Induction of DNA Damage and Apoptosis Mediate Cytotoxic by Oxaliplatin In Human Breast Cancer MCF-7 Cell Lines in Vitro
Cancer is an extremely disease leading cause of human death worldwide, and breast cancer is the most common type of malignancy in women. It is a heterogenous and hormone-dependent disease. Oxaliplatin is a novel platinum derivative and is a potent chemotherapeutic agent. Therefore, oxaliplatin is an apoptotic effectual compound for treating cancer with cytotoxic side effects. To explore the underlying mechanism of action of oxaliplatin, we examined the induction of apoptosis on human breast cancer MCF-7 cell lines by the drug and introduced its possible mechanism of action. The oxidative-induced DNA damage was evaluated by 8-hydroxy-2-deoxyguanosine (8-OHdG) reduction. The cellular pathways involved in the apoptosis of P53 and BCL2 were also assessed by qPCR and Western blotting assays. The results showed that oxaliplatin exposure causes increased oxidative stress levels and activation of P53, and repression of BCL2 was also involved in these mechanistic pathways.
Introduction
Breast cancer is a prevalent type of cancer among women and a significant cause of global death [1]. In 2020, there were approximately 2.26 million breast cancer cases, making it the most frequently diagnosed cancer worldwide [2]. In cancer cells, growth and survival are closely related to a reduced level of genetically programmed cell death referred to as apoptosis [3, 4]. A chemotherapeutic drug, such as oxaliplatin, relies heavily on apoptosis to destroy cancer cells [5].
Oxaliplatin exerts its cytotoxic effect mainly by inducing DNA strand breakage and inhibiting DNA synthesis, eventually leading to apoptosis [6]. P53 is a tumour suppressor protein that controls apoptosis and cell cycle arrest after cytotoxic drugs and is one of the most commonly mutated genes in several types of cancer [7]. It has also been shown that oxaliplatin treatment upregulated p53 in colorectal cancer cell lines [8]. Oxaliplatin can induce p53-independent responses in various types of malignancy [9], and the study furthermore suggests the involvement of downregulation of BCL2 in oxaliplatin-mediated apoptosis [10].
Reactive oxygen species (ROS) are produced during normal cellular metabolic processes and after exposure to harmful factors, such as chemical carcinogens. Various studies in vivo and in vitro have provided supporting evidence between oxidative stress and increased tumour cell growth and cancer development [11]. More recent studies have indicated that altered production of ROS is also associated with cancer cells and regulators of important signalling pathways [12]. Chemotherapy drugs also can impact the coordinated regulation of disrupted cellular ROS balance.
Even though oxaliplatin is currently used to treat breast cancer, the true potential of the drug and its anticancer signalling mechanism remain unknown. As oxaliplatin can employ a variety of mechanisms as yet undefined to induce apoptosis in breast cancer cells, it might be helpful to investigate the exact mechanism by which it acts in breast cancer. In this study, we sought to determine whether oxaliplatin could induce apoptosis in MCF-7 cells of the human breast cancer cell line and explore the underlying mechanisms of apoptosis.
Materials and Methods
Cell Culture and Treatment
MCF-7 cell lines were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Gibco by Thermo Fisher Scientific) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and antibiotics (100 IU/ml penicillin and 100 mg/ml streptomycin) in 5% CO2 at 37°C. Oxaliplatin was added to the cultures at final concentrations of 0.2, 2, and 20 µM after 24 hours of incubation [11]. When the cells were approximately 80% confluent in T75 cm2 tissue culture flask, they were detached with 0.25% trypsin ethylenediaminetetraacetic acid (EDTA) solution and counted using hemocytometer, and then used in experiments.
Cell Viability
Cell Counting Kit-8 (CCK-8) was used to determine cell viability. The cells were seeded in 96-well plates at 5000 cells/well density and incubated at 37 °C in a humidified atmosphere with 5% CO2 for 24 h. The cells were cultured with 0.2, 2 and 20µM oxaliplatin. 10 μL of the CCK-8 reagent were added to each well and incubated for four h at 37°C. Cells cultured in a medium without the addition of oxaliplatin was considered as the negative control. Absorbance was measured at 450 nm using the Microplate reader MRX II (Dynex Technologies, Chantilly, USA). Three replicates were made for each measurement.
Detect 8-Hydroxy-2-Deoxyguanosine Level
The levels of 8-hydroxy-2-deoxyguanosine (8-OHdG) in the culture medium were measured by an 8-OHdG ELISA kit (ab201734, Abcam) following the manufacturer’s instructions. Absorbance values were measured on a microplate reader.
Total RNA Extraction and Reverse Transcription (RT)
Quantitative PCR for determining transcripts of P53 and BCl2 and GAPDH were performed using SYBR Green Supermix (Applied Biosystems) on step one plus a real-time PCR detection system (Applied Biosystems, Warrington, UK) according to the manufacturer’s protocol. Total RNA was extracted from cells using the TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). The total RNA was then synthesised to cDNA using iScript cDNA Synthesis Kit (Bio- Rad) following the manufacturer’s protocol.
Western Blots Analysis
Cells were cluttered at density 1x106 cells/ml in six-well plates with DMEM medium. The cells were washed with PBS and lysed with 1X RIPA buffer containing protease inhibitor cocktail on ice for 5 min. The Bio-Rad Bradford assay kit determined the concentrations of protein. Cellular proteins from the supernatant were dissolved in an SDS-PAGE sample solution and denatured by boiling at 100°C for 5 min. The protein suspension was separated on a 12% SDS-PAGE gel and was transferred to a blotting nitrocellulose membrane for 7 minutes at a constant voltage of 25V using the iBlot® Gel Transfer Device (Invitrogen). The membranes were blocked for one h in buffer (1X TBS, 0.1% Tween-20 with 5% w/v nonfat dry milk). Membranes were incubated with anti-P53 (1: 1,000 dilution), BCL2 (1: 1,000 dilution) and anti-GAPDH (1:1000 dilution) antibodies following the manufacturer’s protocols. The membranes were incubated with anti- Rabbit IgG H&L (HRP) secondary antibodies (Abcam). A chemiluminescent substrate kit (ECL Plus Western blotting Detection Reagents, GE Healthcare) was used for HRP detection on the immunoblots. The relative band intensities according to the band of GAPDH were calculated by the software of Image Lab (Bio-Rad Laboratories).
Statistical Analysis
The final data were expressed as mean values with standard errors, and the results were analysed using one- way ANOVA with Dunnett’s multiple comparisons tests using GraphPad Prism 8.1.2. p-values were considered significant at p*<0.05, p**<0.01.
Results
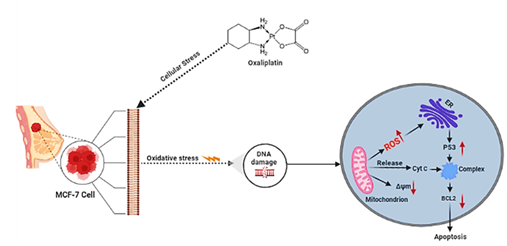
The viability of MCF-7 cells in the presence of 0.2, 2 and 20µM oxaliplatin in the culture medium was evaluated by detecting the activities of intracellular dehydrogenases (CCK- 8 assay). The viability of MCF-7 cells treated with different concentrations was confirmed to be higher than 85% after 24 h treatment (Figure 1).
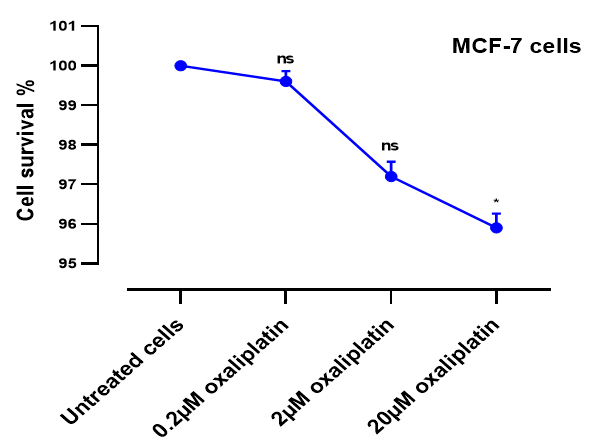
Quantitative Measurement of 8-OHdG Levels Via ELISA
We next examined the effect of oxaliplatin on MCF-7 cells using the ELISA assay. ELISA for 8-OHdG, a recognised marker of oxidative DNA damage, was used to quantify oxidative DNA damage in MCF-7 cells after being treated with 0.2, 2 and 20µM oxaliplatin (Figure 2). Results indicate that the oxidative DNA damage level significantly increased at two and 20µM compared to the untreated cells *p<0.05 and **p<0.01, respectively.
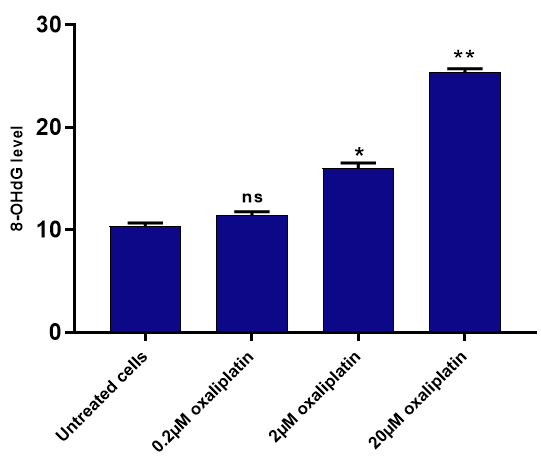
Oxaliplatin Activates the P53 Signalling Pathway Independent of DNA Damage in MCF-7 Cells
Results from the quantitative 8-OHdG levels by ELISA assays showed that oxaliplatin at concentrations two and 20µM induced statistically significant genotoxicity in MCF- 7 cells. To understand the rationale behind this effect, we further examined the effects of oxaliplatin on gene expression levels of P53 and BCL2 in MCF-7 cells at the mRNA level. The qPCR technique was used to analyse the expression of these genes. The qPCR data significantly affected these genes at concentrations two and 20µM (*p<0.05 and **p<0.01) (Figure 3).
To further explore the mechanism of the effect of different concentrations of oxaliplatin on P53 and BCL2 protein expression levels in MCF-7 cells was assessed by Western blot. The results showed that oxaliplatin significantly expresses any of these proteins in MCF-7 cells (Figures 4A & 4B). Furthermore, results from WB were consistent with the gene expression results from the qPCR.
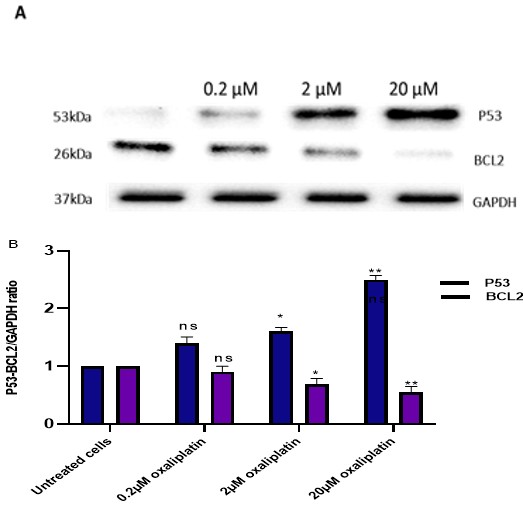
Figures 4A & 4B: MCF-7 cells were treated with oxaliplatin, and the P53 and BCL2 protein levels were examined. Sample Western blots of P53, BCL2 and GAPDH proteins are shown in (A), and quantified summary data are shown in (B). Results are the mean ± SEM From three independent experiments, and statistical comparisons were made using one-way ANOVA followed by Bonferroni’s posthoc comparisons tests, comparing each dose against the control (*P <0.05 and **P <0.01).
Discussion
Breast cancer is the most common contributor to cancer-related deaths in women. However, due to the failure and toxicity of some therapeutic cancers, tumour intrinsic and acquired resistance still exists. Therefore, it is critical to understand the molecular mechanisms underlying the appearance of this phenomenon to find ways of avoiding it and to improve and optimise treatments. Oxidative stress can induce cell death through various mechanisms, including oxidative DNA damage. The nucleobase most susceptible to oxidative stress is guanine; the most common DNA base modification formed by oxidation is 8-oxodG [11]. Therefore, we investigated whether oxaliplatin-induced oxidative stress in MCF-7 cells. Results from the quantitative 8-OHdG level by ELISA assays showed that oxaliplatin caused oxidative stress in MCF-7 cells (Figure 2). This implies that oxaliplatin might insert platinum and adduct into the nuclear DNA causing DNA damage. This implies that the relationship between all the nucleobases, guanine is the most susceptible to oxidation by ROS [12]. The formation of 8-OHdG on DNA can cause G: C—T: A mispairing mutations, which are considered having a close relationship with the development and progression of cancer cells [13]. Therefore, ROS are important signalling molecules for anticancer drug-mediated cell death. We investigated the regulation of apoptosis-related protein expression to elucidate the molecular mechanisms leading to apoptosis in oxaliplatin-treated cells. The P53 expression was significantly increased, whereas BCL2 was reduced potently in MCF-7 cells (Figures 3 & 4A, 4B). In normal cells, the expression of P53 is induced in response to stressful conditions. However, the p53 activation induces cell death by apoptosis [14]. BCL2 was previously shown to be pro- apoptotic via promoting apoptosis in platinum compounds treated breast carcinoma cells [15]. Our data displayed that oxaliplatin markedly downregulated the levels of apoptosis inhibitory factors of BCL2. This implies that the involvement of dysregulation of BCL2 in oxaliplatin could mediate apoptosis in the MCF-7 cells. A study has also shown that downregulated BCL2 expression could favour tumour survival and impart resistance to different types of cell death stimuli, including genotoxic [16].
In present study demonstrates that P53 is specifically involved in cellular responses induced by oxaliplatin, which are associated with oxidative DNA damage caused by oxaliplatin. Oxaliplatin exposure significantly increased P53, decreased BCL2 target gene expression, and increased 8-OHdG levels in MCF-7 cells compared with untreated cells. Therefore, the clinical priority is understanding how P53 harbouring cancer cells avoid the cellular death processes induced via genotoxic stress by anticancer drugs. Thus, this will develop strategies that could improve the sensitisation of these cancer cells to many chemotherapeutics agents.
Conclusion
Understanding the regulation and functionality of essential DNA repair genes and proteins involved in apoptosis regulation can potentially modulate the cell outcomes on DNA damage, specifically in the context of cancer treatment. Oxaliplatin is a third-generation clinical platinum analogue and a DNA-reactive drug. However, the cytotoxic effects of oxaliplatin are believed to be exerted through drug-induced DNA adducts.
References
-
Kashyap D, Pal D, Sharma R, Garg VK, Goel N, et al. (2022) Global Increase in Breast Cancer Incidence: Risk Factors and Preventive Measures. Biomed Res Int 2022: 9605439.
-
Wilkinson L, Gathani T (2022) Understanding breast cancer as a global health concern. The British Journal of Radiology 95(1130): 20211033.
-
Waxman DJ, Schwartz PS (2003) Harnessing apoptosis for improved anticancer gene therapy. Cancer Res 63(24): 8563-8572.
-
Johnson AG, Kranzusch PJ (2022) What bacterial cell death teaches us about life. PLOS Pathogens 18(10): e1010879.
-
Jones BM, Green S (2021) Modern radiation techniques in early stage breast cancer for the breast radiologist. Clin Imaging 80: 19-25.
-
Seo SU, Woo SM, Kim S, Park JW, Lee HS, et al. (2022) Inhibition of cathepsin K sensitises oxaliplatin-induced apoptotic cell death by Bax upregulation through OTUB1-mediated p53 stabilisation in vitro and in vivo. Oncogene 41(4): 550-559.
-
Jemal A, Murray T, Ward E, Samuels A, Tiwari RC, et al. (2005) Cancer statistics, 2005. CA Cancer J Clin 55(1): 10-30.
-
Yang C, Zhou Q, Li M, Tong X, Sun J, et al. (2016) Upregulation of CYP2S1 by oxaliplatin is associated with p53 status in colorectal cancer cell lines. Sci Rep 6(33078).
-
Tomicic MT, Kramer F, Nguyen A, Schwarzenbach C, Christmann M (2021) Oxaliplatin-Induced Senescence in Colorectal Cancer Cells Depends on p14(ARF)-Mediated Sustained p53 Activation. Cancers (Basel) 13(9).
-
Fujie Y, Yamamoto H, Ngan CY, Takagi A, Hayashi T, et al. (2005) Oxaliplatin, a potent inhibitor of survivin, enhances paclitaxel-induced apoptosis and mitotic catastrophe in colon cancer cells. Jpn J Clin Oncol 35(8): 453-463.
-
Urbaniak SK, Boguszewska K, Szewczuk M, Kazmierczak Baranska J, Karwowski BT (2020) 8-Oxo-7,8-Dihydro- 2’-Deoxyguanosine (8-oxodG) and 8-Hydroxy-2’- Deoxyguanosine (8-OHdG) as a Potential Biomarker for Gestational Diabetes Mellitus (GDM) Development. Molecules 25(1): 202.
-
Nakabeppu Y, Ohta E, Abolhassani N (2017) MTH1 as a nucleotide pool sanitising enzyme: Friend or foe? Free Radic Biol Med 107: 151-158.
-
Bowerman B (2005) Cell biology. Oxidative stress and cancer: a beta-catenin convergence. Science 308(5725): 1119-1120.
-
Yu G, Luo H, Zhang N, Wang Y, Li Y, et al. (2019) Loss of p53 Sensitises Cells to Palmitic Acid-Induced Apoptosis by Reactive Oxygen Species Accumulation. Int J Mol Sci 20(24): 6268.
-
Hong Y, Yang J, Wu W, Wang W, Kong X, et al. (2018) Knockdown of BCL2L12 leads to cisplatin resistance in MDA-MB-231 breast cancer cells. Biochim Biophys Acta 1782(11): 649-657.
-
Kim J, Bae S, An S, Park JK, Kim EM, et al. (2014) Cooperative actions of p21WAF1 and p53 induce Slug protein degradation and suppress cell invasion. EMBO Rep 15(10): 1062-1068.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells