Acetylcholinesterase Inhibition of Marine Fish (Rastrelliger kanagurta) by Organophosphorus Pesticides as Biomarker of Neurotoxicants
This paper evaluated the inhibitory effects of various phosphorothioates (Ethyl parathion and Chlorpyriphos) and phosphates (DDVP, Monocrotophos and Phosphamidon) on acetylcholinesterase (AChE) of marine Mackerel fish (Rastrelliger kanagurta) from the Goa coast. The characteristics of the AChE enzymes isolated from different tissues of the fishes for their affinity towards the substrate were determined by the Michaelis Menten constants. The pseudocholinesterase activity was also measured in the muscle tissues in terms of Butyryl-cholinesterase (BChE) activity of the fish. The AChE activity was found to be predominant in the Brain, gills, liver and muscle tissues. However, no significant BChE activity was observed in the muscle tissue. Among the five organophosphorus pesticides, DDVP was found to be most toxic with least I50 values (0.99±0.04nM) followed by Chlorpyriphos (I50, 24.59±2.86 nM), Ethyl Parathion (I50, 183.21±11.68 nM), Monocrotophos (I50, 13536.93±1292.94 nM) and Phosphamidon (I50, 21433.1±2651.96 nM) in the decreasing order. The I50 values of all other pesticides decreased considerably with respect to DDVP. The relative neurotoxicity of the pesticides decreased by an order of magnitude in the case of Chlorpyriphos followed by Ethyl Parathion by an order of magnitude whereas in the case of Monocrotophos and Phosphamidon the toxicity decreased significantly by an order of magnitude. The variation in the muscle AChE inhibition by the organophosphorus pesticides can be attributed to the sensitivity of the enzyme for each of the pesticides as observed by the bimolecular inhibition constants (KII). The muscle AChE inhibition of Rastrelliger kanagurta can be used as biomarker for bio- monitoring of neurotoxic contaminants in the marine environment.
Introduction
Acetylcholinesterase is the enzyme that is present within the synaptic cleft plays significant role in the neurotransmission [1]. Acetylcholinesterase (AChE) is widely distributed among the vertebrates and the invertebrates [2]. It hydrolyses acetylcholine to choline and acetic acid and thus initiating the passage for conduction of nerve impulse. AChE has, in fact, got the major role in Brain and muscle tissues of the animals [3].
Organophosphorus and carbamate pesticides are well known for their anti- cholinesterase activities [2, 4, 5, 6]. The residues of these pesticides are being transported into the coastal ecosystem via run off from the pesticide contaminated agricultural field, riverine input, domestic wastes disposal, municipal drainage and the industrial effluents from the surrounding pesticide manufacturing industries [7, 8, 9]. The marine organisms may thus be exposed to neurotoxic contaminants the concentrations of which are biomagnified through the food chain leading to serious damage to human beings consuming sea- foods. Thus, it is indeed an urgent need of the hour to assess the impact of these contaminants in the marine ecosystem. The acetylcholinesterase activity has been used worldwide as a bio-monitoring tool for assessment of the prevalence of neurotoxic contaminants in the environment [4, 10, 11]. The inhibitory effects by these kinds of pesticides on acetylcholinesterase activity of various species of aquatic organisms such as fishes [12, 13, 14, 15, 16, 17, 18], Valbonesi P, et al. [19]; Matozzo V, et al. [20]; Lau PS, et al. [7]; Binelli A, et al. [21]; Doran WJ, et al. [22], Cajaravalle MP, et al. [1]; Lionetto MG, et al. [23]; Magni P, et al. [24]; Escartin E, et al. [25]; Pfeifer S, et al. [26], gastropods Fite C, et al. [27]; Gaitonde D, et al. [6] copepod Forget J, et al. [28] have been very well documented. Many in-vitro and in-vivo studies were carried out on various marine species using AChE as a biomarker Sarkar A, et al. [5]; Binelli A, et al. [21]; Lionetto MG, et al. [23]; Matozzo V, et al. [20]; Escartin E, et al. [25]. Lionetto MG, et al. [23] tudied the acetylcholinesterase (AChE) activities in Mytilus galloprovincialis and a benthic teleost fish (Mullus barbatus) from the coastal marine environment of Salento Peninsula (Italy) in order to detect the possible exposure/effect induced by chemical pollutants in native marine organisms. An extensive study was earlier carried out on acetylcholinesterase activity in marine gastropod (Cronia contracta) all along the Goa coast to assess the impact of neurotoxic contaminants on the marine organisms [6]. Lau PS, et al. [7] assessed the seasonal variations and potential influence of the riverine discharge from the Pearl River on acetylcholinesterase (AChE) activities in the green mussel (Perna viridis) as a biomarker of exposure to contaminants in Hong Kong waters. Sturm A, et al. [3] monitored neurotoxic contamination in the marine teleost fish (Limanda, Platichthys flesus and Serranus cabrilla) from the Mediterranean Sea using cholinesterase activities.
As the marine coastal environment is exposed to various types of neurotoxic contaminants because of agricultural runoff, indiscriminate disposal of domestic wastes and industrial effluents, the present study emphasises on the evaluation of the effects of neurotoxic contaminants on marine organisms along the Goa coast using muscle AChE activity from the Mackerel fish (Rastrelliger Kanagurta). Mackerel fish was chosen as the sentinel marine species for the present study since it is the most abundantly available edible fish all along the west coast of India and thereby can act as the direct source of the neurotoxic contamination of the human beings leading to serious damage to their health. The in-vitro AChE inhibition study using various organophosphorus pesticides provides an insight into the severity of their presence in the coastal environment. The inactivation of the AChE activity or its disorder due to exposure to anti-acetylcholinesterase active contaminants in the marine environment can lead to several neural diseases such as Alzheimer, Parkinson, and Paralysis etc. Berson A, et al. [29]; Landwehrmeyer B, et al. [30]; Carson KA, et al. [31]. The toxicity of organophosphorus pesticides is measured in terms of their inhibitory effect on acetylcholinesterase activity. The relative toxicity of these pesticides may vary in accordance with their structural significance. In order to assess the inhibitory potencies of various organophosphorus pesticides on acetylcholinesterase activity an extensive in- vitro study was carried out with muscle AChE of Rastrelliger kanagurta.
Materials and Method Sampling
A large number of samples of marine Mackerel fishes (Rastrelliger kanagurta) were collected from Betul along the Goa coast (Lat: 15o 17’33.18”, Long: 73o 54’ 29.232’). Betul was chosen for collection of the fish samples because of its pristine environment. Immediately after collection, the fish samples were brought into the laboratory for isolation of the enzymes from different tissues. About 10 fish samples were used for isolation of AChE enzymes from different tissues.
Physical Characteristics of the Mackerel Fish
The mackerel fishes are in fact, marine pelagic and mostly available in the coastal region. The average length, breath and the body weight of the fishes were in the range 22 ± 2.5cm, 5.3 ± 0.8cm and 105.24 ± 10.14 g respectively. They have dorsal spines (total): 8-11; Dorsal soft rays (total): 12-12; Anal spines: 0; Anal soft rays: 12; fins 2; finlets: dorsal 5-5 and ventral 5-5; anal fin: 1. Head of the fish were found to be longer than body depth. A black spot can be seen on body near lower margin of pectoral fin. Swim bladder is present.
Adults occur in coastal bays usually in some turbid plankton- rich waters. They feed on phytoplankton (diatoms) and small zooplankton (cladocerans, ostracods, larval polychaetes, etc.) Adult individuals feed on macroplankton such as larval shrimps and fish. Their eggs and larvae are pelagic.
Isolation of AChE Enzyme
The fish samples were preserved in the laboratory in ice. The tissues were removed from the fishes at low temperature using ice flakes. The tissues removed from the fishes were mixed together to make composite samples and subdivided into three parts. The quantity of tissues used for isolation of AChE enzymes from the composite samples were in the following ranges, brain, 1.04-1.15g, muscle, 7.60-10.85g, gills, 3.64-3.66g and liver, 6.42-6.60g.
The acetylcholinesterase enzymes were isolated from the tissues by homogenization with 0.01M phosphate buffer (pH -7.4±0.1) spiked with triton-X-100 (0.2%) and 0.25M sucrose (1:1) solution using an Ultra Turrax homogenizer (T-25) at a speed (16000 to 19000 rpm) [6, 10]. The homogenates were centrifuged by refrigerated centrifuge (Eltek) at 20,000G for about 35 minutes at 4℃. The protein concentration of each of the sample extract was determined according to Lowry OH, et al. [32] using bovine serum albumin as the standard.
Measurement of the AChE Activity
The acetylcholinesterase activity was determined following the modified Δ-pH metric method [6, 10], using phosphate buffer (pH-8.0), acetylcholine bromide as the substrate and bromothymol blue as the indicator. The changes in pH (Δ-pH) were measured at an interval of 10 minutes over a period of one hour of incubation corresponding to the amount of acetic acid liberated due to interaction with the acetylcholinesterase enzyme. The AChE activity is expressed in terms of nmoles of acetic acid liberated per mg of protein per minute. The in-vitro inhibition of AChE was carried out with different pesticides after incubation with the enzyme for about 15 minutes before the addition of the substrate into the reaction medium. The percentage of inhibition was calculated with respect to the control (without the inhibitor). BChE was determined using butyryl-cholineiodide as the substrate after suppressing the AChE activity with an AChE inhibitor, BW284C51 (1, 5-bis (4 allyl-dimethyl ammonium phenyl pentan -3 one dibromide).
The bimolecular inhibition constant (KII) for each pesticide were calculated using the equation,
2.3log(A / A ) / I/ t KII o i =
Where, I is a final molar concentration of inhibitor, Ao and Ai
are the enzyme activities without and with inhibitor and t is the time of inhibition pre-incubation (15 minutes) before the substrate addition O’Brien RD, et al. [33].
The I50 values for inhibition of AChE activity by different pesticides were calculated by the regression equation shown in the figures.
Result and Discussion
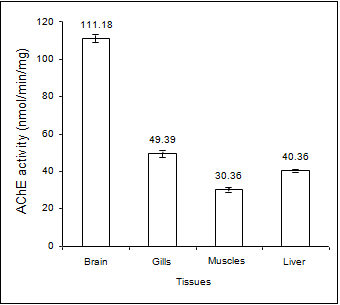
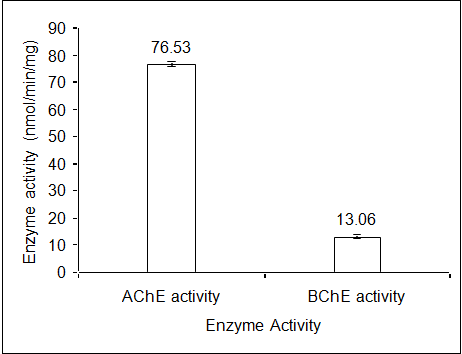
The variation in AChE activities, expressed in terms of nmol/min/mg in different tissues of the edible marine Mackerel fishes showed highest activity in brain tissue (111.18± 3.01) followed by gills (49.39 ± 2.64), liver (40.36 ± 0.91) and muscles (30.36 ± 1.87) (Figure 1). The characteristics of the AChE enzyme in different tissues were determined by the Michaelis Menten constants (Table 1) using the Line weaver-Burke Plot (Figures 2a & 2b). The pseudocholinesteras (i.e. BChE) activity in the muscle tissue of the fish was found to be very low in comparison with the AChE activity (Figure 3). For measurement of AChE activity, a BChE inhibitor, (Iso-OMPA) was used to suppress the BChE activity. Interestingly, the AChE activity was found to be predominant in the muscle tissue even without using the BChE inhibitor, which could be due to low pseudocholinesterase activity in the species.
| Tissue | Km (mM) | Vmax(nmol of acid/min) |
|---|---|---|
| Liver | 0.587 | 217.39 |
| Gills | 0.412 | 7.225 |
| Muscles | 0.45 | 81.967 |
| Brain | 0.21 | 123.46 |
Table 1: Kinetic constants for AChE in different tissues of _R._ _kanagurta._
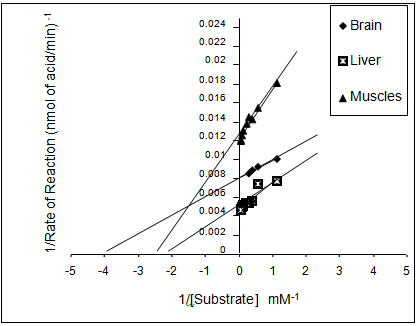
(a) (b) Figure 2: Line weaver Burke plot for AChE enzyme from different tissues a) brain, liver and muscle; b) gills of R. kanagurta.
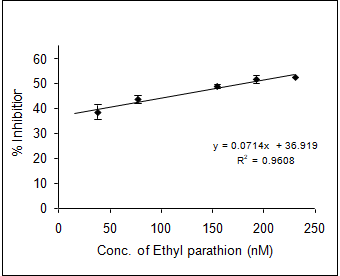
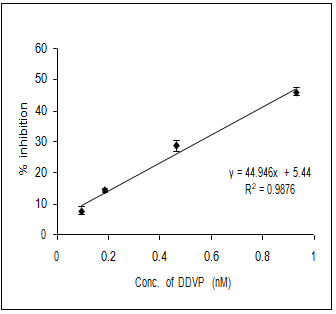
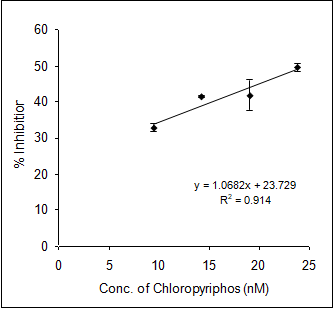
In order to evaluate the neurotoxic potential of various organophosphorus pesticides such as three enol phosphates (DDVP, Phosphamidon and Monocrotophos) and two thiophosphates (Ethyl parathion and Chlorpyriphos) in-vitro inhibition of AChE activity was carried out with muscle AChE of the marine fish from the Goa coast. Figures 4 & 5 clearly show the inhibitory potencies of these pesticides in terms of their I50 value (the concentration of the inhibitor at which 50% inhibition of the enzyme activity occurs).
It has been observed that DDVP shows the least I50 (0.99±0.04nM) followed by Chlorpyriphos (24.59±2.86 nM), Ethyl Parathion (183.21±11.68 nM), Monocrotophos (13536.93±1292.94 nM) and Phosphamidon (21433.1±2651.96 nM). In fact, the toxicity (expressed in terms of 1/I50) of the pesticide is inversely related to the I50 value.
(a) (b) (c)
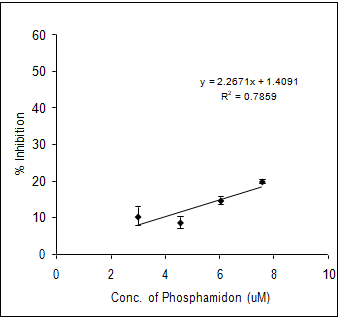
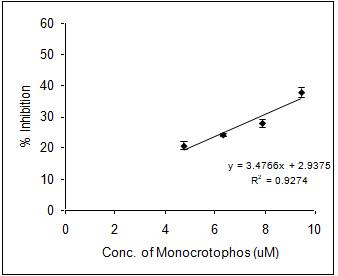
(d) (e) Figure 4: Inhibition of muscle AChE activity of R. kanagurta by different organophosphorus pesticides a) DDVP; b) Chlorpyriphos; c) Ethyl Parathion; d) Monocrotophos and e) Phosphamidon. All the data were in triplicates and highly significant p<0.0001.
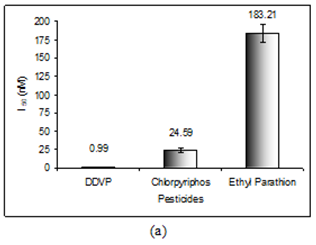
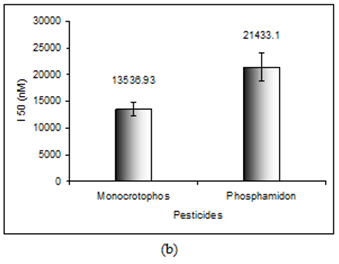
Thus, it is quite clear that lower the I50 value the higher the toxicity responses. The relative toxicity was calculated for each of the pesticides with respect to DDVP being the most toxic organophosphorus pesticide among the five (Table 2).
| Pesticide* | I (nM) 50 | Toxicity (1/I ) 50 | Relative Toxicity |
|---|---|---|---|
| (nM-1) | (1/I Pesticide/1/I DDVP) 50 50 | ||
| DDVP | 0.99±0.04 | 1.01 | 1 |
| Chlorpyriphos | 24.59±2.86 | 4.07 x 10-2 | 4.04 x 10-2 |
| Ethyl parathion | 183.21±11.68 | 5.46 x 10-3 | 5.43 x 10-3 |
| Monocrotophos | 13536.93±1292.94 | 7.39 x 10-5 | 7.34 x 10-5 |
| Phosphamidon | 21433.1±2651.96 | 4.67 x 10-5 | 4.64 x 10-5 |
Table 2: The toxicity of different pesticides in R. kanagurta. *All the data presented are means of triplicate and highly signifi
In this regard Chuiko GM, et al. [14] made a comparative study on the inhibition of brain and serum AChE and BChE of freshwater teleost species belonging to four families (Cyprinidae: common carp Cyprinus carpio, bream Abramis brama, blue bream A. ballerus, white bream Blicca bjoerkna, roach Rutilus rutilus, bleak Alburnus, ide Leuciscus idus; Percidae: perch Perca fluiatilis, pikeperch Stizostedion lucioperca; Esocidae: pike Esox lucius and Coregonidae: whitefish Coregonus albula) by DDVP in-vitro. Chuiko GM, et al. [14] reported that the brain AChE has the highest activity which is in accord with our investigation. It has been observed that the sensitivity of in-vitro cholinesterase inhibition by DDVP in the fresh water fishes depends in accordance with the variation in the species of fishes. However, the BChE sensitivity to in-vitro DDVP inhibition was found to be very insignificant among the different species of fishes. In contrast, Sturm A, et al. [3] reported higher sensitivity of BChE to organophosphates than AChE in marine teleost fish (Limanda, Platichthys flesus and Serranus cabrilla). This clearly substantiated the fact that the BChE activity was not necessarily be always low in all species of fishes rather it varies from species to species.
Moreover, Straus and Chambers (1995) studied the potency of in-vitro inhibition of AChE by the oxon derivatives of Chlorpyriphos and Parathion on channel catfish (Ictalurus punctatus). They reported that the I50 values corresponding to Chlorpyriphos and Parathion were in the range, 28-33nM and 446-578nM respectively which are very comparable to the present study.
Rehiayani A, et al. [34] examined the inhibition potency of some organophosphorus (Paraxon, Fenamiphos, Phorate- sulfoxide) and carbamate pesticides (such as, Aldicarb, Carbofuran and Oxamyl) on acetylcholinesterase in-vitro using selected plant-parasitic nematodes (Meloidogyne javaica, Heterodera avenae and Tylenchulus semipenetrans). The I50 values for each of these pesticides studied were in the range, paraxon, 7 x 10-7 M, Feminaphos, 2 x 10-6 M, Phorate- sulfoxide, 2 x 10-4 M, Carbofuran, 5 x 10-8 M, 7.5 x 10-7 M and 2 x 10-3 M respectively against T semipenetrans species. Whereas high inhibition potency was observed against the other species, M. javanica. Manildo MO, et al. [16] studied brain acetylcholinesterase as a biomarker using Brazilian fishes. The enzyme sensitivity to Methyl Paraxon showed that some of the species namely, Paralonchurus brasiliensis and Genidens genidens and Percophis brasiliensis were found to be very sensitive as indicated by their I50 values, 455 nM and 468 nM respectively. However, relatively less sensitivity was observed with respect to the species Merluccius hubbsi and Percophis brasiliensis (I50, 3339 and 3259 nM respectively). Thus, it is amply clear that the inhibition potency of the organophosphorus pesticides varies according to the species. The most striking point is that the toxic potential of the pesticides in comparison with DDVP decreased by an order of magnitude 2, 3, 5 and 5 for Chlorpyriphos (4.04 x 10-
2), Ethyl Parathion (5.43 x 10-3), Monocrotophos (7.34 x 10-
5) and Phosphamidon (4.64 x 10-5) respectively. The toxicity of the pesticides can be attributed to the variation in the sensitivities of the enzyme towards the pesticides expressed in terms of biochemical inhibition rate constant (KII).
The sensitivity of the enzyme towards DDVP was found to be maximum (614.720 ± 71.30) x105 molL-1 min-1). The variation in the KII value (Table 3) clearly substantiate the order of the sensitivity of the enzyme towards the inhibitors, which increases in the following order, Phosphamidon< Monocrotophos< Ethyl parathion< Chlorpyriphos <DDVP.
Moreover, the interaction between the inhibitor and the enzyme was further evaluated by determining the Michaelis Menten constant (Kmi) using Lineweaver- Burke Plot (Table 4). It provides a clear picture about the affinity of the enzyme for different organophosphorus pesticides. It may be noted that higher the Kmi value the lower the toxicity of the pesticide. The Kmi values for each of the pesticides substantiate the previous trend of their toxicity confirming DDVP as the most toxic pesticide followed by Chlorpyriphos, Ethyl parathion, Monocrotophos and Phosphamidon.
| $K_{\mathrm{II}} \times 10^{5}$ | |
|---|---|
| $(\text{Bimolecular inhibition Constant}) (\text{mol}^{-1}\text{Lmin}^{-1})$ | |
| DDVP | 614.720 ± 71.30 |
| Chlorpyriphos | 23.960 ± 4.040 |
| Ethyl Parathion | 9.290 ± 1.100 |
| Monocrotophos | 0.032 ± 0.003 |
| Phosphamidon | 0.018 ± 0.003 |
Table 3: Bimolecular inhibition rate constant (Kii) of different pesticides with respect to in-vitro muscle AChE inhibition. *All
| Pesticides | Kmi | V (nmol/min/mg) max |
|---|---|---|
| DDVP | 00.69 nM | 45.05 |
| Chlorpyriphos | 11.42 nM | 44.25 |
| Ethyl Parathion | 27.89 nM | 30.12 |
| Monocrotophos | 15.04 µM | 46.51 |
| Phosphamidon | 37.36 µM | 53.48 |
Table 4: Kinetic constants (Kmi) for the inhibition of AChE of _R. kanagurta_ by different pesticides.
Evaluation of the Toxicity in Terms of Structural Significance
Table 5 shows that among the five organophosphorus pesticides, three belongs to enol phosphate group (Monocrotophos, Phosphamidon, DDVP) whereas the other two pesticides belong to thio-phosphate group (Chlorpyriphos, Ethyl parathion). The most interesting point is that all the three enol phosphates contain vinyl group with some variation in substitution of the H-atoms by different functional group that contributes significantly towards the variation in their inhibitory potency.
In the case of Monocrotophos and Phosphamidon, one of the H-atom of the vinyl group is substituted by N-methyl amide and N, N- diethyl amide group with an additional chlorine substitution of the other H-atom respectively. Thus, not only the inductive effect of the amide group but also the steric factor owing to the bulky amide groups play significant roles towards the inhibition of AChE activity.
In the case of phosphorothioate pesticides (Ethyl parathion and Chlorpyriphos), the H-atom of the thio- phosphate is substituted by nitro-benzene and trichloro pyridine group respectively. The higher toxicity of Chlorpyriphos can be attributed to the Substitution of three chlorine atom on pyridine aromatic ring.
| No. | Organophosphorus Pesticides | Structure |
|---|---|---|
| 1 | Ethyl Parathion0,0-Diethyl-O(p-Nitrophenyl) phosphorothioate | C2H5O S P O NO2 |
| 2 | Monocrotophos0,0-diemethyl-O-(2-N-methylcarbamoyl-1-methylvinyl) phosphate | CH3O P O C=CH C NH CH3 |
| 3 | DDVP2,2-Dichlorovinyl dimethyl phosphate | CH3O P O CH=C Cl CH3 |
| 4 | Phosphamidon0,0-Dimethyl-O-(1-methyl-2-Chloro-2-diethylcarbamoylvinyl) phosphate | CH3O P O C=C C N C2H5 |
| 5 | Chlorpyriphos0,0-Diethyl 0-3,5,6-trichloro-2-pyridyl phosphorothioate | C2H5O S P O N Cl C2H5O |
Table 5: The structural characteristics of different Organophosphorus pesticides.
Conclusion
Among the five organophosphorus pesticides, DDVP was found to be the most toxic pesticide for its significant inhibitory potential followed by Chlorpyriphos, Ethyl Parathion, Monocrotophos and Phosphamidon. The in-vitro muscle AChE inhibition in the mackerel fish (Rastrelliger kanagurta) provides an early warning signal for the prevalence of neurotoxic contaminants in the environment. The AChE inhibition can be used as biomarker for environmental contamination.
Acknowledgement
The authors are thankful to the Director, CSIR-NIO for his keen interest and encouragement for the research work. They are thankful to the CSIR for the fellowship offered to one of the authors (D. Vashistha) for the research and development work.
References
-
Carajaville MP, Bebianno MJ, Blasco J, Porte C, Sarasquete C, et al. (2000) The use of biomarkers to assess the impact of pollution in coastal environment of the Iberian Peninsula: a practical approach. Sci Total Environ 247(2- 3): 295-311.
-
Bocquené G, Roig A, Fournier D (1997) Cholinesterases from the common oyster (Crassostrea gigas): Evidence for the presence of a soluble acetyl cholinesterase insensitive to organophosphate and carbamate inhibitors. FEBS Letters 407(3): 261-266.
-
Sturm A, da Silva de Assis HC, Hansen PD (1999) Cholinesterases of marine teleost fish: enzymological characterization and potential use in the monitoring of neurotoxic contamination. Mar Environ Res 47(4): 389- 398.
-
Sarkar A (2006) Marine Pollution and Ecotoxicology, Guest editorial Environment International 2(2): 145- 147.
-
Sarkar A, Ray D, Shrivastava AN, Subhodeep S (2006) Molecular Biomarkers: Their Significance and application in Marine Pollution Monitoring. Ecotoxicology 15(4): 333-340.
-
Gaitonde D, Sarkar A, Kaisary S, Silva C, Dias C, et al. (2006) Acetylcholinesterase activities in marine snail (Cronia contracta) as a biomarker of neurotoxic contaminants along the Goa coast, West coast of India. Ecotoxicology 15: 353-358.
-
Lau PS, Wong HL, Garrigues PH (2004) Seasonal variation in antioxidative responses and acetylcholinesterase activity in Perna viridis in eastern oceanic and western estuarine waters of Hong Kong. Continent Shelf Res 24(16): 1969 -1987.
-
Sarkar A, Nagarajan R, Singbal SYS, Chhapadkar S, Pal S (1997) Contamination of organ chlorine pesticides in sediments from the Arabian Sea along the west coast of India. Water Res 31(2): 195-200.
-
Sarkar, Everaarts (1998) Riverine input of chlorinated hydrocarbons in the coastal pollution. In: Ecology of Wetlands, River and Associated Habitats. The Pennsylvania Acad of Sci, pp: 400-422.
-
Sarkar A (1992) Evaluation of the toxicity of organic matter in marine sediments. Water Science and Technology 25(3): 255-257.
-
Wells PG, Depledge MH, Butler JN, Manock JJ, Knap AH (2001) Rapid Toxicity Assessment and Biomonitoring of Marine Contaminants -Exploiting the Potential of Rapid Biomarker Assays and Microscale Toxicity Tests. Mar Pollut Bull 42(10): 799-804.
-
Sen Gupta R, Sarkar A, Kureishy TW (1991) Bio- degradation and Anticholinesterase activity of methyl isocyanate in the aquatic environment of Bhopal. Water Research 25(2): 179-183.
-
Weiss CM (1958) the determination of cholinesterase in the brain tissue of three species of fresh water fish and its inactivation in-vivo. Ecology 39(2): 194-199.
-
Chuiko GM (2000) Comparative study of acetylcholinesterase and butyrylcholinesterase in brain and serum of several fresh water fish: specific activities and in-vitro inhibition by DDVP, an organophosphorus pesticide. Comparative biochemistry and Physiology C Toxicol Phormacol 127(3): 233-242.
-
Galgani F, Bocquené G, Cadiou Y (1992) Evidence of variation in cholinesterase activity in fish along a pollution gradient in the North Sea. Mar Ecol Prog Ser 91: 77-82.
-
Manildo MO, Filho S, Moacelio V, Bastos C, Vera LF, et al. (2007) Brain acetylcholinesterase as a marine pesticide biomarker using Brazilian fishes. Mar Environ Res 63(4): 303-312.
-
Straus David L, Chambers Janice E (1995) Inhibition of acetylcholinesterase and aliesterase of fingerling channel catfish by chlorpyrifos, parathion and S, S, S- tributyl phosphorotrithioate (DEF). Aquatic Toxicology 33(3-4): 311-324.
-
De la Torre FR, Ferrari L, Salibián A (2002) Freshwater pollution biomarker: response of brain acetylcholinesterase activity in two fish species. Comparative Biochemistry and Physiology C Toxicol Phormacol 131(3): 271-280.
-
Valbonesi P, Sartor G, Fabbri E (2003) Characterization of cholinesterase activity in three bivalves inhabiting the North Adriatic Sea and their possible use as sentinel organism for biosuveillance programmes. Sci Total Environ 312(1-3): 79-88.
-
Matozzo V, Tomei A, Marin MG (2005) Acetylcholinesterase as a biomarker of exposure to neurotoxic compounds in the clam Tapes philippinarum from the Lagoon of Venice. Mar Pollut Bull 50(12): 1686-1693.
-
Binelli A, Ricciardi F, Riva C, Provini A (2005) Screening of POP pollution by AChE and EROD activities in Zebra mussels from the Italian Great Lakes. Chemosphere 61(8): 1074-1082.
-
Doran WJ, Cope WG, Rada RG, Sandheinrich MB (2001) Acetylcholinesterase inhibition in the three ridge mussel (Amblema plicata) by chlorpyrifos: implications for biomonitoring. Ecotoxicol Environ Saf 49(1): 91-98.
-
Lionetto MG, Caricato R, Giordano ME, Pascariello MF, Marinosci L, et al. (2003) Integrated use of biomarkers (acetylcholinesterase and antioxidant enzymes activities) in Mytilus galloprovincialis and Mullus barbatus in an Italian coastal marine area. Mar Pollut Bull 46(3): 324-330.
-
Magni P, De Falco G, Falugi C, Franzoni M, Monteverde M, et al. (2006) Nontoxicity biomarkers and acetylcholinesterase activity in natural populations of Mytilus galloprovincialis along a pollution gradient in the Gulf of Oristano (Sardinia, western Mediterranean). Environ Pollut 142(1): 65-72.
-
Escartin E, Porte C (1997) the use of cholinesterase and carboxyl esterase activities from Mytilus galloprovincialis in pollution monitoring. Environ Toxicol Chem 16(10): 2090-2095.
-
Pfeifer S, Doris S, and Dippner JW (2005) Effect of temperature and salinity on acetylcholinesterase activity, a common pollution biomarker, in Mytilus sp. from the south-western Baltic Sea. J Exp Mar Biol and Ecol 320(1): 93-103.
-
Fite C, Malathi S (2003) Neural And Circulating Holinesterases of the Marine Mollusk Aplasia Californica. Moscow University Chemistry Bulletin 44(1): 62.
-
Forget J, Beliaeff B, Bocquené G (2003) Acetylcholinesterase activity in copepods (Tigriopus brevicornis) from the Vilaine River estuary, France, as a biomarker of neurotoxic contaminants. Aquatic Toxicology 62(3): 195-204.
-
Berson A, Knobloch M, Hanan M, Diamant S, Sharoni M, et al. (2008) Changes in readthrough acetyl cholinesterase expression modulates amyloid-beta pathology. Brain 131(1): 109-119.
-
Landwehrmeyer B, Probst A, Palacios JM, Mengod G (1993) Expression of acetylcholinesterase messenger RNA in human brain: an in situ hybridization study. Neuroscience 57(3): 615-634.
-
Carson KA, Geula C, Mesulam MM (1991) Electron microscopic localization of cholinesterase activity in Alzheimer brain tissue. Brain Res 540(1-2): 204-208.
-
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193(1): 265-275.
-
O’Brien RD (1960) Toxic phosphorus esters. Chemistry, metabolism and biological effects.
-
Rehiayani A, Suleiman M (2008) Acetyl cholinesterase in selected plat-parasitic nematodes: Inhibition, kinetic and comparative studies. Pesticide Biochemistry and Physiology 90(1): 19-25.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells