exposed fish. Figure 4: Plates showing immunohistochemistry technique used in the gonad for Vtg.
Vitellogenin Analysis using Western Blot and Dot Blot
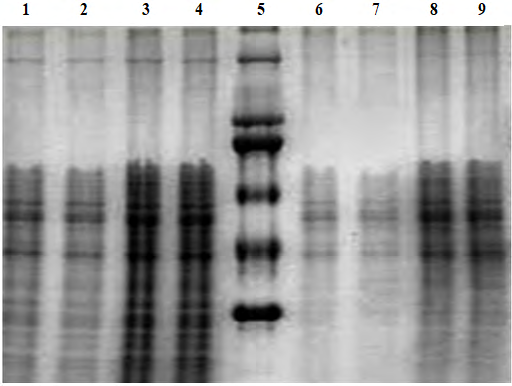
Western blot analysis was run to detect the Vtg protein in fish plasma. Vtg protein was detected in the previous preliminary study carried out by Prof. R. Abukensha from King’s College of London. The study (SDS-PAGE coupled to Western blot tests) was conducted in KISR’s laboratory on the available universal antibody, and the results showed clear evidence that this universal antibody recognizes and reacts with Vtg proteins in several of Kuwait’s priority local fish species, and a reactive band of 250 kDa in the plasma of the fish was detected (Figure 5). The immunological method requires the use of specific antibodies that specifically bind the target Vtg. Therefore, Vtg-immunopositive protein was measured by using a primary antibody apparently similar to the local fish (Acanthopagrus latus). A band of 230 kDa was observed in the SDS-PAGE (Figure 6), but Western blot assay did not show cross-reactivity with any band on the nitrocellulose membrane. Also, there was no positive signal for Vtg in any of the treatments (low and high).


The transfer process of the protein and antibody activity was checked by dot blot tests to check the efficiency of the Western blot procedure. Three tests were carried out to determine if the antibodies and the detection system were effective. The first test was run to check for the efficiency of the Western blot steps; all bands were detected indicating that the blotting procedure was effective. The binding of the primary antibody to the Vtg standard was checked, and the dot appeared only in the first four Vtg standard concentrations (i.e., 16.67, 8.3, 4.2, and 2.1 ng/PBSµl) and none of the plasma samples has showed positive reaction with the DAB solution. The detected spot confirmed the binding of the primary antibody to the Vtg protein. In the last test, 1 µl of secondary antibody-HRP activity was mixed with the DAB solution producing the brown color, which confirmed the activity of the conjugate (Figure 7).
Histological Analysis
Histological analysis results showed no obvious signs of intersex observed in both the control group and the low- dose exposed fish. Whereas, the high-dose exposed fish (10 ng EE2/l of seawater), showed a different trend in more than one slide. There were signs of intersex showed (Figure 8).
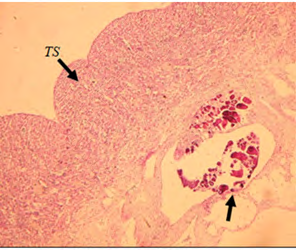
A) Cross section of male testicular tissue (×5) showing signs of intersex (arrow).
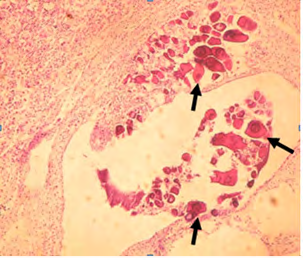
B) Higher magnification (×40) showing the perinuclear oocytes (arrow)within the transverse section.
C) Cross section of the testicular tissue (×20) the showing perinuclear oocytes scattered in the tissue in another male fish from the same group.
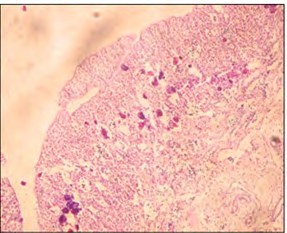
D) Higher magnification (×40) showing perinuclear oocytes (arrows). Figure 8: Light micrograph of high-dose exposed fish group.
Hepatosomatic Index and Gonadosomatic Index
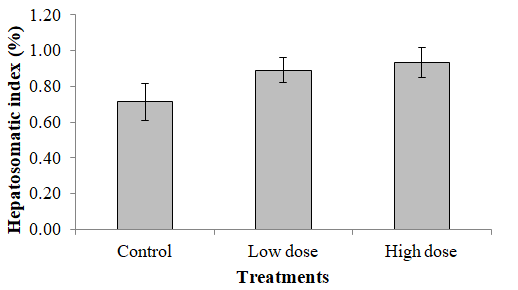
The results of HSI revealed no significant difference in the different treatments as the P values were found to be > 0.05, although there is a trend of elevation in the value (Figure 9).
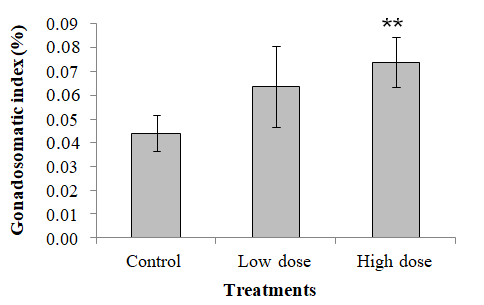
In the case of the GSI, the results obtained from this study revealed that there were some treatment related differences in EE2 treated fish. Results showed that there was a significant increase of gonad growth in high-dose EE2 treated group (0.074 %) in comparison to the control group (0.044 %), as expressed by the GSI (Figure 10).
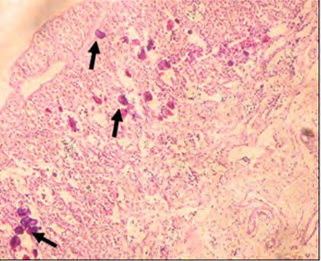
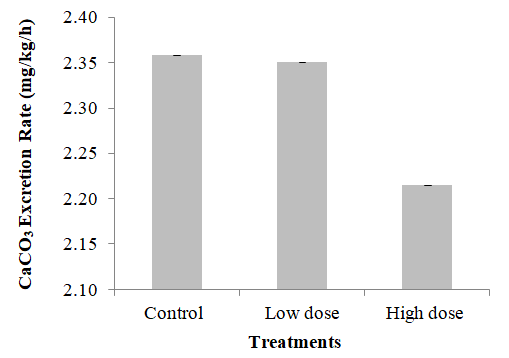
Figure 10: Gonadosomatic index of sea bream exposed to EE2 for two weeks. Calcium Carbonate Excretion Rate The results obtained after two weeks of exposure to EE2 revealed no significant difference between treatments. The control group excretion rate was 2.36 mg/kg/h, the low-dose group was 2.35 mg/kg/h, and the high-dose group rate was 2.21 mg/kg/h (Figure11).
Although the result showed a trend of decline in the excretion rate, but the P value was more than 0.05.
Discussion
Exposure Regime and Terminal Sampling
The reason for using semi-static system in this study due to multiple advantages;
- This system can be mainly used for the small organisms,
- It was the best system to be used for collecting CaCO3 on a daily basis without worrying about flushing it out when using the flow though system,
- This system is useful when limited amount of test materials are available and a prolonged test is required. There are some disadvantages that should be considered also such as the frequent handling of the test organisms that increases stress and organisms’ injury, the test material concentration may not be constant throughout the experiment, and it is more labor- intensive than the static [31].
During the two weeks of exposure, fish feeding was stopped to avoid any mixing between the excreted CaCO3 and faeces, therefore the trend of declining in the total body weight in each group observed after two weeks of exposure was due to starvation.
Determination of 17α-Ethinylestradiol in Exposure Tanks
There are possible reasons for the observed reduction in the EE2 level in the water samples. Most of the loss in concentration recorded might be immediate, prior to 0 min sampling, indicating an adsorption of EE2 to the glass walls of the experimental containers, exposure tanks, and extraction procedure for concentration analysis since EE2 is described as sticky.
Vitellogenin Analysis by using Immuno histochemistry
In general, Vtg synthesized in the liver is transported via the blood to the ovary where it is incorporated and sequestered by the maturing oocytes Arukwe A, et al. [32]. An increased synthesis of Vtg in the liver could be a reason for such existence in liver tissue of exposed fish. Similar observations to our results were noticed in the rainbow trout (Oncorhynchus mykiss) injected intraperitoneally with E2, where localization was observed in the perinuclear regions due to the intensive accumulation of rough endoplasmic reticulum Bieberstein U, et al. [28]. In another study an exposure of juvenile fathead minnow (Pimephales promelas) to 2, 5 and 20 ng EE2/l induced a dose-related response in Vtg concentrations at 7, 14 and 21 days of exposure Panter GH, et al. [32]. Zebrafish (Danio rerio) that was exposed to a nominal concentration of EE2 (2 ng/l) had significantly elevated concentrations of Vtg in the whole body Örn S, et al. [33].
The present study revealed that the IHC technique is a good tool to investigate the effect of estrogen exposure and a suitable tool to indicate Vtg in liver hepatocytes. The immune positive staining of gonads and liver indicated the ability of antibodies to detect Vtg in this study. It has been reported that after the administration of a single intraperitoneal injection of E2 dissolved in 100% ethanol (1mg/kg body wt), Vtg could be detected in the hepatocytes of male rainbow trout with intensity almost similar to that in the liver of female individuals Bieberstein U, et al. [28].
Vitellogenin Analysis using Western Blot and Dot Blot
From the earlier tests and results obtained, it is concluded that there was no problem with either the binding of the Vtg protein to the primary antibody or the binding of the secondary antibody to primary antibody. The lack of a statistically significant difference in Vtg level or induction might be attributed to a high variability in Vtg in the reference male fish [34, 35]. The reason might be attributed to the level of exposure recovered too.
Histological Analysis
It is generally reported that the histopathological biomarkers are useful as indicators of the general health of the fish and are considered as a mirror that reflects the exposure to a variety of anthropogenic pollutants [36].
Many studies reported the intersex phenomena in different marine organisms when exposed to ECDs, especially estrogens. Intersex is defined as the simultaneous presence of male and female gonadal tissue in an individual of a gonochoristic (fixed-sex) species Tyler CR, et al. [37]. Chang CF, et al. [38] reported that the induction of sex reversal by an oral administration of E2 to two-year old black porgy (Acanthopagrus schlegeli) and the higher levels of plasma E2 in the sex reversed fish further support the importance of E2 in the sex change of protandrous black porgy . In another study, Bjerregaard LB, et al. [39] reported that male roach (Rutilus rutilus) from Danish streams that receive discharges of domestic sewage effluent, intersex was observed at a prevalence of 4.5 to 5% at reference sites and 6.7 to 6.5% at sewage effluent-impacted sites. The primary objective of a study done by Tetreault GR, et al. [40] was to find out the occurrence of intersex condition and reproductive disruption present in wild fish exposed to multiple sewage effluent discharges in a system with limited surface water for dilution. Male testicular tissue was scanned for the presence of eggs and intersex was observed, whereas only the previtellogenic oocyte stage was observed in the intersex fish. Although many studies have measured elevated levels of Vtg in male fish and reported incidence of the intersex condition when exposed to sewage treatment plants effluent, few studies have developed a strong correlation between Vtg concentration and the presence of intersex. Substantive evidence has been documented to support the hypothesis of estrogenic compounds contributing to the cause of intersex in exposed fish; however, estrogen exposure has correlated less with plasma Vtg in the same fish [41].
Analysis of Vtg was consistent with the hypothesis that the intersex observed in white perch (Morone americana) was the result of exposure to estrogenic endocrine disrupting substances; however, this study did not correlate Vtg concentration with the degree of intersex as all exposed males displayed some measurable plasma Vtg [42]. Similarly, the incidence of intersex in white sucker could not be consistently correlated with contaminants Hinck JE, et al. [43]. In the present study, this might be the case where no Vtg was induced in the plasma although intersex was present in the gonads of the highly exposed group of fish.
Hepatosomatic Index and Gonadosomatic Index
Some studies reported a significant decrease of the HSI in rainbow trout (Oncorhynchus mykiss) injected with paraquat after nine weeks [44]. In contrary, Figueiredo-Fernandes A, et al. [45] found an increase of HSI in male and female tilapia, O. niloticus, exposed to paraquat. Barse AV, et al. [46] reported elevated HSI values of Cyprinus carpio subjected to 4-tert-butylphenol. This lack of significance might be due to the short exposure time of the experiment that caused effects on the subcellular structural and biochemical level and was likely to be too short to capture effects on growth and/or development. Similarly, Hecker M, et al. [47] observed greater GSI values in African clawed frogs (Xenopus laevis) caught from an area with elevated aqueous atrazine concentrations.
Calcium Carbonate Excretion Rate
The results obtained followed the trend observed in one of the studies Al-Jandal N, et al. [2] in which elevated levels of circulating E2 was related to enhanced Ca2+ uptake via the gut and simultaneously reduced the CaCO3 formation and excretion. The reason that might have affected the results was the difference in nominal and actual concentration of EE2 in the exposure tanks, and the reduction in exposure concentration to ~50% in comparison to the initial concentration which resulted in non-significant result although the declining trend was observed [48, 49].
Conclusion
The present study is the first of its kind conducted in the native fish species of Kuwait on the effect of the exposure to estrogen mimics, EE2 on the osmoregulation process and on reproductive process. Studies of calcium balance have mostly focused on freshwater and euryhaline fish rather than marine fish. Our novel finding in this study that is that EE2 induces plasma Vtg in local fish when exposed for two week under laboratory conditions, and has no effect on CaCO3 production by the intestine in yellow-finned seabream of Kuwait. The potential of synthetic estrogen to influence CaCO3 excretion. The immunefluorescence immmunohistochemical labeling technique showed a concentration-dependent reactive localization of Vtg in the liver and gonad tissues of the exposed fish. Histological analysis showed that the high- dose exposed fish showed distinct signs of intersex after two weeks of exposure.
References
-
Jones L, Regan F (2019) Endocrine Disrupting Chemicals. Reference Module in Chemistry, Molecular Sciences and Chemical Engineering.
-
Tyler CR, Jobling S, Sumpter JP (1998) Endocrine Disruption in Wildlife: A Critical Review of the Evidence. Criti Rev Toxicol 28(4): 319-361.
-
Garriz A, Menendez-Helman RJ, Miranda LA (2015) Effects of estradiol and ethinylestradiol on sperm quality, fertilization, and embryo–larval survival of pejerrey fish (_Odontesthes bonariensis_). Aquatic Toxicology 167: 191-199.
-
Al Ansari AM, Saleem A, Kimpe LE, Sherry JP, McMaster M, et al. (2010) Bioaccumulation of the pharmaceutical 17α-ethinylestradiol in shorthead redhorse suckers (Moxostoma macrolepidotum) from the St. Clair River, Canada. Environ Pollut 158(8): 2566-2571.
-
Atkinson SK, Marlatt VL, Kimpe LE, Lean DRS, Trudeau VL, et al. (2012) The occurrence of steroidal estrogens in south-eastern Ontario wastewater treatment plants. Sci Total Environ 430: 119-125.
-
Forrez I, Carballa M, Noppe H, De Brabandar H, Boon N, et al. (2009) In Fl uence of manganese and ammonium oxidation on the removal of 17α-ethinylestradiol (EE2). Water Res 43(1): 77-86.
-
Zuo Y, Zhang K, Deng Y (2006) Occurrence and photochemical degradation of 17α-ethinylestradiol in Acushnet River Estuary. Chemosphere 63(9): 1583- 1590.
-
Desbrow C, Routledge EJ, Brighty GC, Sumpter JP, Waldock M (1998) Identification of estrogenic chemicals in STW effluent. 1. Chemical fractionation and in vitro biological screening. Environmental Science and Technology 32(11): 1549-1558.
-
Belfroid AC, Van der Horst A, Vethaak AD, Schafer AJ, Rijs GB, et al. (1999) Analysis and occurrence of estrogenic hormones and their glucuronides in surface water and waste water in The Netherlands. Science of The Total Environment 225(1-2): 101-108.
-
Larsson DGJ, Erici MA, Parkkonen J, Pettersson M, Berg AH, et al. (1999) Ethinyloestradiol an undesired fish contraceptive? Aquatic Toxicology 45(2-3): 91-97.
-
Ternes TA, Kreckel P, Mueller J (1999) Behaviour and occurrence of estrogens in municipal sewage treatment plants – II. Aerobic batch experiments with activated sludge. Sci Total Environ 225(1-2): 91-99.
-
Tyler CB, van der Eerden S, Jobling GP, Sumpter J (1996) Measurement of vitellogenin, a biomarker for exposure to oestrogenic chemicals, in a wide variety of cyprinid fish. Journal of Comparative Physiology B 166: 418-426.
-
Kime DE, Nash JP (1999) Gamete viability as an indicator of reproductive endocrine disruption in fish. Science of The Total Environment 233(1-3): 123-129.
-
Bergink EW, Wallace RA (1974) Precursor–product relationship between amphibian vitellogenin and yolk proteins, lipovitellin and phosvitin. Journal of Biological Chemistry 249(9): 2897-2903.
-
Specker JL, Sullivan CV (1994) Vitellogenesis in fishes: status and perspectives. In: Davey KG, et al. (Eds.), Perspectives in Comparative Endocrinology. Ottawa: National Research Council Canada, pp: 304-315.
-
Purdom CE, Hardiman PA, Bye VVJ, Eno NC, Tyler CR, et al. (1994) Estrogenic effects of effluents from sewage treatment works. Chemistry and Ecology 8(4): 275-285.
-
Wangh LJ, Knowland J (1975) Synthesis of vitellogenin in cultures of male and female liver regulated by estradiol treatment in vitro. Proc Natl Acad Sci USA 72(8): 3172- 3175.
-
Emmersen BK, Petersen IM (1976) Natural occurrence and experimental induction by estradiol–17b of a lipophosphoprotein (vitellogenin) in flounder (_Platichtys flesus_ L). Comparative Biochemisrty and Physiology 54(4): 443-446.
-
Al–Jandal N, Whittamore JM, Santos EM, Wilson RW (2011) The influence of 17β– estradiol on intestinal calcium carbonate precipitation and osmoregulation in seawater-acclimated rainbow trout (_Oncorhynchus_ _mykiss_). The Journal of Experimental Biology 214(16): 2791-2798.
-
Länge R, Hutchinson TH, Croudace CP, Siegmund F, Schweinfurth H, et al. (2001) Effects of the synthetic estrogen 17α–ethinylestradiol on the life–cycle of the fathead minnow (_Pimephales promelas_). Environ Toxicol Chem 20(6): 1216-1227. 21. Schultz IR, Skillman A, Nicolas JM, Cyr DG, Nagler JJ (2003) Short–term exposure to 17 α–Ethinylestradiol decreases the fertility of sexually maturing male rainbow trout (_Oncorhynchus mykiss_). Environ Toxicol Chem 22(6): 1272-1280.
-
Segner H, Caroll K, Fenske M, Janssen CR, Maack G, et al. (2003) Identification of endocrine–disrupting effects in aquatic vertebrates and invertebrates: report from the European IDEA project. Ecotoxicol Environ Saf 54(3): 302-314.
-
Lange A, Paull GC, Coe TS, Katsu Y, Urushitani H, et al. (2009) Sexual reprogramming and estrogenic sensitization in wild fish exposed to ethinylestradiol. Environ Sci Technol 43(4): 1219-1225.
-
Parrott JL, Blunt BR (2005) Life–cycle exposure of fathead minnows (_Pimephales promelas_) to an ethinylestradiol concentration below 1 ng/L reduces egg fertilization success and demasculinizes males. Environ Toxicol 20(2): 131-141.
-
Pelissero C, Flouriot G, Foucher JL, Bennetau B, Dunogu`es J, et al. (1993) Vitellogenin synthesis in cultured hepatocytes; an in vitro test for the estrogenic potency of chemicals. Journal of Steroid Biochemistry Molecular Biology 44(3): 263-272.
-
Heppell SA, Denslow ND, Folmar LC, Sullivan CV (1995) Universal assay of vitellogenin as a biomarker for environmental estrogens. Environmental Health Perspectives 103(S7): 9-15.
-
Palmer BD, Huth LK, Pieto DL, Selcer KW (1998) Vitellogenin as a biomarker for xenobiotic estrogens in an amphibian model system. Environmental Toxicology and chemistry 17(1): 30-36.
-
Bieberstein U, Berbner T, Islinger M, Braunbeck T (1999) Immunohistochemical localization of vitellogenin in rainbow trout (_Oncorhynchus mykiss_) hepatocytes using immunofluorescence. The Science of the Total Environment 233(1/3): 67-75.
-
Kelly C (2000) Analysis of steroids in environmental water samples using solid-phase extraction and ion- trap gas chromatography-mass spectrometry and gas chromatography- tandem mass spectrometry. J Chromatogr A 872(1-2): 309-314.
-
Schägger H (2006) Tricine-SDS-PAGE. Nat Protoc 1(1): 16-22.
-
van Leeuwen CJ, Vermeire TG (2007) Risk Assessment of Chemicals: An Introduction: Springer Science and Business Media.
-
Arukwe A, Goksoyr A (2003) Eggshell and egg yolk proteins in fish: hepatic proteins for the next generation: Oogenetic, population, and evolutionary implications of endocrine disruption. Comparative Hepatology 2(1): 4-25.
-
Panter GH, Hutchinson TH, Länge R, Lye CM, Sumpter JP, et al. (2002) Utility of a juvenile fathead minnow screening assay for detecting (anti-)estrogenic substances. Environ Toxicol Chem 21(2): 319-326.
-
Örn S, Holbech H, Madsen TH, Norrgren L, Peterse GI (2003) Gonad development and vitellogenin production in zebrafish (_Danio rerio_) exposed to ethinylestradiol and methyltestosterone. Aquat Toxicol 65(4): 397-411.
-
Vine E, Shears J, Van Aerle R, Tyler CR, Sumpter JP (2005) Endocrine (sexual) disruption is not a prominent feature in the pike _(Esox lucius)_, a top predator, living in English waters. Environmental Toxicology and Chemistry 24(6): 1436-1443.
-
Van der Oost R, Beyer J, Vermeulen NPE (2003) Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environmental Toxicology and Pharmacology 13(2): 57-149.
-
Tyler CR, Jobling S (2008) Roach, sex, and gender- bending chemicals: the feminization of wild fish in English Rivers. Bioscience 58(11): 1051-1059.
-
Chang CF, Lee MF, Chen GR (1994) Estradiol-17β associated with the sex reversal in protandrous black porgy, _Acanthopagrus schlegeli_. Journal of Experimental Zoology 268(1): 53-58.
-
Bjerregaard LB, Korsgaard B, Bjerregaard P (2006) Intersex in wild roach (_Rutilus rutilus_) from Danish sewage effluent-receiving streams. Ecotoxicology and Environmental Safety 64(3): 321-328.
-
Tetreault GR, Bennett CJ, Shires K, Knight B, Servos MR, et al. (2011) Intersex and reproductive impairment of wild fish exposed to multiple municipal wastewater discharges. Aquat Toxicol 104(3-4): 278-290.
-
Jobling SR, Williams R, Johnson A, Taylor A, Sorokin MG, et al. (2006) Predicted exposures to steroid estrogens in U.K. rivers correlate with widespread sexual disruption in wild fish populations. Environ Health Perspect 114(S1): 32-39.
-
Kavanagh RJ, Balch GCK, Kiparissis Y, Niimi AJ, Sherry J, et al. (2004) Endocrine disruption and altered gonadal development in White Perch (_Moroneamericana_) from the Lower Great Lakes Region. Environ Health Perspect 112(8): 898-902.
-
Hinck JE, Blazer VS, Schmitt CJ, Papoulias DM, Tillitt DE (2009) Widespread occurrence of intersex in black basses (_Micropterus spp._) from US rivers, 1995–2004. Aquatic Toxicology 95(1): 60-70.
-
Ackerman G, Amcoff P, Tjarnlund U, Fogelberg K, Torrissen O, et al. (2003) Paraquat and menadione exposure of rainbow trout (_Oncorhynchus mykiss_). Studies of effects on the penthose-phosphate shunt and thiamine levels in liver and kidney. Chemico- Biological Interactions 142(3): 269-283.
-
Figueiredo Fernandes A, Fontaínhas Fernandes A, Peixoto F, Rocha E, Reis Henriques MA (2006) Effects of gender and temperature on oxidative stress enzymes in Nile tilapia Oreochromis niloticus exposed to paraquat. Pesticide Biochemistry and Physiology 85(2): 97-103.
-
Barse AV, Chakrabarti T, Ghosh TK, Pal AK, Jadhao SB (2006) One-tenth dose of LC50 of 4-tertbutylphenol causes endocrine disruption and metabolic changes in _Cyprinus carpio_. Pesticide Biochemistry and Physiology 86 (3): 172-179.
-
Hecker M, Giesy JP, Jones PD, Jooste AM, Carr JA, et al. (2004) Plasma sex steroid concentrations and gonadal aromatase activities in African clawed frogs (_Xenopus_ _laevis_) from the corn- growing region of South Africa. _En_vironmental Toxicology and Chemistry 23(8): 1996- 2007.
-
Clouzot L, Marrot B, Doumenq P, Roche N (2008) 17α -ethinylestradiol: An endocrine disrupter of great concern. Analytical methods and removal processes applied to water purification. A review. Environmental Progress 27(3): 383-396.
-
Zohar Y, Abraham M, Gordin H (1978) The gonadal cycle of the captivity–reared hermaphrodyte teleost Sparus aurata (L.) during the first two years of life. Ann Biol Anim Bioch Biophys 18(4): 877-882.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells