Are Bases Alkylated by Branched Alkylating Agents Removed by Nucleotide Excision Repair or Base Excision Repair?
We have shown that comet-positive responses by adducts that are repaired by nucleotide excision repair (NER) and base excision repair (BER) are reduced by α-amanitin (AMN) and enhanced by 2’, 3’-dideoxythymidine (ddT), respectively. Based on this finding, bases alkylated by n-alkyl groups with <5 carbons and those with >4 carbons were discussed to be removed by BER and NER, respectively. To study whether bases alkylated by branched alkyl groups can be removed by NER or BER, 4 alkyl methanesulfonates that have a branched methyl group with different branching positions [iso-propyl methanesulfonates (iPMS), 1-methyl-pentyl methanesulfonate (1MPeMS), 2-methyl-pentyl methanesulfonate (2MPeMS), and 3-methyl-pentyl methanesulfonate (3MPeMS)] were synthesized. We studied the reduction of comet-positive responses by AMN and their enhancement by ddT for those alkyl methanesulfonates with a branched methyl group.
Introduction
Base adducts are classified into bulky adducts and small adducts in relation to their removal processes; the former including pyrimidine dimers induced by UV causes disturbances to the helical DNA structure and are removed by the nucleotide excision repair (NER) and the latter causes relatively minor disturbances to the helical DNA structure and are removed by the base excision repair (BER) [1, 2, 3, 4]. We have established a method to find out whether NER or BER acts to remove damaged bases [5]. In this method, whether NER or BER is working can be distinguished by the enhancement or reduction of comet-positive responses; a comet-positive response upon exposure to mutagens that produce damaged bases removed by BER is enhanced by 2’,3’-dideoxythymidine (ddT) and a comet-positive response upon exposure to mutagens that produce damaged bases removed by NER is reduced by α-amanitin (AMN). By using this method, we have shown that bases alkylated by n-alkyl groups with ≦5 carbons are removed by BER and those with ≧4 are removed by NER [5]. However, it has not been studied whether bases alkylated by alkyl groups with side chains can be removed by NER or BER.
In this study, to determine whether bases alkylated by alkyl groups with side chains can be removed by NER or BER, we synthesized one ethyl methanesulfonate with a branched mehyl group i.e. iso-propyl methanesulfonates (iPMS), and three pentyl methanesulfonates that have a branched methyl group with different branching positions: i.e. 1-methyl-pentyl methanesulfonate (1MPeMS), 2-methyl- pentyl methanesulfonate (2MPeMS), and 3-methyl- pentyl methanesulfonate (3MPeMS). To query whether bases alkylated by these are removed by NER or BER, the enhancing effect of ddT and reducing effect of AMN on the comet responses for them were studied. Materials and Methods
Chemicals
Methanesulfonate esters were prepared as previously described [6], Briefly, approximately 0.1 mol of the methyl pentyl alcohol [1-methyl-pentyl alcohol, 2-methyl-pentyl alcohol, 3-methyl-pentyl alcohol, or iso-propyl alcohol] in methylene chloride solution containing a 20% molar excess of pyridine at 0°C was added to methanesulfonyl chloride over a period of 30 min, and then stirred for an additional 120 min. The reaction mixture was first extracted with ice water, followed by cold 10% hydrochloric acid, saturated sodium hydrogen carbonate, and distilled water, and the methylene chloride solution was dried over anhydrous sodium sulfate and the oily ester was distilled under reduced pressure to afford 50- 60% yield of the methanesulfonate ester as a colorless liquid. The structures of alkyl methanesulfonates with a branched methyl group were shown in Figure 1. These were dissolved in dimethyl sulfoxide (DMSO, Wako Pure Chemical Industries, Ltd.). The DNA repair inhibitors hydroxyurea (HU), cytosine- 1-β-D-arabinofuranoside (araC), and 2’,3’dideoxythymidine (ddT), purchased from Wako Pure Chemical Industries, Ltd., were dissolved in DMSO. Regular (GP-42) and low-melting- point (LGT) agarose were obtained from Nacalai Tesque, Inc., and diluted to 1% in physiological saline.
TK6 human lymphoblastoid cells were maintained using RPMI 1640 medium (Nissui Pharmaceutical Co., Ltd.) supplemented with 10% horse serum (SAFC Biosciences) and 200 µg/mL sodium pyruvate.
Cell Treatment
Cells were exposed to alkyl methanesulfonates with and without AMN, ddT, or a combination of araC and HU (araC/ HU) as described in a previous study [5]. Briefly, cells at 2.5×106 cells/6-cm dish were treated with AMN (0.1μg/mL) for 5 h followed by the exposure to AMSs. Cells were exposed to AMSs with and without ddT (10 µM) or a combination of 1.8 mM araC and 10 mM HU (araC/HU) for 2h. AMN, ddT, and araC/HU were maintained in the medium until the end of the culture. Exposed cells were sampled immediately after chemical treatment and the percentage of viable cells was measured by the trypan blue exclusion test. Relative survivals (survivals of treated cells compared with that of an untreated control cells) were obtained.
Comet Preparation
The comet slides were prepared at the dose range at which relative survival levels were >70%. Treated cells were suspended in 1% agarose-LGT at 2.5 × 106 cells/75 µL, and 75 µL of cell suspension was immediately deposited on a fully frosted slide (Matsunami Glass Ind., Ltd., Osaka, Japan), which was coated with 1% agarose GP-42 and then covered with another slide glass. The slides were placed so as to allow the agarose to gel. The samples on the slides were then immediately exposed to a lysing solution (pH 10) of 2.5 M NaCl, 100 mM EDTA disodium (Na2EDTA), 10 mM Trizma, 1% sarkosyl, 10% DMSO, and 1% Triton X-100, and left at
4°C for 1 h. The slides were then placed on a horizontal gel electrophoresis platform and covered with a pH>13 alkaline solutions composed of 300 mM NaOH and 1 mM Na2EDTA. The slides were left in the solution at 0°C for 20 min to allow unwinding of nuclear DNA and the expression of alkali-labile sites to occur. The slides were subjected to electrophoresis at 0°C for 20 min and were rinsed with 400 mM Trizma (pH 7.5) to neutralize the excess alkalinity. The power supply was set at 1 V/cm and 250 mA. Each slide was stained with 50µL of 20µg/mL ethidium bromide (Wako Pure Chemical Industries, Ltd.) and covered with a coverslip. Fifty cells per dose were examined and photographed (black and white ASA 400 Fuji Film) at 200× magnification using a fluorescence microscope (Olympus) equipped with an excitation filter of 515–560 nm and a barrier filter of 590nm. The whole length of the comet and the diameter of the comet head were measured and tail length was obtained as the difference between the comet length and the head diameter. The assays were performed independently 3 times to ascertain the reproducibility of the results. After the reproducibility of the results was ascertained, mean values were calculated from the data obtained in the 3 independent trials. The effect of mutagen treatment on tail length was analyzed using ANOVA and Dunnett’s test. The mean values were compared between the treated slides and the control slides by Dunnett’s test after being tested by 1-way ANOVA. The mean values were compared between the inhibitor-treated slides and the untreated control slides at each mutagen concentration by Student’s t test. P values lower than 0.05 were considered statistically significant in all analyses. Results In the presence and absence of repair inhibitors, relative survival levels were >70% at the studied dose range of mutagens in TK6 cells (Table 1), from which the observed comet-positive response was not considered to be due to cytotoxicity. The effects of ddT, araC/HU, and AMN on the comet responses for AMSs in TK6 cells are shown in Figures 2A-2D. All studied AMSs showed positive comet responses in TK6 cells; without ddT, araC/HU, and AMN, iPMS at ≧62.5 µg/mL, methyl-PeMS at ≧313 µg/mL increased tail length significantly. Tail length with araC/HU was significantly higher than that without it for methyl PeMS and iPMS at studied concentration. Tail length with ddT was significantly greater than that without it for 2MPeMS and 3MPeMS at ≧1250 µg/mL, but no significant differences were observed between tail length with ddT and without it for 1MPeMS and iPMS. Significant decreases in tail length by AMN were observed for 1MPeMS, 2MPeMS, and iPMS but not for 3MPeMS; tail length was significantly lower with araC/HU plus AMN than with araC/HU for 1MPeMS and2MPeMS at ≧313 µg/mL and iPMS at ≧125 µg/mL, but tail length was almost the same with araC/HU and araC/HU plus AMN for 3MPeMS.
| AMS | Inhibitors | Relative survival (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dose of branched alkyl methane sulfonates (μg/mL) | ||||||||||||
| araC/HU | AMN | ddT | 0 | 62.5 | 125 | 250 | 313 | 500 | 625 | 1250 | 2500 | |
| iPMS | - | - | - | 100 | 93.1 | 88.8 | 90.2 | - | 95.4 | - | - | - |
| - | - | + | 98.7 | 92.2 | 98.3 | 105.4 | - | 103.1 | - | - | - | |
| + | - | - | 99.1 | 90.1 | 89.8 | 92.4 | - | 98.3 | - | - | - | |
| + | + | - | 95.4 | 93.1 | 98.3 | 96.2 | - | 105.7 | - | - | - | |
| 1MPeMS | - | - | - | 100 | - | - | - | 96.7 | - | 98.1 | 95.2 | 88.9 |
| - | - | + | 95.2 | - | - | - | 99.7 | - | 103.3 | 87.1 | 90 | |
| + | - | - | 105.3 | - | - | - | 83.3 | - | 111.2 | 91 | 95.1 | |
| + | + | - | 98.8 | - | - | - | 91.2 | - | 108 | 110.3 | 91 | |
| 2MPeMS | - | - | - | 100 | - | - | - | 77.7 | - | 103.9 | 108.6 | 106.1 |
| - | - | + | 103.1 | - | - | - | 90 | - | 98.9 | 100.9 | 92.1 | |
| + | - | - | 90 | - | - | - | 88.8 | - | 95.5 | 104.8 | 94.3 | |
| + | + | - | 101 | - | - | - | 95.2 | - | 109.6 | 88.6 | 91.2 | |
| 3MPeMS | - | - | - | 100 | - | - | - | 90.1 | - | 92.7 | 108.2 | 99.5 |
| - | - | + | 105.2 | - | - | - | 89.8 | - | 90.1 | 96.3 | 107.2 | |
| + | - | - | 88.2 | - | - | - | 107.3 | - | 93.4 | 97.6 | 96.7 | |
| + | + | - | 97.1 | - | - | - | 108.3 | - | 88.2 | 104.6 | 90 |
Table 1: Cytotoxicity of AMS in the presence and absence of DNA repair inhibitors. TK6 cells were exposed to AMSs in the presence
Table 1: Cytotoxicity of AMS in the presence and absence of DNA repair inhibitors. TK6 cells were exposed to AMSs in the presence and absence of each inhibitor. Exposed cells were sampled immediately after chemical treatment and the percentage of viable cells was measured by the trypan blue exclusion test and relative survival (%, survival compared with that of an untreated control) was obtained. Mean of three trials was shown.
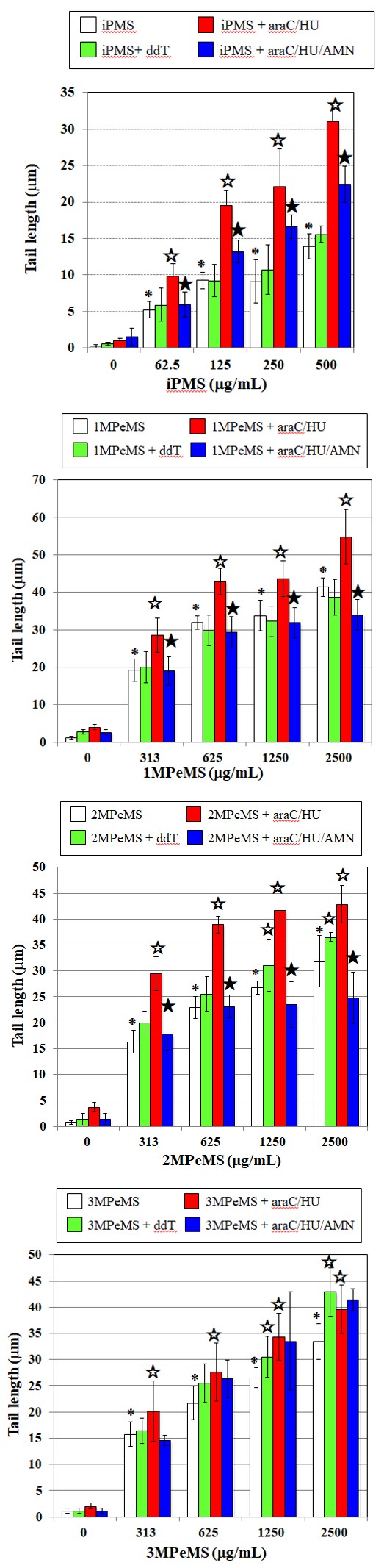
Figure 2A: iPMS Figure 2B: 1MPeMS.
Figure 2C: 2MPeMS. Figure 2D: 3MPeMS. *Significantly higher than untreated control: p<0.05 ☆Significantly higher than repair inhibitor-untreated control at each AMS concentration: p<0.05. ★Significantly lower than araC/HU at each AMS dose: p<0.05. Figure 2: Effects of AMN on comet responses by 4 kinds of AMSs with a branched methyl group in TK6 cells. Comet slides were prepared after culture for 2 h exposure to AMS for 2 h with and without DNA repair inhibitors. The error bars indicate standard deviation of the mean of three independent trials.
Discussion
The enhancement of comet-positive responses by ddT is due to its inhibition of DNA polymerase β depending re- synthesis in BER [7, 8, 9]. The suppression of comet-positive responses by AMN is due to its inhibition of RNA polymerase depending TCR sub-pathway in NER [10, 11, 12]. In this study, the effects of araC/HU, ddT, and AMN on the comet-positive responses upon exposure to 4 kinds of AMSs with a branched methyl group were studied. We have shown the method to know whether NER or BER acts to remove base adducts, in which ddT enhances a comet-positive response upon exposure to mutagens that producing damaged bases removed by BER and AMN can reduce a comet-positive response upon exposure to mutagens producing damaged bases removed
by NER [5]. The enhancement of comet-positive responses by ddT was observed for pentyl methanesulfonates with a methyl group branching at 2- and 3-position (2 MPeMS and 3MPeMS), suggesting that BER takes part in the excision of bases alkylated by 2MPeMS and 3MPeMS. AMN reduced comet-positive responses for pentyl methanesulfonates with a methyl group branching at 1- and 2-position (1MPeMS and 2MPeMS) and ethyl methanesulfonates with a methyl group branching at 1-position (iPMS), suggesting that NER works to remove bases alkylated by 2MPeMS, 1MPeMS, and iPMS. Considering that NER acts to remove bases alkylated by pentyl groups with a methyl group branching at 1-position (1MPeMS) and 2-position (2MPeMS) and ethyl groups having a methyl group branching at 1-position i.e. iso-propyl groups and that BER acts to remove bases alkylated by n-pentyl
group, ethyl groups [5] and pentyl groups with a methyl group with branching 3-position (3MPeMS), it could be considered that bases alkylated by these alkyl groups are not recognized as bulky bases. Therefore, the existence of a methyl group branching at 1- and 2-position could be considered to be the reason why alkylated bases are recognized as bulky bases and removed by NER.
Both NER and BER act to remove bases alkylated by pentyl groups with a methyl group branching at 2-position. Both NER and BER were shown to act to remove n-pentylated bases [5] and BER was not considered to act to remove bases alkylated by pentyl groups with a methyl group branched at 1-positions. Therefore, the possibility is shown that bases alkylated by pentyl groups with a branched methyl group whose branching position is closer to 1-position are not repaired by BER. Considering that the combination of araC and HU inhibits DNA re-synthesis without polymerase specificity [13, 14, 15, 16], it is reasonable that comet positive responses by studied 4 AMSs were enhanced by araC/HU regardless of whether alkylated bases are repaired by NER or BER.
Conclusion
Bases alkylated by methyl-alkyl groups of which branching position is closer to 1-position are shown to be repaired by NER. Therefore, the existence of a methyl group branching at closer to 1-position is considered to be the reason why alkylated bases are recognized as bulky bases and removed by NER.
Disclaimer
The products used for this research are commonly and predominantly used products in our area of research and country. There is absolutely no conflict of interest between the authors and producers of the products because we do not intend to use these products as an avenue for any litigation but for the advancement of knowledge. Also, the research was not funded by the producing company rather it was funded by personal efforts of the authors.
Acknowledgements
This research was conducted in Material and Biological Engineering Course, National Institute of Technology, Hachinohe College as part of a graduation research project by Kotaro Sasaki and Runa Kobayashi under the research guidance of course and staffs based on allocation of school educational expenses. The authors acknowledge the Material and Biological Engineering Course, National Institute of Technology, Hachinohe College.
Ethical Approval
It is not applicable.
Competing Interests
Authors have declared that no competing interests exist.
References
-
Budd ME, Campbell JL (1997) The roles of the eukaryotic DNA polymerases in DNA repair synthesis. Mutat Res 384: 157-167.
-
Ma L, Hoeijmakers JH, van der Eb AJ (1995) Mammalian nucleotide excision repair. Biochem Biophys Acta 1242(2): 137-163.
-
Sancar A (1995) Excision repair in mammalian cells. J Biol Chem 270(27): 15915-15918.
-
Mohrenweiser HW, Jones IM (1998) Variation in DNA repair is a factor in cancer susceptibility: a paradigm for the promises and perils of individual and population risk estimation?. Mutat Res 400(1-2): 15-24.
-
Odajima C, Nakamura T, Nakamura M, Miura M, Yamazaki K, et al. (2014) Role of nucleotide excision repair or base excision repair in movement of various n-alkylated Bases, investigated by the comet Assay. Genes Environ 36(1): 10-16.
-
Muller RK, Joos R, Felix D, Schreiber J, Wintner C et al. (1988) Preparation of N-aminoaziridine: trans-1-amino- 2,3-diphenylaziridine, 1-amino-2-phenylaziridium acetate. Organic Synthesis Coll 6: 56.
-
Dresler SL, Kimbro KS (1987) 2’,3’-Dideoxythymidine 5’-triphosphate inhibition of DNA replication and ultraviolet-induced DNA repair synthesis in human cells: evidence for involvement of DNA polymerase delta. Biochemistry 26(10): 2664-2668.
-
Budd ME, Campbell JL (1997) The roles of the eukaryotic DNA polymerases in DNA repair synthesis. Mutat Res 384(3): 157-167.
-
Odajima C, Nakamura T, Miura M, Yamazaki K, Honda G et al. (2013) Can an inhibitor of DNA polymerase β enhance the formation of comet tail?. Genes Environ 35(2): 46-52.
-
Zanotti G, Petersen G, Wieland T (1992) Structure- toxicity relationships in the amatoxin series. Structural variations of side chain 3 and inhibition of RNA polymerase II. Int J Pept Protein Res 40(6): 551-558.
-
Christians FC, Hanawalt PC (1992) Inhibition of transcription and strand-specific DNA repair by alpha- amanitin in Chinese hamster ovary cells. Mutat Res 274(2): 93-101.
-
Nguyen VT, Giannoni F, Dubois MF, Seo SJ, Vigneron M, et al. (1996) In vivo degradation of RNA polymerase II largest subunit triggered by α-amanitin. Nucleic Acids Res 24(15): 2924-2929.
-
Mayor PP, Egan EM, Herrick DJ, Kufe DW (1982) Effect of Ara-C incorporation on deoxyribonucleic acid synthesis in cells. Biochem Pharmacol 31(18): 2937-2940
-
Rojas E, López MC, Valverde M (1999) Single cell gel electrophoresis assay: methodology and applications. J Chromatogr B Biomed Sci Appl 722(1-2): 225-254.
-
Collins AR (2004) The comet assay for DNA damage and repair: principles, applications, and limitations. Molec Biotechnol 26(3): 249-261.
-
Collins ARS, Squires S, Johnson RT (1982) Inhibitors of repair DNA synthesis. Nucleic Acid Res 10(4): 1203- 1213.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells