Review: Cytohistophysiological Aspects of Blood Organ/Tissue Barriers
General physiology is the basis of basic and applied aspects of life processes and moleculo-biochemical aspects that ensures the survival of species efficiently. There is a need to maintain specific amount and numbers of metabolites in the body fluid and the fluid present in the organs. This needs a specific check on the body fluid that provides the required metabolites and removes unwanted ones. For this scrutiny, there is something like a ‘check post’ and it is referred to as ‘blood organ/tissue barrier’. Further, this ensures the restoration of appropriate ambient environment for the ongoing physiological processes in an organism. Structural, biochemical and molecular aspects of blood organ barriers have potential to illustrate on the survival mechanisms. Further, biotechnological research, artificial mode of physiological techniques and the technological advancement help to understand the intricacies of various physiological mechanisms occurring in the life activities and the remedial aspects in cases of pathogenesis. For normal physiological functioning a suitable physiological homeostasis is an important basis. The respective balanced biochemical and molecular interactions depend on processes like paracellular diffusion of metabolites, activated transport of hydrophobic and hydrophilic metabolites and drugs from the circulating blood stream. There is a specific need of an effective drug therapy along with appropriate drug delivery system that can regulate the correct dose of drug at the site to meet the remedial parameters while dealing with pathogenesis and dysfunctioning of the organ. The investigations relating blood barrier are significant while investigation the toxicological and pathological aspects of a given organ.
Abbreviations
CAMs: Cell Adhesion Markers; TJ: Tight Junction; MAGUK: Membrane Associated Guanylate Kinase; PDGFR-β: Platelet derived Growth Factor Subunit β-Receptor; TEER: Transepithelial Electrical Resistance; GFB: Glomerular Filtration Barrier; BOB: Blood Ocular Barrier.
Introduction
When a specific dye is administered in a test animal it may not succeed in reaching the site organ. This condition indicates the existence of some sort of interruption between the peripheral blood supply and the organ. Based on investigations, this physiological interruption is cytohistological and confirms the presence of blood barrier specifically for organs like brain, kidney, reproductive organs, liver, skin, eye, and most of the organs etc [1, 2, 3, 4]. This physiological check-post regulates the to-and-fro movements of biomolecules, nutrients and other unwanted molecules between the peripheral blood and the vital organs. Thus, these natural biological barriers ensure the supply of the appropriate metabolites and removal of unwanted ones to the specific organ-site as per the physiological demands and help to maintain the respective healthy physiological efficacy. This arrangement also provides a suitable cellular and ambient microenvironment that facilitates a biochemical and physiological homeostasis. Structurally, a blood barrier has a selectively semi-permeable membrane, endothelium, pericytes, capillary basement membrane, astrocytes, tight junctions and enzyme or metabolic barriers. These components are suitably architectured involving the interstitium of the concerned organ. There exists some difference between the blood of peripheral blood capillaries and blood in the related organ. Generally, a specific suitable level of homeostasis is maintained with the help of four processes. These prevent paracellular diffusion of hydrophilic metabolites, mediating the active nutritive metabolites, activate the transport of hydrophobic metabolites and drugs between peripheral blood and the organ. There is a regulated migration of pathogen and blood cells in the peripheral circulation via transendothelium. In general, there is a cytohistological, biomolecular and physiological protection against most of the xenobiotics and therapeutic drugs. These blood barriers are the specialized cytomorphological sites and are capable to perfect selective transport of metabolites in a specific organ during the systemic circulation. In general, epithelial and endothelial cells are the cytological components in a blood barrier which exhibits some of the specific adaptations which facilitate the formation of intercellular junction. This association involves “tight junction” and act as intercellular connections that seal the intercellular cleft [5, 6]. Normal functional blood barrier is a dynamic site manifesting suitable physiological behaviour adapted to respond to the specific stimuli corresponding to the selectively control biomolecules from vascular circulation to the organ and reverse. These structural and functional aspects are specifically related to therapeutic drugs/ agents to ensure appropriate drug delivery and prevent the interference of unwanted metabolites [6]. The inter and intracellular coordination and communication bring about the basic cellular sustenance and the social dynamic cellular functionality in an individual. Thus, each cell has a potential to receive, metabolize, distribute and maintain the required level of homeostasis for its successful performance and ability to respond to the changes in the respective ambient environment [7, 8]. Further, it seems quite rational to understand the molecular basis of the cellular communications to maintain a healthy cellar status, development, aging and remedial therapeutics. The investigations related to molecular heterogeneity in ‘omics’ significant helps to understand cellular status and its functionality. The microengineering and high throughput genomics elaborate spatiotemporal control of the cellular interactions. Any type of disruption in the cellular communication is likely to results in metabolic, biochemical malfunctioning that it is likely to lead to pathological conditions like tumorigenesis, metastasis, and senescence [9].
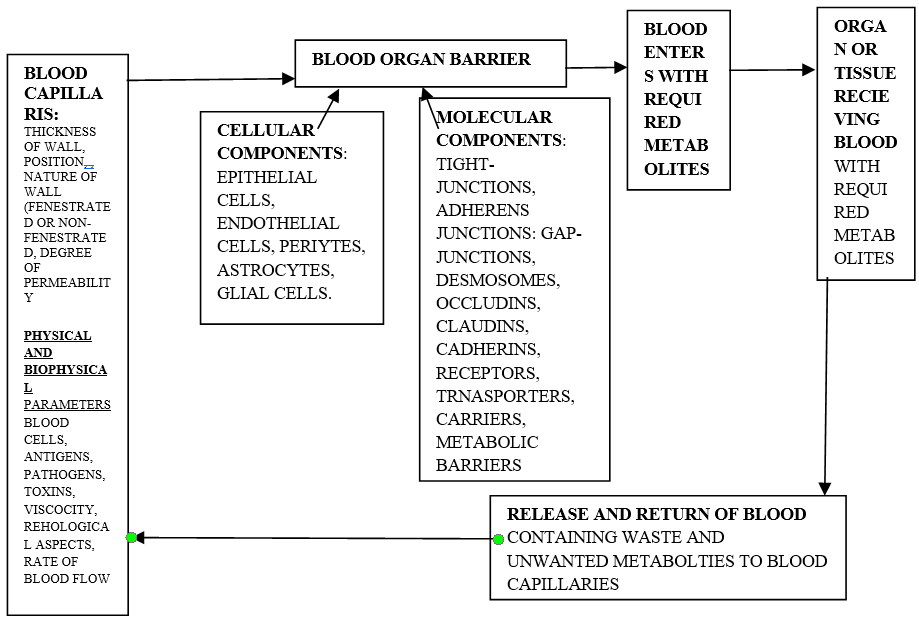
Materialistic nature of blood and the biological nature of organ play significant roles during structural aspects and functionality of a blood barrier parameter. The blood components i.e., plasma along with its inorganic and organic constituents and the cells present in blood exhibit biophysical mechanics and influence the blood behavior. The biochemical and cellular components of blood i.e., proteins like albumin, globulins or fibrin, etc., leucocytes, platelets and erythrocytes and biological and non-biological antigens also affect the blood behavior. The behavior of these blood constituents depends on the fluid status of plasma and serum. Thus, blood components experience relatively wide range of biophysical forces like hydrodynamic force, rheological forces- shear stress and shear rate (deforming strain), forces due to viscosity of plasma, forces due to the specific shapes of cells and molecules flowing in plasma. These parameters also vary during healthy and any pathological status of an individual. The blood vessels offer specific form of force on the blood flowing through them; thus, the blood vessels of any size act as mechanosensitive barrier. The status of blood vessels i.e., stiffness and thickness of the wall of blood vessels, add to the shear stress on the blood flowing, it also correlates with the plaque formation within the blood vessels. It becomes obvious that while designing vascular grafts one should consider the mechanical homeostasis, correlation between the effects of fibrin and plaque formation and forces due to the impacts of platelets and the vascular and haematological status of the individual [10, 11, 12]. Investigations related to various blood barriers indicate the need for involving a functional vertebrate model to mimic the concept of functioning of blood barriers; such investigations also help to investigate some of the related diseases and pharmacodynamics and pharmacokinetics testing [13, 14]. There are, however, significant challenges due to mimicking human disease in animals, as many models do not completely recapitulate human disease manifestations and instead allow only for studies of certain disease aspects [8, 13]. However, the use of animal models is of particular importance for pharmacodynamics and pharmacokinetics testing, where the effects of pharmaceutical interventions can be examined at the systemic level to determine drug safety and efficacy before entering human trials phase.
Basic Concept of Blood Barriers
In different organs the components of blood barrier remain structurally and functionally similar with suitable modifications. The investigation related to blood barrier is of significance while studying toxicological and pathological aspects in a given organ. Normal functional blood barrier is a dynamic site exhibiting physiological behavior that is suitably adapted to the specific stimuli corresponding to selective permission and prevention of biomolecules from vascular circulation to the tissue and reverse. Each prime organ requires essential micro and ambient environment to maintain its specific assigned physiological functionality and blood barrier has its exclusive role accordingly. These specifications depend on the homeostatic, biochemical and neuronal-physiological conditions of the organ and blood barrier helps in maintaining such conditions. The rate of blood flow, degree of vessel permeability, physicochemical properties of the biomolecules, drugs, unwanted molecules and cellular blood components undergo suitable changes or variations to attain the suitable healthy physiological environment. These structural and functional aspects are specifically related to therapeutic drugs/agents to ensure appropriate drug delivery. It will be very apt to investigate the kinetics of pharmaceutical drugs in this context. Basically, the epithelial cells are mesodermal in origin and modified and suitably form the wall of blood vessels (endothelium). These cells are dedicated to permit strict and controlled the to-and-fro movements of hydrophobic, hydrophilic and ionic metabolites between peripheral blood and the tissue. The proteins of tight junctions namely, claudin and occludin, limit the paracellular molecular movements. The cells of endothelium are structurally and functionally suitable to acts as a selective barrier and to regulate the flow of biomolecules traversing between blood and the tissue of the organ. During stressful conditions, there is an increased permeability resulting in edema and disruption of the functional aspects of the organ. This further interferes with the molecular mechanism and maintenance of the specific hemostatic condition. The newly developed condition of vascular hyperpermeability interferes with drug involved as pharmacological corrections [15]. The overall degree of permeability of blood barriers fluctuates under normal and pathological conditions. In case of neurological and clinical conditions like cerebral ischemia, brain trauma, there is a relatively higher degree of permeability that facilitates the entry of drugs. If, due to some reason the degree of permeability declines, then the drugs will lose their therapeutic efficacy and hamper the recovery. Similar aspects are true in case of other organs like liver, gonads, lungs etc [16] (Figure 1). Endothelial cells and pericytes (also described as perivascular cells), are the two primary structural, functional and interacting components of blood vessels. The endothelial cells constitute the inner lining of a blood vessel while pericytes are smooth muscle cells (also referred mural cells). These cover the vascular surface of the blood vessel. These cells along with their other cellular components establish the structural aspects and the functionality of blood vessel with respect to regulation, formation, proliferation, survival and migration but the mechanism of such coordinated morphological regulation specifically the tubular sprouting, fusion and formation of vascular network is ambiguous but the vascular endothelial growth factor is the prime factor which controls the formation and functions of blood vessels. This vascular endothelial growth factor acts as an important factor related to angiogenic sprouting and guides the formation of extension of filopodia from the specific apical endothelial cells. The cells of the apical region of vessel work in response to the vascular endothelial growth factor for proliferation and migration of the newly formed vascular sprout stalks. These endothelial cells give antagonistic response to vascular endothelial growth factor-A and vascular endothelial growth factor receptors-2. The apical cells show dependence for their efficacy during migration and proliferation on the gradient and concentration of vascular endothelial growth factor-A and these help to attain balanced effectiveness of endothelial cells [17]. The solutes that involve active efflux carriers undergo passive diffusion through endoplasmic epithelial cells. The polar molecules such as gluconucleosides and amino acids move across such membrane due to selective transportation. Macromolecules like proteins and peptides make use of ‘receptor-mediated-transcytoses’ to move to-and-fro across the membrane. Some of the common transporters and receptors are excitatory-amino-acids transporters 1-3, L-system of large neutral amino acids, and P-glycoproteins. Some of the common receptors and carriers that are associated with endothelial cell membrane are 5-hydroxytryptamine (serotonin), angiopoietin-1, basic fibroblastic cells, endothelium-1, glial cells, derived trophic factor-β, endothelial specific receptor tyrosine kinase-2. The cell culture techniques are most suitable for the related cellular investigations [18]. Pericytes develop from mesoderm and originate from mesenchymal cells. These cells invade endothelial tubes during their development and formation. The pericytes exhibit different morphological appearances and perform different functions too. These are associated with only one tissue and single capillary bed. Primarily, these modulate the flow of blood, vascular permeability, stability and growth of vessels [19]. The pericytes are smooth muscle cells (mural ells) and cover the vascular surface of the blood vessel. These cells have contractile fibres like smooth muscle fibres. Typically, the body of a pericytes has a well distinguished nucleus, relatively less cytoplasm and many elongated processes that embrace the abluminal wall of endothelial cells [19, 20]. When pericytes are absent in a blood vessel, hemorrhagic and hyperdilated conditions are not observed in such cases. These conditions result in edema, embryonic lethality and diabetic retinopathy. Pericytes also participate in angiogenesis in tumours and as a result these are the targets during antiangiogenic therapies. The pericytes are complex cellular components of blood vessels and their advanced functionalities and ontogeny need further exploration. It is very exciting topic for investigation in relation to growth, maintenance of blood vessels and also during pathological angiogenesis [20]. These pericytes play significant role during cellular communication with endothelial cells ensuring their physical contact and the success of pathways involved during paracrine signalling. This communication directly involves the gap junctions to ensure exchange of ions and smaller molecules. Pericytes anchor with endothelial cells. The peg and socket contacts enable the cellular penetration but discontinuities are present in the basement membrane of blood vessels. The mechanical contractile forces transmit through these junctions between pericytes and endothelial cells. Additionally, molecules like N-cadherin, cell-adhesion molecules, β-catenin based molecules of adhesion junctions of extracellular matrix like fibronectin also play significant role during these communications [17]. Astrocytes have distinct body and the end feet; the body associates with the tissue of the organ of the location of blood barrier while the end feet link with tight junctions of the epithelium cells and establish a firm link with tight junctions. Their functions relate to barrier is unclear but their ability to maintain the established firm contact [21]. The cell membrane of epithelial cells has high numbers of tight junctions that restrict or prevent paracellular flux of hydrophilic biomolecules. There are either least or no fenestration and few pinocytic vesicles present that help the movement of metabolites across. The roles of pinocytes present in the basement membrane play significant role during angiogenesis and relates with the structural and developmental differentiation of microvessels [22]. Blood capillaries present around circum-tissue of the organ within the specific organ and these exhibit fairly good degree of fenestration. This structural aspect facilitates selective diffusion of metabolites between circulating blood and the tissue of the respective organ. Tight junctions are present in between the endothelial cell and show highly selective barrier efficacy. These also achieve effective diffusion which permits least metabolites into the concerned organ. Tight junctions are made of specific proteins that form a direct contact and form “tight seal’. This site provides one way passage to the metabolite i.e., the metabolites move selectively from peripheral blood into the specific organ/tissue. The transmembrane proteins such as claudin, occludin types measuring around 3-4 nm in diameter are present and are arranged in interlocking manner as either claudin or occludin types. In addition to these proteins the junctional adhesion molecules establish intercellular contacts proteins. These proteins prevent leakage of metabolites and prevent the passive diffusion of proteins and polar solutes across them [6, 23]. The tight junction proteins also interact with zonula occludin proteins, actin cytoskeleton, protein kinases, small GTPases and heterotrimeric G-proteins present in the membrane [23, 24]. The adherens junctions are adhesion complex, mostly seen very near to the apical membrane in epithelial cells. The adherens junctions appear as minute link-sites, these junctions involve transmembrane E-cadherin (calcium dependent adhesion), intracellular components, p-120 catenin, β-catenin and α-catenin. These protein molecules act as loading or linking component between cells surface and cytoskeleton. Such junctions facilitate maturation and maintenance of the junctions [6]. These junctions also help to establish the structural integrity of other junctions like tight junctions involving transmembrane and cytoplasmic plaque proteins [25]. These also help to make mechanical microdomains at the cellular interfaces which facilitate the physiological aspects of these junctions [26, 27]. Other protein components present in adherens junctions are vascular endothelial-cadherin, scaffolding proteins catenins, p120 and plakoprotein. The vascular endothelial-cadherin manifests the cell-to-cell adhesion [28, 29]. The vascular endothelial-cadherin protein exhibits two domains, extracellular and cytoplasmic domains. Generally, extra cellular domain links with the extracellular domain of the neighbouring endothelial cell while cytoplasmic domain connects with actin filaments involving scaffolding proteins. The catenins, p120, plakoprotein physically abridges with zonula accludens-1, and actin filament; zonula accludens- 1is a protein component of tight junction [30, 31]. Platelet and endothelial cell adhesion molecule-1, CDPP (circulating dipeptidyl peptidase) and nectin also play structural and functional roles in the adherens junction. In most of the blood barriers adherens junctions are responsible for their structural and functional integrity [6, 32].
Basic Concept of Intercellular Communication in Blood Barriers
The three-dimensional aspect of cells constituting a tissue along with extracellular matrix, intercellular placement, flow of interstitial and vascular fluids plays major roles during the process of cellular communication. There are possibilities that the speed and the components in the fluid around the cell may show difference in comparison to the speed and composition of components in fluid moving through the main vessels. This difference may be due to the different physiological requirements of different organs. For example, liver is a site for detoxification, metabolic synthesis, formation of bile, bile salts, and bile pigments; pancreas is the site of synthesis of insulin, glucagon; gonads are the for formation, storage and maturation of gametes; kidneys are the site of selective excretion, etc. Similarly, some differences should be expected in case of targeted pharmaceutical products, metabolic fuel or products, metabolites and/or components of immune system, antigens and any biological or non-biological product which is not recognized as ‘self’ by the cellular components of blood barriers [33, 34]. There is a possibility of change in the degree of membrane permeability of the cells of blood barrier in accordance to the change and speed of flow of the ambient body fluid and vascular blood [35, 36]. Epithelial membrane in case of blood barriers have ability to express varied transporters that help to perform successful movements of selected molecules including multidrug molecules across the membrane. Disrupted functioning of such transporters and membranes are one of the prime factors for development of inflammation and tumors. This functional defect needs specific remedial strategy to ensure the proper functioning of the transporter and the membranes involved. Generally, in such cases, a co- application of the respective drug along with mediator such as bradykinin or mannitol in hyperosmolar concentration appears to be a good option. Other options are lipidization of the test molecule, enclosing the molecules within liposomes or coupling the molecule which can readily enter in the cell. Thus, the understanding of physiological aspects of blood barrier helps to retain high efficacy of drug and the strategic applications [37].
Epithelia with barrier-function typically have dense intercellular junctions along with few pinocytotic vesicles. There are many transporters for the selective transportation and for the exchange of molecules. One of the groups helps to transport multidrug in a controlled manner. Commonly, inflammations and tumors cause disturbances in the blood-tissue-barriers. Strategies available for drug delivery to tissues with barriers include the opening of the barrier and the modification of the drug. The increase on the permeability can be achieved using the co-application of the respective drug with mediators as bradykinin or hyperosmolar concentrations of mannitol. Modifications of the drug include lipidization of the molecule, enclosure into liposomes and coupling to substances that are actively taken up by the cells. The pharmaceutical treatment of organs with blood-tissue barriers requires both an efficacious drug and special application strategy [37, 38]. In general, there are three modes of effective direct intercellular communications. These are enumerated (i) direct cell to cell engaging biomolecules exchange through cell-to- cell contact, (ii) cells can undergo polarization developing tension which results in mechanical communication, and (iii) cells can develop mutual binding using ‘cell adhesion markers’ between the interacting cells and the cell whose gap junctions have association with the cytoplasm of the neighbouring cells. The interacting cells can develop fine projections forming a tube-like structure (nano-sized) called ‘tunnelling nanotube’. All these cellular modifications ensure intercellular communication in a biosystem. Utilizing cell adhesion markers (CAMs) in this complex process involving different transmembrane proteins, like selectins, integrins, cadherin, and specific group of immunoglobins having very high cellular specific cell functions. These proteins facilitate homophilic and heterophilic interactions [9, 38, 39, 40, 41, 42] (Figure 2).

The other mode of biochemical intercellular communication is indirect in nature. Molecules mostly move due to the processes like body fluid movements, diffusion or due to cellular deformation some strain is induced in extracellular matrix. Generally, minute hydrophobic soluble signaling molecules diffuse passively through plasma membrane, when these are unable to move through membrane then hydrophilic molecules participate. Further, the specific membrane receptors identify such molecules and facilitate their journey across the membrane into cytoplasm. Even processes like pinocytosis, endocytosis, and carrier proteins also play a significant role during such molecular movements through the membrane. The nature of the signal is an important parameter that judges the soluble factor signal and the distance between target cell and the effector. The paracrine signals induce responses when these are very close to the target cell. The autocrine signals are the product of the distant cells and if not effective, these reach back to the cells of origin. The vascular blood acts as a mediator for the endocrine cellular communication. The lipids, proteins, exosomes, microvesicles and genetic mater reach their destination through extracellular vesicles [9, 40, 43, 44, 45, 46, 47]. In addition to biochemical signalling, the communicating cells and their location induce mechanical forces on the surrounding cells. The extracellular matrix and the cytoskeleton perceive these stimuli as the induced physical forces and result in rearranging, remodelling their orientation in accordance to epigenetic modes. [Epigenetics relates with the study of impacts on behaviour, environment and the effects of genes. These modifications are temporary and do not affect the DNA sequence of an individual; they only change the pattern of the body i.e., how it reacts in congruence with the induced changes]. The extra cellular matrix exhibits modifications like non-linear mechanical effectiveness in its proteins and increases the strain. These modifications and increased strain induce the cells to communicate with the surrounding cell placed within five cell diameter distance. The indirect mode of mechanical cellular communication affects the cellular behavior, cell migration, enrolment of immune cells, forms special cellular structures and network in endothelial cells [9, 47, 48, 49, 50, 51] (Figure 3).

Molecular Aspects of Junction Proteins
In general, epithelial cells form the internal and external surface of an organ in an organism and act as selective barrier-interface between the organism and the internal as well as external environment. The selective nature of these interfaces acts as defensive cover against pathogens, and as a regulating device against unwanted transport barrier for metabolites like, ions, fluid, solutes, toxicants and pharmaceutical drugs along with thermos-regulation. The efficacy of this selective cytological hinderance relates with the successful functioning of the biomolecular conjugations like intercellular junctions such as tight junctions, subjacent adherens junction; these collectively known as the apical junctional-complex. The desmosomes and the gap junctions are present inner to this apical junctional-complex and effectuates the intercellular adhesion and appropriate molecular cross communication between adjacent cells [52]. Tight junctions are made of three main families of hepatic tight junction proteins namely, claudins, tight junction-associated proteins MARVEL proteins (microtubule associated for vesical trafficking and membrane link), (TAMPs- a family of proteins having four transmembrane helical structures and help in the formation of diverse membranous subdomains; these are referred to as myosin and lymphocyte related proteins involved in the MARVEL domain proteins MAL [(myosin and lymphocyte) and related proteins for vacuole formation and membrane link] represent a family of proteins with four transmembrane helical structures and mediate the formation of diverse membrane subdomains Hernandez V, et al. [53], of vacuole and membranous [54]. TAMPs proteins are the product of occludins, MarvelD3 and tricellulin and receptors of pathogens [55, 56]. Primarily, most of the tight junction proteins maintain the normal fabrication and coherence of tight junctions while the family of TAM-proteins affect the functional barrier capabilities involving the critical signalling capabilities of occludin and thus, accomplish the epithelial homeostasis [57]. Interactions between different tight junction (TJ) proteins and the cytoskeleton are essential for the normal assembly and maintenance of TJ integrity. While the TAMP family of proteins has been implicated to influence barrier function, proteins such as occludin also have critical signalling properties that regulate epithelial homeostasis [58]. Itch- a response that causes strong desire to scratch. Itch- comes under HECT type E3 ubiquitin ligase and it controls the autoimmune developmental diseases among mice and humans. It also monitors the differentiation of T-cells to tumorigenesis in hepatic cells]. There is a molecular inter action involving E3 ubiquitin-protein ligase itch; this may facilitate the degradation of occludin which relates to the inhibition of proteosome [58]. Most of the transmembrane proteins of tight junctions are of tetraspanins type that are characterised by transmembrane domains which form two extracellular loops. Further, the C-terminal present in the cytosol constitutes a complex when it joins with adaptor proteins like zona occludens-1. When these adaptors, coordinate with other proteins, then these harbour in actin cytoskeleton. The zona occludens proteins come under the scaffolding types of protein; these proteins are responsible for structural aspects of multiprotein complexes present on the intercellular junctions located on the surface of cytoplasm. These proteins associate with integral membrane proteins and also with filamentous cytoskeleton. These proteins associate with membrane guanylate kinase (MAGUK-like proteins with different numbers of domains but have similar sequence. Zonula occludens proteins establish cell to cell structural contacts, these also regulate cell growth and proliferation [59]. ZO (zonula occludens) proteins are scaffolding proteins providing the structural basis for the assembly of multiprotein complexes at the cytoplasmic surface of intercellular junctions. In addition, they provide a link between the integral membrane proteins and the filamentous cytoskeleton. ZO proteins belong to the large family of membrane-associated guanylate kinase (MAGUK)-like proteins comprising a number of subfamilies based on domain content and sequence similarity. Besides their structural function at cell-cell contacts, ZO proteins appear to participate in the regulation of cell growth and proliferation [59]. When there is a barrier deficiency, F-actin and misexpression associates with elevated viral entry and dysfunction of cell adhesion occurs [60].
Adherens Junctions
Cadherins are a type of trans-membrane proteins that participate in the functioning of adherens junctions and positioned very close to tight junctions at the basal zone of lateral plasma membrane. These are present in epithelial cells and responsible for homo-type interaction. One end of these protein molecules associates with cadherins of other cells and the other end towards the cytoplasmic end links with catenins and actin cytoskeleton forming cell to cell adhesion. Beta-catenins of adherens junction binds with E-cadherins and actin of cytoskeleton involving α-catenin. Beta and α-catenins help in the post-translation of E-cadherins and its trafficking [61]. Adherens junctions also affect the efficacy of ‘blood bile barrier’. Loss of α and β-catenins cause alteration in bile canaliculi, inflammation, degeneration or loss of microvilli, jaundice and cholestasis (phenotype) and increase of basal hepatic bile acids and mild cholestasis [62, 63, 64].
Gap Junctions
These junctions connect two adjacent cells and control the inter cellular movements of selected ions, molecules and electric impulses. The gap junctions have connexins group of proteins as a primary constituent; these interact with occludins of tight junction. When these associate with ZO-1 proteins they facilitate the integrity of blood bile barriers. The gap junctions are the major targets for the drug induced liver injury like acetaminophen (inhibition of connexin-32 affects impact of acetaminophen) and are potential proteins that relate with drug safety phenomena [65, 66, 67]. Environmental toxins influence functioning of these inter cellular junctions while drugs (pharmaceuticals in excess doses) induce injury and/or chronic or acute pathological conditions [63, 64].
Desmosomes
These intercellular connections articulate the adjacent cells involving coupling desmosome cadherins to intermediate filament present in cytoskeleton. Structurally, these are made of desmocollin, desmoglein, and γ-catenin. Although there are few references but these play significant functional and structural roles and are helpful to either tight junctions or adherens junctions. All together these are quite relevant in the process of intercellular articulative cytophysiology [6]. The lake or deficiency of β- and γ-catenins adversely decline E-cadherin, occludin, and claudin-2 and as a consequence dysfunctioning of blood bile barrier and induction of progressive intrahepatic cholestasis occurs [64, 65, 66].
Physiological Aspects of Blood Barriers
The histological and cellular component of blood barrier sustain controlled physiological well-suited microenvironment in the specific organ and ensures its optimal biophysical, biochemical, biomolecular. and physiological functionalities. This cellular device is capable of selective exchange of ions, biomolecules, therapeutical drugs, over-all hemostasis and hemostasis in a given organ. Homoeostasis relates with the physiologically optimum stable suitably regulated state in an organ and ensures the normal well-coordinated, synchronized functions of an organ. The hemostasis basically regulates the composition of blood so that proper supply, use and elimination of unwanted metabolites and availability of various required metabolites is maintained. The blood barrier is responsible to provide protection, facilitates to-and-fro transport of the essential and unwanted wanted metabolites. It regulates inflammation, immune response and role of specific ion channels. It also facilitates the responses to the inter and intracellular communication, maintenance of electrochemical static stability (gradient) and fluctuations due to pathological alterations [67, 68, 69]. The asymmetrical distribution of ions along the luminal and abluminal membranes of the cells play functional roles in working of blood barrier. The Na-K ATPase regulates the influx and efflux of Na+ and K+ ions against concentration gradients and maintains electrochemical gradient across the concerned cell membrane. The co- transporters are the proteins which help in the secondary active transport of sodium, potassium, and chloride ions into the cell; the Na-K-Cl co-transporters regulate the cellular ionic balance of Na+, K+, and Clˉ. When the ionic balance gets disturbed resulting in pathological and dysfunctional states. There are transporters to carry solutes, ATP-binding case families that mobilize essential biomolecules, nutrients needed for the homeostasis of the respective organs. Various signalling pathways like cytokinases, Janus kinases (intracellular, non-receptor tyrosine kinases and signal mediators), phosphoinositide-3-kinase, myosin light chain kinase pathways, phosphoinositide-dependent kinase, and protein kinase-G, along with others take active part in the active transport of biomolecules [9, 67, 68, 69, 70, 71].
Blood Barriers
The blood organ barriers although have basic structural cellular components but these vary in different organs like brain, kidney, reproductive organs etc. The following description relates with basic components of blood barriers in different important organs.
Blood Brain Barrier
This barrier has been discussed extensively and still the pursuit continues. Mostly reports are based on its location, structure and physiology in mammals. This barrier is well developed and functional system. It consists of dynamic semipermeable membrane which is efficient to differentiate most of the unwanted metabolites and microbial agents. It is solely responsible to provide a haemostatic ambient physiological environment in brain. The specialized and organized tissue of blood brain barrier includes arachnoid epithelium, middle layers of meninges, epithelium of choroid plexus and tissue secreting cerebrospinal fluid. The cells of this site have many tight junctions which regulate the permeability of the paracellular path-ways that take place in intercellular cleft. The circum-ventricular organ also have specialized neurosecretory and/or chemosensory cells and leaky endothelium. This type of cellular setup facilitates exchange of metabolites from the circulating blood. A specific external glial barrier differentiates this arrangement of tissue from the rest of the brain tissue and the barrier from cerebrospinal fluid present at ependyma [5]. The cellular constituents of blood barrier are endothelial cells of capillaries, basal lamina and perivascular-end feet of astrocytes which encircle the endothelial cells. The distance between capillaries and the neural cells is around 25 µm. There is a copious supply of capillaries at the site of blood brain barrier [72].
The basic cellular functional aspects of brain barrier include cells of endothelium, pericytes, astroglial (astrocytes and glial cells), and complexes of the cellular junctions like tight and adherens and these play major roles in functionality of blood brain barrier [69]. The astrocytes provide a structural and functional contact with the neurons involving pericytes and microglial cells. The related cell culture study indicates the involvement of many transporters and receptors like excitatory amino acid transporters 1-3, glucose transpoter-1, L-system for larger neutral amino acids and P-glycoproteins. The two-way inductors are present between astroglial and endothelial cells that help in establishing and maintaining the proper functionality of blood brain barrier. Some of the receptors and transporters present on endothelial cells are 5-hydroxytrypamine (serotonin), angiopoetin-1, basic fibroblast growth factor, endothelin-1, glial cell line derived neurotrophic factor, leukaemia inhibitory factor, purinergic receptor (P2Y2-under G-protein coupled receptors), transforming growth factor-β and endothelium-specific receptor kinase-2 [69]. Endothelial cells are involved in blood barrier (specifically in brain) deviate structurally and physiologically in comparison to other peripheral endothelial cells. These cells do not have permeability functionality as their innate feature. Good numbers of tight and adherence junctions bring about the intercellular connections forming two distinct membranous partitions namely, luminal and abluminal. There are no fenestrations or minute trans- cellular pores; hence, these cells exhibit highly restricted free and rapid cellular exchange of molecules in the region between the capillary blood and the surrounding tissue. The number and the distribution of mitochondria are higher and dense in these endothelial cells. These provide the energy for the physiological activities. Their typical cellular behavior shows three main features that make them more suitable for the designated functions; first is the development of negative surface charge, this helps them to prevent the entry or interaction with negatively charge molecules and adherence of molecules that prevents the adherence of leukocytes and stop the immune cells to enter, second: the endothelial cells have limited number of transporters; this feature facilitates the regulation of the specific substrates; third: such endothelial cells have the ability to develop high transendothelial electrical resistance as a result there is a restriction on the to-and-fro movements of transcellular vesicles via the wall of the blood vessel. These features of endothelial cells maintain a physiologically suitable ambient environment and prevent the movements of unwanted molecules across the specific zone [69]. Mostly two types of transporters (efflux) are functional in epithelial cells of blood barrier; firstly, the polarized luminal surface brings about the movements of diverse lipophilic biomolecules which intend to diffuse towards blood from the tissue. The second (efflux) is about the transporters that help the movements of highly specific nutrients from the blood into tissue; these transporters also remove the waste molecules from brain to blood [73, 74, 75, 76]. Astrocytes are the cellular component of blood brain barrier; these are responsible for bringing about signalling and biomolecular communication related to elimination of waste, controlling flow of blood, other vascular functions, and ionic homeostasis. These astroglial cells also ensures organised and balanced neuroimmune responses. The astrocytes are a type of neural cell and present in large number deeper within the tissue. In case of brain, the astrocytes differentiate as protoplast localized in well vascularized grey matter while other are distributed in fibrous form and are present within less vascularised white matter. Their end feet connect with basement membrane. The specific binding proteins like aquaporin-1V and ‘dystroglycan-dystrophin complex’ along with proteoglycan agrin that plays a major structural and functional role in this intercellular link. The astrocytes and pericytes embrace the blood vessels and act as interface between endothelium and neurons. In case of central nervous system of invertebrates in which vascularized, no circulatory system, these astrocytes function as barrier between the tissue of central nervous system and the humoral body fluid. The functional aspects of astrocytes vary in healthy conditions, participate in astrogliosis (a process in which series of changes take place in astrocytes; these changes are molecular, cytological and physiological and in accordance to the level and/or varied pathological neuro-disorders) including formation of glial scar. These play significant roles during the formation of lesions and course of specific pathological conditions [69, 77, 78, 79, 80]. Mural cells are heterogeneously distributed in the vascular system and include smooth muscles and pericytes. These cells are the structural and functional aspects of microcirculation in metazoans. This set-up plays a major role in controlling blood distribution, blood pressure and integrity of vasculature of the organ and the region. Their dysfunctions have potential to cause derogative impacts on progression of pathogenesis, cardiovascular diseases like formation of atherosclerotic plaque, and degenerative neural diseases like Alzheimer’s and Parkinson’s disease. There is a possibility that it helps during signalling between the brain blood barrier and the neural tissue, specifically PDGF (platelet-derived growth factor; platelet derived growth factor receptor-A. It is also known as CD14a; it is present on the surface of wide range of cells and participate in cell signalling specifically during the process of growth of cell and the differentiation of cell), ‘Notch and signal’ related to inflammation, and the signalling in these neural degenerative disorders. These signalling pathways are also effective in conserving cells of mural tissue and cardio and neuroprotection [69, 81]. Pericytes are another cellular component of mural cells (earlier known as rouget cells). Pericytes are the cellular communicating bridge between the endothelial cells and basement cells (membrane), ensuring physical and paracrine communication and signalling. The pericytes make adjustments in the blood flow within cerebral region, development, maintain vesiculation and control inflammation in neurons. The pericytes embed themselves at the abluminal site of the endothelial cells. These pericytes enclose around full length of endothelium of central nervous system. The location of pericytes facilitates the intercellular communication between them and endothelial cells. Primarily, the platelet derived growth factor subunit-b (PDGF-B) pathway plays a significant role during such communications. During the cellular communication/ signalling process, the endothelial cells produce ‘derived growth factor-B’ (PDGF-B) which binds with platelet derived growth factor-subunit β-receptor (PDGFR-β) present on pericytes; thereby ensures the association of pericytes with blood vessels. During this cellular communication the pericytes release molecular signalling factors and these factors influence the endothelial cells depending on the number of tight junctions present and also change the membrane charge on the end-feet of astrocytes. Thus, there is a molecular communication and also change in electrical charge at the site of this cellular association. Further, the number of pericytes play important role in the number of tight junctions present at the site [69, 82].
Tight junctions: These intercellular connections help to maintain specific level of permeability, tissue homeostasis, and morphological aspects of the cellular status in a given organ. It is not only a cellular connection but also participation in the cellular physiology as per its location. Mostly, these interconnections are seen between endothelial cells forming ‘interendothelial cleft’ and act as a regulating entrance for paracellular permeability and it also facilitate lateral diffusion specifically pertaining to proteins of integral membrane, along with lipids. This process helps to maintain the polarization of cells. Primarily, proteins like claudins and occludins ensure a firm intercellular connection. Of these two proteins claudins participate as barrier component while occludins take care of firmness of the tightness junction [6]. In addition to these proteins the lipolysis- stimulated lipoproteins receptors, junction adhesion molecules, calcium/calmodulin dependent serine protein kinase, monoclonal antibody 7H6, heterotrimeric G-protein, and tricellulin proteins are also the structural components of tight junctions. The adhesion molecules bring about initial attachment during the development phase and the kinase like proteins retains the specific level of permeability in the complex of blood brain barrier. The interactions among these proteins result in physical sustenance of tight junctions [69, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92]. Adherens junctions: These junctions are also referred to as intermediate junctions and belt-desmosomes based on their ribbon like appearance. Their location of attachment appears as adhesion plaque. These junctions associate with tight junctions specifically in the epithelial tissues at the basal region. These junctions are also seen in non-epithelial tissues such as cardiac myocytes as facial adherens. These ensure synchronised functioning. These junctions exhibit varieties in their forms such as, focal adhesion, hemidesmosomes. The cellular cytoplasmic face links with actin components of cytoskeleton or ambient extracellular matrix. The adherence junctions are responsible for the maintenance of tissue integrity and help to withstand the mechanical stress/ stretching [6]. This molecular assembly acts as a provision to maintain the structural aspects and functionality of tight junctions at cellular interfaces as these units reorient spatially, chemically and mechanically microdomains at the specific sites. Adhesion junctions and tight junctions get associated with cytoskeleton engaging transmembrane and cytoplasmic plaque proteins. The compositional proteins of adhesion junctions are vascular endothelial-cadherin, scaffolding proteins-catenins, scaffolding proteins p120, plakoprotein. Vascular endothelial cadherin is a homo- dimeric transmembrane protein and ensures the intercellular adhesion involving extracellular domain. These extra cellular domains effectively associate with the same or similar molecules of surrounding endothelial cells, specifically, at the paracellular cleft while its cytoplasmic domain chemically links with interacting filaments of actin engaging proteins of related scaffolding. The catenins, p120 and plakoprotein form a molecular bridge which connects with the protein of tight junction-zona occludins and filaments of actin, thereby providing the necessary physical support. Other proteins molecules related to platelets and endothelial cells adhesion, CD99 and nectin are also concerned with adhesion junctions. Thus, the integrity of adhesion junctions plays an essential structural and functional role in the inter-endothelial interconnection [28, 29, 93, 94, 95]. These junctions appear like a ribbon hence, also referred to as intermediate junctions and belt-desmosomes. Their location of attachment appears alike adhesion plaque. These junctions associate with tight junctions specifically in the epithelial tissues at the basal region. These junctions are also seen in non-epithelial tissues such as cardiac myocytes as facial adherens and ensure synchronised functioning. These junctions exhibit varieties in their forms as focal adhesion and hemidesmosomes. The cellular cytoplasmic face links with actin components of cytoskeleton or ambient extracellular matrix. The adherence junctions are responsible for the maintenance of tissue integrity and help to withstand the mechanical stress/stretching [6]. This molecular assembly acts as a provision to maintain the structural aspects and functionality of tight junctions at cellular interfaces as these units reorient the spatial, chemical and mechanical microdomains at the specific sites. Adhesion junctions and tight junctions get associated with cytoskeleton engaging transmembrane and cytoplasmic plaque proteins. The compositional proteins of adhesion junctions are vascular endothelial-cadherin, scaffolding proteins-catenins, scaffolding proteins p120, plakoprotein. Vascular endothelial cadherin is a homo-dimeric transmembrane protein and ensures the intercellular adhesion involving extracellular domain. These extra cellular domains effectively associate with the same or similar molecules of surrounding endothelial cells specifically at the paracellular cleft while its cytoplasmic domain chemically links with interacting filaments of actin engaging proteins of related scaffolding. The catenins, p120 and plakoprotein form a molecular bridge which connects with the protein of tight junction- zona occludins and filaments of actin, thereby providing the necessary physical support. Other proteins molecules related to platelets and endothelial cells adhesion, like, CD99 and nectin are also concerned with adhesion junctions. Thus, the integrity of adhesion junctions plays an essential structural and functional role in the inter-endothelial interconnection [27, 28, 29, 93, 94, 95].
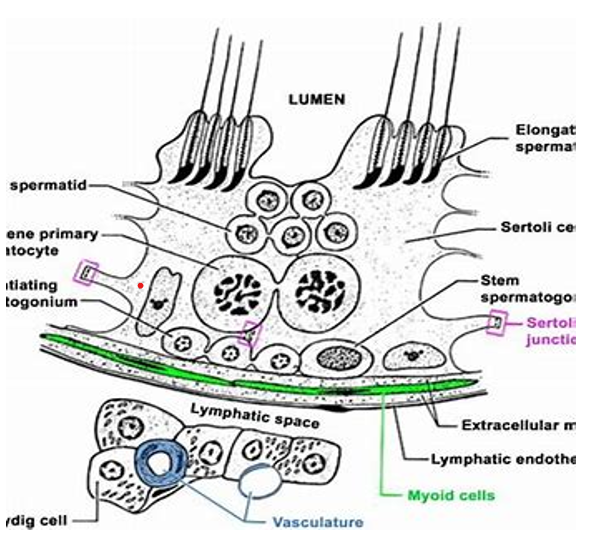
Testis Blood Barrier (Sertoli Cell Barrier)
Among vertebrate testis differentiation is an important process during the sexual reproductive cycle or otherwise. Seminiferous tubule has a somatic component called Sertoli cell and the germinal component germinal epithelium. Both work in physiologically coordinated manner during spermatogenesis and spermiogenesis resulting in the formation of haploid sperms. This physiologically synchronized coordination relates with germ cells and Sertoli cells ensuring qualitatively and quantitatively the physiological cellular efficacy of male gonad, the testis [96, 97, 98]. In general, the two adjacent Sertoli cells play structural and functional roles in the formation of ‘Sertoli-cell blood barrier’ (blood testis barrier) having developing stages of spermatogenesis and spermiogenesis. The Sertoli-cell blood barrier is closer to the basement membrane and isolates the premeiotic stages from post-meiotic stages of spermatogenesis [99, 100]. As the spermatogenesis progresses, various stages move from peripheral zone towards the central zone (luminal zone). Preleptotene spermatocytes (spermatid stage) transports itself through blood-testis barrier and reaches the apical (central) compartment. This stage undergoes spermiogenesis and relates with apical ‘ectoplasmic specialization’ involving testis-specific adherens junctions present along the interfaces between Sertoli cells and spermatid [99, 101]. This zone of interface showing cell-adhesion is under unexplored regulatory molecules. The coexistence of tight junctions, basal apical ectoplasmic specialization, a hybrid adherens junction are mostly made of proteins like cadherins and focal adhesion complex of mostly, integrins and laminins, gap junction (primarily connexin-43) and tight junctions constitute blood testis-barrier (Sertoli-Cell barrier) [99, 101]. This blood barrier regulates the transportation of metabolites between circulating blood in adjacent blood vessels and Sertoli cells present in seminiferous tubules containing developing stages of spermatogenesis. The primitive germinal epithelium and Sertoli cells line the seminiferous tubules and the related blood barrier is present between Sertoli cell of seminiferous tubule and supporting cells, also called sustentacular cells. This cytophysiological structure partitions the space of the seminiferous tubules as basal compartment and endoluminal compartments. The basal compartment is towards the periphery of seminiferous tubule while endoluminar compartment is present towards its center. The Sertoli-cell blood barrier is in biological and physiological communication with blood, lymph and endoluminar compartment that contains the developing stages of spermatogenesis. The testis blood barrier (Sertoli-Cell blood barrier or seminiferous blood barrier) structurally involves tight junctions, adherens junctions and gap junctions. These are located in between Sertoli cells and supporting cells (also called sustentacular cells) of the seminiferous tubules, and these can be investigated using immunofluorescence techniques [102, 103]. This blood barrier brings about specific functionality that ensures the maintenance of suitable physiological ambient environment in the seminiferous tubule and prevent the interference of physiological unwanted metabolites. This also ensures the quantitative and qualitative uptake of required metabolites and drugs, from the peripheral blood, lymph and the interstitial fluid surrounding stages of spermatogenesis. The germinal epithelial cells carry out a selective performance with respect to the recognition of self and non-self-entities and injected medical supplements in accordance with the specific body’s immune system. The germinal epithelial cells are not exposed directly to the peripheral blood but the molecular requirements are maintained [104, 105]. There are some interactions between Sertoli cells and the germ cells that represent the physiology of testis. There exists cytological correlation between Sertoli cells and the stages of spermatogenesis and role of blood- testis barrier present in the seminiferous tubule. These interactions involve cell-to-cell contact and paracrine factors [106]. The investigations using the electron microscopic technique (using lanthanum as tracer) reveals that the development of blood testis barrier relates with the sexual maturing stage of the vertebrate. In case of domestic fowl, the blood-testis barrier appears to be non-functional prior to puberty in spite of the presence of the structural and functional components like focal tight junction and desmosomes [107]. These researchers did not notice synchronized developmental correlation between the development of blood-testis barrier and the meiosis in the seminiferous tubules. Theoretically, the seminiferous tubule should be partitioned into peripheral basal chamber and central chamber endoluminar chamber resulting the separating the stages of spermatogenesis. No such arrangement is seen in the case of domestic fowl and early stages of spermatogenesis i.e., primary spermatocytes and spermatogonia are seen in unpartitioned chamber of seminiferous tubule. The effective blood-testis barrier was not seen as expected [107]. Figure 4 with reference to Sertoli cell barrier, in other vertebrates one finds variations in the complexes of junctional constitutions. Generally, among fishes and amphibian tight junctions and desmosomes are present very close to the Sertoli cells barrier. In these animals, this barrier develops after the completion of meiosis and the haploid spermatids. In case of zebrafish, an effective functionality of the said Sertoli cell barrier is not observed Gerhardt H, et al. [108], but in reptiles and aves, this barrier develops on the onset of meiosis and gets well-established before the onset of spermiogenesis [108, 109, 110]. This indicates that the Sertoli cell barrier develops during the process of spermatogenesis and it may not be observed during other phases of reproduction and non-reproductive phase. The prime function of tubulobulbular complex appears to remove the intercellular ectoplasmic specialized adhesion junctions and to provide specific shape to the head of spermatids and acrosome during spermiation. The exosomes communicate Sertoli cells and germ cells; this communication regulates spermatogenesis [101, 108, 109, 110].
Blood Oviduct Barrier
Oviduct is another reproductive component that plays major role in the process of reproduction. Its development, structure and role vary in different vertebrates. Because of its functional significance, its blood supply needs to be scrutinized. The blood flow in oviduct during its development and functional physiology play an important role to maintain its genesis and oviducal fluid; these are essential for the formation and maintenance of appropriate luminal fluid medium that helps to maintain suitable environment and embryo undergoing development and maturation within. In case of mammals, intense adrenergic innervation and higher numbers of β-adrenoreceptors (its presence is debatable) helps the controlled blood flow in oviduct. Acetylcholine imparts vasodilatory effects as it releases endothelium-derived relaxing factors, specifically in blood vessels. The roles of adrenergic and cholinergic innervation need further confirmation; even the role of neuropeptide-Y and vasointestinal peptide containing nerve fibres in relation to blood vessels need further confirmation although their presence is reported in mammals. The relaxing and contracting factors modulate the vascular tone of oviduct. Acetylcholine and histamine are prominent neurotransmitters which act on endothelial receptors and helps in the production of nitric oxide. Further, activation of guanylyl cyclase and after appropriate accumulation of cyclic GMP, these both effects cause relaxation of arteries of oviduct. In oviducal arteries the acetylcholine helps to release the endothelium-derived-hyperpolarizing factor and specific form of nitric oxide. This type of action is the result of opening of low-conductance Ca++, and activated K+ channels, thus, hyperpolarization and relaxation occur in oviduct. The oviducal vascular tone and potent and long-lasting contraction and relaxation are the impact of endothelium-derived contractile factor and endothelin. The high concentration of 5-hydroxytryptamine and histamine present in the mast cells present in oviduct and the histamine cause relaxation involving endothelium-dependent factors and activation of hydrogen receptors. The histamine plays a functional role during increase in the flow of blood and occurrence of edema, specifically, during ovulation and the antagonistic role of histamine and 5-hydroxytryptamine. This concept is debatable and needs confirmation. Other factors like products derivatives of relaxing and contracting cyclooxygenase, overall endocrine regulation, direct influence, modulation of neural and autocrine parameters also have their role during the functionality of oviduct [114]. The role of epithelial cells of the lumen as a ‘check post’ that helps to maintain optimum physiological milieu within oviduct; it prevents transport of unwanted and noxious metabolites and facilitate the entry of required metabolites; this concept is well established. This metabolic check is very essential for the processes like maturation of final gamete, fertilization, early embryonic development; but the physiological mechanism of these epithelial cells is still unclear [115, 116]. The epithelial cells function as barrier and prevent the transport of noxious molecules in addition to other regular functions. Mostly gastrointestinal epithelium has been the target of investigations. Their properties like permeability and permselectivity play significant roles during transport of molecules across their membrane. The physical parameters like pressure, electric current, and polyvalent cations, organic molecules-both channels blocking and nonspecific (detergent), properties of the membrane, cyclic nucleotides, agents that activate microfilament, and specifically H- and Na+ influence the transport functions of epithelium [117]. The luminar epithelium of oviduct actively particate in the transportation, maturation of gametes, fertilization, and early development of newly formed embryo within oviduct. This epithelium seems to play very active in maintaining the suitable physiological medium that facilitates the processes occurring in oviduct Schweigert FJ, et al. [115], Epithelia may be considered as an electrical circuit that includes batteries and resistance [117]. The transepithelial electrical resistance (TEER) acts as a significant qualitative functional parameter in oviduct. This parameter reflects on the qualitative efficacy of the oviducal luminar epithelium. Comparative study between transepithelial electrical resistance and the morphological aspects and apical- basal polarization (with respect to cellular height) and the degree of hormonal stimulation which is helpful in evaluating qualitative efficacy [116, 118]. Specifically, in case of epithelial cells, the intercellular communications involve cytological, biochemical and cytophysiological features in the form of intercellular junctions [6, 7, 117]. Tight junctions, gap junctions, and desmosome junctions are present in these cells. The density and location of the cell junctions, and the geometrics of a cell are of prime concerns for the cellular functionality and behavior under pathogenesis and normal physiological status. Gap junctions allow the movements of selected biomolecules through the tubular hexagonal array. Spot-desmosomes and belt-desmosomes are on the whole connecting cells and provide mechanical strength to the connecting cells. Tight junctions are the site of passive diffusion barriers [6, 7, 117, 119].
Blood Uterus Barrier
The uterus as a whole along with its extracellular fluid exhibit exclusive specificity that ensures the most suitable extracellular fluid physiological environment to the developing embryo within. The different rate and concentrations of various biomolecules in the fluids of uterus indicate the existence of some sort of barrier between the blood and the respective fluids of uterus. The extracellular uterine fluids like vascular, endometrial and luminal fluids, play very significant role during journey of embryo from its entry up to the developed stage till parturition. Any type of cytological, functional and molecular discrepancy in these fluids and uterus influence the implantation, viability of the embryo, baby and the mother [120]. The uterine microvascular endothelium and epithelium act as lipoid layers and their water filled channels function like an efficient barrier and exhibit different degree of restrictiveness. Even, there is a selective transfer of biomolecules and stromal and migratory cells from one fluid compartment into other compartment and also in endometrial fluid as observed while investigating the compositional values of these fluids and role of some sort of check or cytological barrier during the passage of the biomolecules. During embryonic development, the fluid luminal milieu changes and is maintained due to permeability of the membranes involved or presence of blood-uterine barrier along with cytological functionalities of endometrium and also of the embryo. This is so because of the presence of active dynamism of the cellular boundaries. Further, the cytological dynamic efficacy of the membranes, these are considered as ‘blood-uterine lumen barrier’ [120].
Blood Placenta Barrier
Tables 1-4 Placenta is a unique temporary organ formed in the uterus of female mammal as result of coitus/ sexual mating. There are morphological differences in the placenta of different groups of mammals like, canine, equine, bovine and humans [121]. Its basic histological aspects may show variations. It remains structurally and physiologically active till parturition, thereafter, it gets expelled from the system. It is the product of foetal and maternal tissues and remain separated but maintain very intimate contact at cellular and physiological levels. This intimacy is responsible for maintaining appropriate blood circulation between foetus and the mother. It is responsible for the exchange of respiratory gases, nutrients, along with waste metabolites from the foetus. It also protects the foetus from the infection Overall Representation of Types of Placentae Found in Veterbrates. (Amoroso, 1952) Basically, Placentae Are Classofied Based on the Degree of Intimacy Between Foetal and Maternal Tissues Types of Placentae A-Non-Deciduos or Semiplacenta: In This Type the Placental Tissue is Superficially Embeded in the Uterine Wall. It Is Present in those Vertebrates Where Blastocyct Is Located within the Cavity of Uterus and Forms Projections That Embed Uterine Wall Superfically uniting With it Loosely. These Projections are Celled Chorionic Villi. at Paerturation. These Villi are Withdrawn from the Point of Attachment to the Uterine Wall as a result there no Damage of Ghe Tissue Hence no Bleeding takes Place During Parturition.
A-Classification Based on Degree of Intimacy Between Foetal and Maternal Tissue
B-Placenta Vere or Deciduous Placenta (True Placenta): In this Type of Placenta Shows Highest Degree of Intimacy between Chorionic Villi (From Foetal Tissue) and Endometrium of Wall of Uterine (Maternal Tissue). During Parturition. The Mucosa of Uterine Wall Gets Eroded Causing Injury to the Point of Attachment, Hence Bleeding After Birth. Since this Placenta is Shed from the Ureter, this Condition Instructs Maximum Care of Mother After the Birth. It is Called Deciduous Placenta. This Placenta Shows Very High Degree of Efficacy, Activeness.
in case mother gets infected [122]. Among mammals, placenta is made of syncytiotrophoblast cells (maternal tissue) and capillaries from foetus. It permits the transfer of IgG immunoglobin only among other maternal immunoglobins. The neonatal Fc receptor (crystallizable fragment of IgG) plays significant role in this transfer. Binding between Fc receptor and maternal IgG and this complex reach the target through the endosomes of syncytiotrophoblasts. The appropriate foetal physiological pH ensures its release in the circulation of foetus [123].
Name of the Vertebrats in Which Present
This Type of Placenta Is Present in Vertebrates Like Pig, Cattle and Horse Etc.
This type of Placenta is Present in Primates, Dog, Cats, Rodents.
- C-Contra-Deciduate Placenta: In this type of Placentae
- Exhibit Loss of Both Maternal and Foetal Components of
- Placenta, Hence It is Called Contra-Deciduate Placenta.
Table 1: Overall Representation of Types of Placentae Found in Vertebrates.
| Placenta / Implantation | Types of Placentae Based on the Site of attachment in the Uterine Tissue to Ensure Attachment/Implantation of Embryo for Nutrition and Embryonic Development and to Ensure Successful Full Term of Pregnancy (Amoroso, 1952) | Examples |
|---|---|---|
| Central and Superficial Implantation | This type of Implantation is Placed Centrally and is Superficial attachment with Uterine Mucosa. It Keeps the Embryo Within the Lumen of Uterus. | Lower Vertebrates: Eutherians like Pigs, Cows, Rabbits, Sheep, Dogs and Cats |
Table 2: Types of Placentae Based on the Site of attachment in the Uterine Tissue to Ensure Attachment/Implantation of Embryo for
| Ecentric Implantation | In this type of Implantation the Early Blastocyst attaches it Self in an Eccentric Manner with in Uterine Epithelial folds. In Due Course of Development these Folds Completely Envelope the Developing Embryo | This type of Implantation/Placenta are Seen in Mouse, Rat, Beaver, Squirrel |
|---|---|---|
| Interstitial Implantation | In this type of Implantation, the Developing Embro Gets Embedded into the Uterine Mucosa Below Epithelium. The Uterine Endometrium Surrounds the Developing Embryo | This type of Placenta is Present in Vertebrates Like Hedgehog, Guinea-Pig, Some Bats, Humans and Chimpanzee |
Table 3: Types of Placentae Based on the Site of attachment in the Uterine Tissue to Ensure Attachment/Implantation of Embryo for
| Names of Placenta Types | Types of Placentae Based on the Distribution of Villi Distribution of Villi in Uterus is another Parameter Tp Classify Placentae (Amoroso, 1952) | Examples |
|---|---|---|
| Diffused Placenta | In This Type of Placenta Innumerable Villi Distribute Them Selves Uniformly Over the Complete Chorion | This type of Placenta is Present in Vertebrates Like Pigs, Horses, Mares and in Cetaceans (Marine Mammals Such as Whales, Porpoise and Dolphins |
| Cotyledonary Placenta | In type of Placenta, The Villi Aggregate in Specific Region (also Called Patch). This Placenta is in the form of Small Tuft, referred to as Cotyledon) While Rest of the Chorion does not Show any Villi and its Surface Remains Smooth | This type of Placenta is Seen in Ruminants (Cud Chewing Vertebrates), Ungulates Like Cattle, Sheep, Dears; In Camels and Giraffes the Placenta Exhibits Intermediate form where Villi are Distributed in a Scattered Manner and also Appears as Cotyledon |
| Zonary Placenta | In this type of Placenta, the Villi are Restricted to a Zone, Like Girdle Zone or an Annular Zone /Region of the Elliptical Chorion. This Distribution of Villi Covers the Girdle Zone Completely or Incompletely. This Morphological Form of Placenta is Seen Very Rarely. | This Morphological Placenta Is Seen as Incomplete Zone in Animals like Raccoon and as a Complete Zone in Cases of Animals Like Dog, Cat, Seal |
| Discoidal Placenta | In this type of Placenta, the Villi are arranged as a Compact Circular Disc or Platelike Structure on the Dorsal Region of Developing Blastocyst | This type of Placenta is Seen in Insectivores like Bats Rodents Like Mouse, Rats, Rabbit and Bear |
| Meta discoidal Placenta | In this type of Placenta, the Villi are Present in a Scattered Manner all Over the Surface of the Chorion and in Due Couse of Duration of Development arrange themselves in Either one or two Disc Like Structures, Hence, Referred to as Mon discoidal and Di-Discoidal Placentae | This type of Placentae is Seen in Primates; Mon discoidal Placenta is Seen in Humans and Di- Discoidal is Seen in Monkeys |
Table 4: Types of Placentae Based on the Distribution of Villi Distribution of Villi in Uterus is another Parameter Tp Classify P
| Names of Types of Placentae | Types of Placentae based on the Degree of Intimacy between the Uterine and Foetal Tissues; (Amoroso, 1952); Recap: The Uterine Blood Vessels and Foetal Blood Vessels Tissues Exhibit together Six Layers of Tissues: Namely I- Endothelium of Maternal Blood Vessel, Ii-Maternal Endometrial Connective Tissue, Iii-Uterine Epithelium, Iv-Chorionic Epithelium, V-Chorionic Connective Tissue, Vi-Chorionic Epithelium. These Histological Tissue Strata Ensure Selective Transportation of Metabolites to and Fro between the Tissues of Maternal Blood Vascular System and Foetal Blood Vascular System. Hence, are Regarded as Physiological Barrier between Maternal and Foetal Blood Vascular System | Examples |
|---|---|---|
| Epitheliochorial Placenta | In This type of Placenta the Epithelium of Chorion and Uterine Epithelium are in Contact with Each other, Hence, It is Referred to as Epitheliochorial Placenta | This type of Placenta is Present in Pig (Sow- Female Pig Used to Bread) Mare, Horse Cattle |
| Syndesmochorial Placenta | In this type of Placenta, the Chorion Comes Very Close to the Connective Tissue of Uterus Because the Quantity of Uterine Epithelium is Either Very Less or Becomes Absent, Hence, this type of Placenta is Called Syndesmochorion Placenta | This Type of Placenta is Present in Ruminant Ungulates like Cattle, Sheep, Deer, Giraff |
| Endotheliochorial Placenta | A Condition in Which the Uterine Mucosa is Reduced and the Epithelium of Chorion and Endothelial Layer of Maternal Uterine Blood Vessels Have Contact with Each Other. Thus, a Contact is Established between the Blood Stream in Maternal Uterine and the Foetal Blood Stream Invovling only 4 Histological Layers out of Six Layers. | This type of Placenta Is Present In Carnivores Such as Dogs, Cats, Bears |
| Haemochorial Placenta | In This type of Placenta there is A Direct Involvement of Epithelium of Chorionic Blood Vessels and that of Maternal Blood Stream Because the Endothelial Tissue of Maternal Blood Vessel is Absent. Thus, Only three Historical Layers Get Involved in this Haemochorial Placenta | This type of Placenta is Present in Primates, Insectivores (Moles, Shrews) and Chiroptera (Bats) |
| Haemoendothelial Placenta | In this type of Placenta there is a Maximum Reduction of Mesenchymal Tissue So that the Epithelium of Villi of Chorion And Endothelial Layer of Foetal Blood Vessel System Becomes in Direct Contact with Each Other. Thus, there Appears to be Only one Histological Layer between Maternal and the Foetal Blood System | This type of Placenta is Seen in Mouse, Rat, Rabbit, Guinea-Pig |
Table 5: Types of Placentae based on the Degree of Intimacy between the Uterine and Foetal Tissues; (Amoroso, 1952); Recap: The U
Table 4: Types of Placentae based on the Degree of Intimacy between the Uterine and Foetal Tissues; (Amoroso, 1952); Recap: The Uterine Blood Vessels and Foetal Blood Vessels Tissues Exhibit together Six Layers of Tissues: Namely I- Endothelium of Maternal Blood Vessel, Ii-Maternal Endometrial Connective Tissue, Iii-Uterine Epithelium, Iv-Chorionic Epithelium, V-Chorionic Connective Tissue, Vi-Chorionic Epithelium. These Histological Tissue Strata Ensure Selective Transportation of Metabolites to and Fro between the Tissues of Maternal Blood Vascular System and Foetal Blood Vascular System. Hence, are Regarded as Physiological Barrier between Maternal and Foetal Blood Vascular System.
Structural and Physiological Aspects of Placenta Blood Barrier
Placenta is basically a temporary structure formed during the onset of implantation during pregnancy and remains functional till the parturition. Placenta is embryonic structure to start with and becomes foetal functional organ. It is dedicated to maintain a suitable physiological environment for healthy development, growth of the embryo and foetus. It is the prime factor in maintaining full gestation period and a successful parturition. In addition to mammals, it is present in viviparous vertebrates including fish (genus: Poeciliopsis) [124], reptiles (placentotrophic species: Chalcides chalcides) [125]. Placenta is a complex, multilayered membranous and cytohistological entity dedicated to regulate the supply and eliminate the specific exogenous and endogenous metabolites between maternal and foetal blood circulatory systems. These membranes consist of syncytiotrophoblast, capillaries of foetal blood circulation, endothelium and interstitial layer. Placenta exhibits variations in its structure, location, in various mammals but all these modifications ensure the developmental and functional efficacies and successful termination of pregnancy in the form of parturition. Any physiological and functional aberrant fluctuation can result in complications during pregnancy and/or parturition. (for various modification in the placentae in various mammals refer table nos-1,2,3,4). In human being, complications like pre-eclampsia, intrauterine growth restrictions and some physiological complications may occur [126, 127]. Placenta histologically and physiologically plays an essential role in maintaining the most suitable intrauterine environment. The placenta ensures appropriate co-ordination of the physiological signals, mediation of exogenous and endogenous biomolecules and pharmaceutical products that facilitate the development, growth, nutrition and protection against infection and erroneous biochemical molecules.
The placenta distinguishes the maternal and the foetal blood vascular system cytologically its structural and functional aspects adapt in accordance to the progress of pregnancy. During first three months its syncytiotrophoblasts (also called Langerhans’ cells) along with mesenchymal tissue of the villi dominate as functional blood barrier. The mesenchymal tissue of villi has many ovoid ‘Hofbauer cells’ that exhibit macrophage behaviour; this mesenchymal tissue and the capillary walls of foetal blood system also facilitate the blood barrier function. The intact syncytio and cytotrophoblast layer/s and the mesenchymal tissue of the inner villi communicate with the microphagous cells and capillaries of foetus. As the 4th month progresses the cytotrophoblast disappears within the walls of villi and the thickness of the barrier tissue declines and the surface area elevates and attains say roughly 12 m2 area at the final stage of pregnancy. During 5th month the foetal blood capillaries become much more intense and also shift closer to the surface of villi. The capillaries move very close to the surface of villi in the mid-third term of the pregnancy because the cytotrophoblast layer gently die out and syncytiotrophoblast becomes very thin. The nuclei of the syncytiotrophoblast form a dense group and appear as proliferation knot while other region of syncytiotrophoblast do show the presence of nuclei and becomes adjacent to the capillaries. The syncytiotrophoblast layer slowly die out making it very thin. This zone acts as exchange zone. This nucleus free syncytiotrophoblast region merges with the basal membrane and the endothelium acts as placental blood barrier. Thus, the exchange zone is well established in 6th month of pregnancy [128].
Physiologically, the maternal-foetal interface-a histologically constituting placental blood barrier, becomes capable to gets signals from maternal as well as from foetal blood circulation [129]. The tissues of the barrier receive these signals and response accordingly. These signals relate to regulation of transfer of metabolites, synthesis of hormones, growth factors, and allocation of various biomolecules and signals. All these functions relate with development of foetus, maintenance of pregnancy and appropriate intrauterine environment, selective movements of metabolites, elimination of waste and prevention form the pathogens and toxins [130, 131]. Any disruption or dysfunction in placental tissue is likely lead to pathogenesis (intrauterine growth restriction, malnutrition, infection etc.). Further, on medication the pharmacological drugs have to reach the target for suitable remedy. The investigation on placenta provides relative better input in tissue engineering techniques, microphysiological studies, functioning of cells under developmental process, cytomacrophagy [131, 132, 133, 134, 135, 136, 137].
The physiological maternal health and medicophysiological status in all probabilities are bound to affect the health of the developing foetus. In addition to pathological status of mother, the nutritional metabolic status, during obesity and/or diabetic conditions affects the nutritional transport through placenta. Thereby, inducing the degree of growth of placenta and foetus [138]. In spite of the overall efficacy as selective nature of placenta as blood barrier, some deficient mother-to-foetal/child transmission may occur; few viruses like rubella virus, Zika virus and cytomegalovirus (CMV), find their way to the blood circulation of developing embryo via placenta [139, 140, 141]. The appropriate foetal physiological pH ensures its release in the circulation of foetus. Quantitatively, its transfer depends on the concentration in maternal fluid and this relates with the specific subclass. Thus, IgG1, IgG3, IgG4, and IgG2 show respective transfer efficacies in decreasing order [123]. The proteome in an organ has the ability to either synthesize the required proteins or procure them from the vascular circulation. The placenta along with the trophoblast have capability to synthesize the required biomolecules as per the functional needs. Generally, a placenta has approximately 350 genes which are functional during pregnancy; these express proteins like developmental retrotransposon-derived protein; in cytotrophoblast, gene for cancer testis antigen, prostate associated gene that protects cell against stress involving increased concentration of 21p and supressing ROS formation. In syncytiotrophoblasts genes for hCG1 (human chorionic somato-mammograph/human chorionic somatotropin/ human placental lactogen kiss peptins like kiss peptin 54 (metastin) for G-protein coupled receptor (GPR54) (KISS 1) are functional. Genes for (PAPPA2: Pregnancy-associated plasma proteinA2) and (PRG-
2: Proteoglycan-2/Eosinophil major basic protein) are functional in extraviollous trophoblast [142].
The process of DNA methylation seams to play effective role in the physiology of an individual. The methylation also influences the functioning of DNA and it does not change its sequence. When methyl group is attached to gene promotor the resultant methylated DNA inhibits the degree of gene transcription. DNA methylation has a significant role in the embryogenesis, developmental process and survival of embryo in humans. Basically, the process of methylation plays an effective role during genomic imprinting, inactivation of X-chromosome; even process like aging, carcinogenesis and functioning of transposable elements [143].
Hepatic Blood Barrier: (Blood Bile Barrier)
Primarily, liver is a prime site of detoxification, carbohydrate, protein and lipid metabolisms, and other metabolic activities. This demands a perfect suitable physiological environment to maintain physiologically healthy liver. The hepatic blood barrier, mostly referred to as blood-bile-barrier, plays most role in accomplishing the biochemical, biophysical physiological hepatic functionality. This cytohistological structure protects the liver tissue i.e., hepatocytes from the toxic impacts of bile and establishes histological demarcation between blood vascular tissue and the bile canaliculi system within liver. For healthy liver, a perfect and selective supply and transfer of metabolites, pharmaceutical drugs are required and blood-bile-barrier plays significant role in this aspect [63]. This barrier is cytohistologically consists of tight junctions, adherens junctions, desmosomes, gap-junctions, signalling pathways and transporters related to hepatocytes and bile. All these components of blood bile barrier ensure the physiologically appropriate hepatic functions and help to keeping unwanted and toxic metabolites at bay. This barrier also corelates with hepatic pathogenic conditions like cholestasis and other hepatochronic conditions or hepatic dysfunctions and pathogenic conditions [64]. The structural and functional cellular unit is hepatocytes. Fully developed hepatocyte has three recognizable membranes namely, sinusoidal or basal, lateral and canalicular or apical membranes. Sinusoidal/basal membrane faces hepatic sinusoids. These sinusoids (passages) have a lining of lose hepatic sinusoidal endothelium and direct blood to central vein from hepatic artery and portal vein. The canalicular membrane constitutes bile canalicular tubular- mesh and carry bile acids from hepatocytes to intrahepatic bile duct via portal triads. The second membrane i.e., lateral membrane is placed in between the bile canaliculi and apical/ canalicular membrane; it is fused and coexists forming tight junctions also called ‘zonula occludens’. This arrangement separates the canicular/apical membrane and sinusoidal/ basal membrane and constitutes ‘blood-bile-barrier.’ The intercellular junctions affix the hepatocytes with each other and form liver cell plates. These intercellular junctions involve gap, tight and adherens junctions along with desmosomes. These aggregate densely in the region of canaliculi. The tight junctions between hepatocytes and surrounding the bile canaliculi pin the paracellular spaces present in between hepatocytes and also restore polarity of hepatocytes. In addition to tight junctions, adherens junctions, desmosomes, the gap-junctions are also present and facilitate the working of blood-bile-barrier. Because of the polarity of hepatocytes and the specific transporters present on them, all these regulate the collection of bile inside bile canaliculi and help the working of tight junctions. The cholangiocytes are the cuboidal epithelial cells and are present in the region of interlobular bile ducts. These get modified into columnar cells and secrete carbonate in the region of larger bile ducts, extrahepatic ducts and porta hepatica. Primarily, these cells transport bile acids and facilitate the functioning of hepatic cells [144]. These also have tight junctions and play functional role as barrier promoting transfer of bile from liver to extrahepatic biliary network [64, 145, 146].
Renal Blood Barrier
Renal blood barrier is technically known as glomerular filtration barrier (GFB). Glomerulus/nephron is the structural and functional unit of a kidney and it is the site of filtration of blood and formation of urine. Glomerulus is differentiated in to double walled Bowman’s capsule, neck, renal tubule and collecting tubule. A knot (dense bunch of capillaries) called glomerulus is enclosed in a double walled Bowman’s capsule while the renal tubule is enwrapped by copious blood capillaries. Both sets of capillaries are very intimate with the nephron. The trilaminar aspect of capillaries consisting of glycocalyx rich layer, fenestrated endothelial cellular layer and podocytes along with a membranous layer between the interdigital foot of podocytes. This cytological aggregation constitutes ‘glomerular filtrate barrier trilaminar’. The glomerular Bowman’s capsule performs ultrafiltration of blood. The glomerular filtrate blood barrier plays a significant role during the selective transport of metabolites. This selective metabolic transport is effectively applicable to drug screening, elimination of molecules used during imaging and research. The related studies are useful in the search of repertoire of mechanism processes like signalling in cell physiology [14]. This renal blood barrier/glomerular blood barrier offers an efficient cytohistological interface dedicated to specific selective molecular transport based on their charge and size. The milieu and the dynamics of blood flow through nephron have significant impacts on the activity and efficacy of the interface formed between glomerular endothelium, basement membrane of glomerulus and podocytes. This is also responsible for its structural and physiological functional integrity [14, 147, 148, 149]. Any infection, pathogenic conditions, dysfunctions and damage lead to the deficient functioning of glomerular blood barrier [149].
Structural and Functional Aspects
Glomerular blood barrier exhibits a complex structural interconnectivity and an active interplay of the constituting components. This barrier is a trilaminar composition made of well-formed glycocalyx layer, fenestrated cellular endothelial layer and podocytes with their membranes sandwiched between interdigital podocyte feet. Glomerular endothelial cells cover the luminal interface surface of blood capillaries of glomerulus. The podocytes encase externally the blood vessel extending their primary and secondary feet. This association forms an interdigital motif. Permselectivity i.e., qualitative and quantitative separation of ions is one of the basic functions of glomerulus. The slit diaphragm present between the neighbouring foot process of podocytes also accomplishes this function. Glomerular filtrate blood barrier exhibits fenestrations in the basement membrane. The glycoproteins, proteoglycans and glycosaminoglycans of glycocalyx make a distinct lining in this region. It also measures and maintains charged ions, thus, effectively facilitate the process of permselectivity in glomerulus. The foot processes of podocytes are present on the opposite side of the basement membrane. These feet processes are in communication with the interlink proteins like podocin and nephrin, of slit diaphragm. These proteins help to regulate albumin in glomerular blood filtrate barrier. Blood flowing through this cytohistological setup undergoes selective filtration due to the efficient glomerular filtration barrier [14]. Podocytes and its cytoskeleton are responsible to make the architectural basic frame work and provide mechanical stability to glomerular filtration. The slit diaphragm is the result of interdigitated specific projections and these contribute to the overall physical stability of glomerular filtration barrier [150]. Podocytes play a significant functional role in establishing intercellular exchange of information between podocytes and other cellular components of glomerular filtration barrier. The studies based on genetics and genomics are helpful in elaborating the therapies related to glomerular filtration barrier. The mechanism related to inhibitor of apolipoprotein L-I, short transient receptor; potential channels, soluble free-forms like tyrokinase-I (also called soluble vascular endothelial growth factor receptor-I), roundabout homologue-2, endothelin receptor-A, soluble urokinase plasminogen activator surface receptor and substrate intermediate coenzyme Q10/CoQ10. These molecules reflect on the participation of mitochondria and mechanism of contractile actin-myosin during the physiology of glomerular filtration barrier [150].
Two of the prime parameters namely, renal blood flow and glomerular filtration contribute to renal physiology. A well organized balanced interplay of renin-angiotensin- aldosterone system is another parameter that is essential for renal physiology. The changes in renal arterial pressure (decline) and elevated renal sympathetic activation due to beta- adrenergic receptors or reduced sodium supply to the cells of maculadensa influence the production of renin from juxtaglomerular cells. The interaction between renin and angiotensin (a product of liver) results in the formation of angiotensin-1. Angiotensin-1 involving angiotensin converting enzyme (product of lungs) forms angiotensin- II. This angiotensin is responsible for induction of vasoconstriction and elevation of the pressure. When renal blood flow is weak or low angiotensin-II constricts the blood efferent arterioles and ensure appropriate rate of filtration. The angiotensin-II formed also activates the expression for aldosterone with in adrenal cortex, this results in increased sodium channel insertion, elevated activity of sodium/ potassium pump and the enhanced cellular potassium and hydrogen excretion. All these interactions together create an elevated gradient and affect the sodium and water reabsorption. Angiotensin-II also escalates the expression of antidiuretic hormone in the posterior pituitary gland. This hormone helps to bring about water adsorption by positioning aquaporin channels in the apical interface of membrane of the principal cells. This hormone also activates hypothalamus resulting in enhanced thirst. It also influences the mechanism for dehydration or mechanism to activate signalling for the state of low volume of water in body [151]. Under normal physiological functioning of glomerular filtration barrier maintains free permeability of water and molecule with small and midsized solutes of blood plasma. Any form of dysfunction or disruption of interactivity in the trilaminar constitutional or pathogenic status or defective inherited or acquired parameter results in defective performance of the glomerular filtration barrier [14, 152, 153].
Molecular Aspects of Glomerular Filtration Barrier
The glomerular filtration barrier needs specific structural, mechanical and molecular environment to carry out its task. The coherence of cytoskeleton of podocytes plays significant role in this regard. The slit-diaphragm arrangement is established due to the integrated nature of specialized projections of the podocytes and these contribute to the functional efficacy of glomerular filtration barrier physically and physiologically. Nephrin related proteins family is essential for the structural associated integrity of slit-diaphragm and the cytoplasmic tails are important for the signalling process. Additionally, the nephrin protein family members form the molecular tight, adhesion, gap and neuronal junctions present in slit-diaphragm; these are responsible for the specific cell-to-cell connections Dalal R, et al. [154] and gene NPHS-I encodes nephrin and gene NPHS-II encodes podocin. These genes express exclusively at the site of slit-diaphragm. The proteins nephrin and podocin fasten and secure the cytoskeleton with plasma membrane. These proteins establish a bipartite scaffold among the interlace formed by the projections of podocytes and it may be regarded as tertiary foot processes. There is a spring-like-protein-bridge formed and this regulates the transfer of macromolecules. These proteins and the related molecular constituents establish the podocyte tertiary foot processes. The proteins encoded with CD2AP gene and proteins associated with attaching the podocytes to the basement membrane like α3β2 integrin and those constitute slit-diaphragm maintain glomerular filtration barrier. The dynamic control of actin cytoskeleton associates with the focal segmental glomerulosclerosis and nephrotic syndrome [150].
Pulmonary Blood Barrier/Pulmonary Blood Gas Barrier
The pulmonary blood gas barrier is very specific example of bioengineering where diffusion, exhibition of cellular functional applications of strength and stress principles can be seen. A constant rhythmical ventilation and self- maintenance of tissue are in action under varied conditions. Its study is essential to understand the pulmonary physiology under normal and pathological conditions. Its successful regeneration, recapitulation and controlling efficacy of the pulmonary blood barrier during pathophysiological conditions can be observed simultaneously. Related investigations may be helpful in evaluating the derogative impacts of environmental stress, impact and fluctuations on the pulmonary blood gas barrier in human being and test animals in space, personnel working at high altitudes and deep-sea under-water assignments during ocean and oil explorations. The probable parameters involve during the functionality of pulmonary gas blood barrier or alveolar blood barrier, it isolates gas from blood from the enormous surface area of alveoli and capillaries. A fine thinness of wall of alveoli permits gaseous diffusion, robustness to withhold transmural pressure (inside pressure relative to outside pressure) and repeated dynamic transpulmonary impact, perpetuate physiological active air-liquid interface and facilitate effective ventilation and unobstructed alveolar functions [155, 156].
Structural and Functional Aspects
The alveolar capillary barrier/pulmonary blood-gas barrier is present in the specific zone of lungs where exchange of gases can occur effectively. Generally, this barrier consists of type-I pneumocytes present in the alveolar wall and cells of capillary endothelium. The basement membrane is sandwiched between pneumocytes and cells of endothelium. Basically alveolar-capillary barrier concerns with the controlled gaseous exchange and prevents the formation of air bubble in the blood and the blood that enters the alveoli. This barrier permits the transportation of molecular carbon- di-oxide, oxygen and carbon-mono-oxide. The thinness of the blood-gas barrier varies between 600nm to 2µm and at some zones it about 200nm. This feature facilitates sufficient diffusion of oxygen. In spite of fine thinness of the membrane of this barrier it is very tough and strong. The type-IV collagen is present between the epithelial and endothelial cells and this collagen is the cause of the toughness of the membrane of this barrier. The pressure difference around 40 mm of mercury causes ruptures of the barrier membrane. The major five types of cells are present in the pulmonary parenchyma. Alveolar-I type epithelial cells (around 8%,), alveolar-II types cells (around 7%), type-I-pneumocytes (around 16%), cells of endothelium of capillary (around 49%), cells present in the interstitial space and alveolar macrophages (around 19%). The proportion of alveolar cells and the percentage of other cells of blood alveolar blood barrier varies in different species. The numbers of alveolar macrophages varying between smokers and no-smokers (male and female both) [156].
Gaseous exchange, thermal and immunological responses are the prime functions of lungs. These responses are the result of intense interactions between the functional epithelium, intact alveolar capillaries membrane and viable endothelium. Pulmonary epithelium is the primary target and vulnerable to the personal human habits, pathological, congenital and acquired diseases, and environmental insults. During these insults and other physical and physiological impacts this alveolar component exhibits processes like repair, remodelling and regeneration/reorganization. When the derogative impacts become sever, these processes take a back seat and as a result dysfunctioning of epithelio- vasculo-mesenchymal tissue occurs [157]. These conditions call for some technologies and efforts for restoration of the structural and functional normalization suggested the technique like de-epithelization in the said tissue is suitable for such investigations [157].
Skin Blood Barrier
Skin is a multifaceted organ and it is protective, limiting membrane of a biosystem. It is responsible for the formation of epidermal derivatives like scale, hair, fir, feather, and exoskeleton among crustaceans. This cellular multilayered seemingly simple is very complex and this complexity renders it the most useful components of a biosystem. A healthy skin is an effective barrier and prevents skin form mechanical, chemical, thermal and site for microbial damages or environmental insults. It consists of three cellular layers namely epidermis, dermis and subcutis. Among animals it is made of epithelial cells arranged is specific layers. Cells of this layer are interconnected through cellular junctions and exhibit very strong cohesion. This barrier has outermost layer of keratinocytes (horny/cornified dead layer-stratum corneum, product of keratinization). Just inner to this layer there is a living cellular resilient layer that is responsible for mechanical cohesion among the cells involving desmosomes, adherens and tight junctions [158]. The cells of epidermis have corneocytes (epidermal cells) arranged in a brick-mortar manner. Each corneocyte is brick shaped and embedded in epidermal lipid matrix communicating and adhering with neighbouring cells via corneodesmosomes; their cytoplasm has many natural moisturizing factors. This lipid is in the form of multilayered lamellar sheets rich in ceramides, cholesterol, and free fatty acid [159]. This cytological arrangement controls molecular diffusion (trans-epidermal water loss/diffusion) form interior to outside as well as chemicals from outside into inside. This diffusion is intracellular involving epidermal lipid matrix and also functionally transcellular via corneocytes. The parameters such as, tight junctions, the proteins constituting scaffolding, layered brick-mortar arrangement within lipid matrix, the interactions between proteins of keratinocytes, the distance/thickness of a layer of corneocytes, size and cohesion between these corneocytes contribute to the rate of diffusion through this part of the skin. The filaggrin-rich keratohyalin derivatives along with filaggrin and lamellar granules are present in keratinocytes. These also have lipids, proteases, protease-inhibitors, antimicrobial peptides and corneodesmosin-a structural protein. Granular keratinocytes have lamellar granules which discharge their contents into the extracellular space present below. This whole process is helpful to maintain homeostasis and barrier ability of the skin [158, 160]. The structure and mechanical integrity of stratum corneum is due to filaggrin which is a specific structural protein itself and its aggregated form. This molecular congregation is responsible for the structural and mechanical property of stratum corneum [161]. The epidermal proteins like filaggrins and its derivatives present in the keratohyalin granules of keratocytes of stratum granulosum are the primary constitutional components and responsible for epidermis as a multifunctional entity. Dephosphorylation, enzymatic deimination of cornified keratinocytes and pro- filaggrins play major roles in dispersing filaggrin within corneocytes and form crosslinked cover over them. The free amino acids from the filaggrin are responsible for the natural moisturising factor related to stratum corneum. The acidification of liquid-liquid phase of cell contents of these cells has modified physiochemical features that facilitates the incrimination of dispersed proteins at the interface and help their condensation after its cornification [161, 162, 163].
The interconnections between intercellular junctions with the cytoskeleton is the key functional barrier component of skin blood barrier. A well-established transcellular network is essential and intercellular units like adherens junctions, are very helpful in this regard. These intercellular units associate with cell membrane and cytoskeleton. Cadherin/cadherins-complex–a component of adherens junctions, also involves in the formation of tight junctions and desmosomes. Occludins, claudins and E-cadherin are major transmembrane proteins and these also form tight junctions. These proteins link with zonula occludins and are present on the inner side of cell membrane towards the cytoplasm connecting with cytoskeleton via actin filaments. Tight junctions are also a component of the barrier (external) and control the to-and-fro movements of water molecules. Exchange of ions, metabolites, and secondary messengers move in and out of the epidermis through gap junctions forming channels that regulate cytosols. Connexins are the constitutional components and participate in proliferation of keratinocytes. These are essential for wound healing, psoriasis and dermal cancer. Gap junctions participate in coordinating cellular differentiation and ensure appropriate coupling between electrical and metabolic constituents. All these are also important components of skin barrier. Desmosomes are present in the stratum corneum but in the form of corneodesmosomes. Desmosomal cadherins, desmogleins 1-4, desmocollins 1-3 together form desmosomes. The desmosomal plaques-a product of link between desmosomal cadherins and molecular network, connects keratin filaments [158, 164, 165, 166].
The appendages derived from epidermis also facilitate the efficacy of skin as barrier, namely, nails, hair, sebaceous glands and sweat glands. The proximal nail fold and nail plate acts as a barrier to water molecules, chemical and microbes. Hair help to maintain body temperature, to detect warning signals for the scalp and safe-guards. It provides protection from minor skin trauma; density of hair also protects epiderma against solar exposure and production of sebum. Sebaceous glands regulate water loss, temperature and act as an antimicrobial agent. Sweat glands maintain body temperature and dermal pH (normal rage 4.5 to 5.5); dermal acidity is essential for dermal permeability. The dermis/ subcutis consists of predominantly of adipocytes and is helpful in thermoregulation, to reduce impact of external dermal mechanical shocks and protection. Connexins and its family members help in epidermal hemostasis and epidermal barrier structurally and functionally. These proteins and their defective expression and their functioning are the causes of pathological conditions like ectodermal dysplasia, eczematous dermatitis, non-syndromic hearing loss, and wound healing related abnormalities [167].
Biochemical and physiological environment of skin blood barrier. The skin has acidic environment pH (range The acidic 4.5-5.5). The dermal acidity is essential for its permeability nature. The enzymes like β-glucocerebrosidase and acid sphingomyelinase are present in stratum corneum and exhibit optimum respective enzyme activities. These enzymes help in the formation of ceramides and free fatty acids. These molecules result in the suitable diffusion via epidermis and its functional maintenance. The acidic pH controls the efficacy of kallikrein-5 and kalloikerin-7; these bring about cleavage activity of corneodesmosomal proteins. These proteins are responsible for the adhesion amongst corneocytes. The dermal acidic environment is very effective in preventing colonization of pathogens and provides support to the normal dermal microbiome [158].
Blood-Ocular Barrier
This blood barrier controls the movements of unwanted metabolites from the systemic blood to the parts of an eye and provides the suitable physiological ambient environment. Conceptually, this is a cytophysiological barrier that regulates to-and-fro molecular transfer to the parts of eye and the blood flowing in the local blood capillaries. The vitreous (clear transparent fluid), lens and ciliary body, along with its associated components divides the interior of bye- ball into smaller anterior and bigger posterior chambers. Anterior chamber is filled with aqueous humour which is a clear, transparent and relatively thinner fluid while posterior chamber is filled with vitreous humour, transparent, clear and relatively thicker fluid. Chemically, it is 99% water and remaining 1% is collagen and hyaluronic acid. The blood ocular barrier (BOB) system consists of two sub-systems namely blood aqueous ocular barrier and retino-blood barrier/blood retinal barrier [168]. Cytohistologically, endothelium of blood capillaries of retina, iris, ciliary body and retinal pigment of epithelium constitute blood ocular barrier. This barrier regulates the transfer of metabolites and pharmaceutics between blood, the interior fluids the parts of an eye. The functioning of this barrier gets affected during its inflammation. The cells of non-pigmented ciliary epithelium and endothelium of the blood capillaries in the vicinity are the structural and functional parts of the ‘blood ocular barrier’. The tight junctions present between non-pigmented ciliary epithelial cells play major role in formation of this epithelial barrier. The proteins constituting tight junctions present between vascular endothelial cells are similar to those present in epithelial tight junctions; such junctions are seen in iris also. The endothelial barriers are present in vascular capillaries of iris [168]. Blood ocular barrier is responsible for maintaining suitable nutritional state, functionality of cornea and the lens. Dysfunctioning of this barrier results in conditions like trauma, ocular inflammation, vascular diseases and ocular surgery. Leakage of unwanted proteins (plasma proteins) due to the defective blood ocular barrier results in ‘cloudy conditions’ in anterior as well as posterior chambers. The presence of fibrinogen and other proteins in these components causes plasmoid condition. Any structural damage to the components of blood ocular barrier also induces inflammation in the nearby cells [168]. Blood retinal barrier system: Biophysically, blood interacts with retina at two interfaces; interface between retinal blood capillaries and interface between choroid-retinal pigment epithelium. These two interfaces are involved cytophysiologically with blood retinal barrier. There are tight junctions between the endothelial cell of retinal blood capillaries; this set of tight junctions may be considered as ‘inner blood retinal barrier’. The other set of tight junctions is located between the cells of retinal pigment epithelium and this may be considered as ‘outer blood retinal barrier’. The molecules having molecular mass between 20 to 30 kDa do not move through the inner blood retinal barrier. The molecules with lesser molecular mass such as glucose, ascorbate move across this barrier due to ‘facilitated diffusion’ i.e., ‘active transport’. This mode of molecular transport also helps the movements of organic anions, fluorescein and prostaglandins. The outer blood retinal barrier i.e., the cells of the pigmented epithelium, are interconnected with zonula occludens (a complex made of tight intercellular junctions) that permits to-and-fro movements (bidirectional moments) of variety of molecules including glucose. Mass flow of fluid towards the choriocapillaris occurs and involves functional mechanisms of Na+/K+ATPase pump. This transport is passive transport and is responsible for the transfer of amino acids and glucose as non-ionic solutes. The directions of this movement are from vitreous towards blood depending on the concentration gradient. Its efficacy is restricted and involve the movements of sodium, potassium, phosphate and glucose [168, 169]. Blood retinal barrier is another component of blood ocular barrier. The endothelial cells (single layer) of non-fenestrated blood capillaries of retina and pigmented cells of retinal pigment cell (epithelial cells) together form ‘blood retinal barrier’. The tight junctions of these cells check the movement of larger molecules from capillaries of choroid and retina [170, 171, 172].
Blood Thymus Carrier
Thymus is a lymphoid bilobed gland functionally related to the immune system. It is situated in thorax anteriorly to superior mediastinum behind/dorsally to sternum and in front of heart. Basically, it is the site of formation and maturation of thymus lymphocytes (thymocytes) or commonly called T-cell. There is a lining of epithelial cells, these participate in the maturation of thymocytes and are considered under adaptive immune system.
The thymus gland is covered by a septae. Each thymic lobe is differentiated in to central zone called thymus medulla and peripheral zone called thymus cortex. The subcapsular cells are squamous epithelial layer and function as blood thymus barrier. The epithelial cells of middle layer of cortical region are called thymus stellate and are made of cyto- reticulate cells. The cells of inner most layer of cortical region is squamous in nature and act as corticomedullary barrier. The thymic medullary zone differentiates in to squamous thymic epithelial and cyto-reticulate layer. The blood thymus barrier consists of epithelial cells and are placed very close to the vicinity of capillaries wall. These cells have dense intercellular junctions, transporters and less numbers of pinocytotic vesicles. These regulate and select molecular movements across. The numerous cytoplasmic processes arising from the cells of endothelial cells and the adventitial cell extend towards the lumen of outside capillaries which also facilitate the selective molecular transport. This cytological set up can be readily seen using intravenous thorium dioxide movements in traces. This barrier also distinguishes the T-cells from the other blood cells flowing in the vicinity of least fenestrated capillaries. The blood thymus barrier permits restricted entry of circulating antigens to the developing cortical lymphocytes. This barrier allows limited movement of macromolecules and circulating antigens; in the juxtamedullary region, cortical venules and in medulla stops the circulating antigens from blood stream into cortex of the thymus where development of T-cells occurs [173]. The selective and controlled distribution of macromolecules in the medulla of thymus from the circulating blood in the vicinity, depends on degree of differential permeability of the vascular membrane forming blood capillaries in the region, strategic placement of macrophages in the parenchyma of blood capillaries and spatial arrangement of the histological components of medulla. The endothelial junctions present in the blood capillaries in cortex are completely impermeable. Only slight amounts of tracers move across plasmalemmal vesicle via capillary endothelium and the macrophages present in stretched manner in the cortical capillaries [174, 175]. The self-antigens present on the dendritic cells, and other molecules present in the blood stream of the vicinity of thymus medulla get transferred. This molecular transfer induces tolerance and maintain uninterrupted immigration of T-cells precursor into medulla. The cortico-medullary junction helps to withdraw the mature thymocytes in thymus medulla. There is an optimum expression for proteins specifically claudin-5 (Cld-5) in all vascular epithelial cells present in thymus cortex. The protein, claudin-5 is present in all tight junctions present in thymus cortex. But the expression of claudin-5 declines in the epithelial cells present in thymus medulla and corticomedullary junctions. Intravenous administration of biotin tracer did not show access successfully in the blood capillaries present in the cortical region while the traces of biotin reach parenchymatous tissue of medulla through blood capillaries having tight junction having claudin-5. This indicates the declined degree of expression of protein claudin-5 in these cells. In all probability, a distinct profile of expression of claudin-5 plays an important role in regulating the selective transfer of molecules, and T-cells and it acts as gate-way between thymus cortex and thymus medulla [176].
The size and activity of thymus reduces as the individual grows; it attains maximum size and activity during the neonatal and preadolescent; in later stages of the individual. It declines gradually and adipose tissue takes over the functional tissue. This does not stop the formation of T-cells even in adult stage. The epithelial cells together form Hassall corpuscles. The thymus nerve cells help in functional maturity of thymocytes (T-lymphocytes). Thus, in this region the thymocytes becomes positively and negatively responsive. When these matured thymocytes (T-cells) interact with the receptors of major histocompatibility complex (MHC), these thymocytes are classified as positively selected T-cells and their response is positive selection. Those matured T-cells which do not interact with the other proteins of the body are classified as negative selected T-cells and their behavior is negative selection [173].
Blood Pancreatic Barrier
Pancreases is one of the digestive glands associated anatomically with intestine within the close proximity of stomach. Pancreas gland has exocrine as well as endocrine components, acinar glands and Islets of Langerhans respectively. Both of these components have modified epithethilial cells arranged in a specific group. In case of acinar gland these cells form a hollow alveoli like structure enclosing a lumen that leads to the pancreatic duct while in case of Islet of Langerhans the modified cells are grouped as one rounded spherical mass within the pancreatic tissue. In both cases there is a distinct base membrane to which the peripheral cells are in communication. The β-cells of Islets of Langerhans are enclosed by either single or double basement membranes with very thin or no endothelial tissues. Generally, the basement membrane is effective, protective and functionally active. The basement membrane establishes an organized intercellular contact between the respective cells and the components of surrounding tissue. This intercellular link provides a physical, biochemical, biophysical and molecular contact that brings a physiological molecular functionality with the ambient tissue organization and the microenvironment. The laminins are the prime functional component present in basement membrane. The isolaminin-511 (formerly known lamini-10) is present as functional component of basement membrane that encloses human endocrine islet. This membrane exhibits powerful and polarized expression of a protein that is called Lutheran glycoprotein which is prime receptor that interacts with laminin α5 chain. The human endocrine basement membrane has specific receptor -Lutheran glycoprotein, on the basal membrane around human islets facing it [177]. Pancreas being exocrine as well as endocrine gland has a complex functional mode and is harmonized involving integrated responses because of functionality of nervous circuit. This nervous circuit includes sensory fibres form sympathetic and parasympathetic nervous systems and vagus nerve. Other systems like reticular activating system, mesencephalic centres, hypothalamic autonomic nervous system, thalamocortical circuits and interoceptive centres are the prime regulatory agents that synchronize pancreatic functionalities [177, 178, 179, 180, 181, 182].
Conclusion
The composition of a blood barrier includes selective semi-permeable membrane, endothelium, pericytes, capillary basement membrane, astrocytes, tight junctions and enzyme or metabolic barriers. These components are suitably architectured involving the interstitium of the concerned organ. Fine blood vessels from peripheral circulation in a manner that maintain fine difference between the supply of peripheral blood and the blood present in the related organ. Each prime organ requires essential micro and ambient environment to maintain its specific assigned physiological function and blood barrier has its exclusive role accordingly. These specifications depend on the homeostatic, biochemical and neuronal-physiological conditions of the organ. The blood barrier helps to maintain such conditions. The rate of blood flow, degree of vessel permeability, thickness of wall of capillaries, physicochemical properties of the biomolecules, drugs, unwanted molecules and cellular blood components influence the physiological environment. Each and every organ is physiologically efficient to carry-out its assigned function/s with the help of its neural, hormonal coordination and specific blood barrier. Techniques like immunohistochemical techniques, cell culture, electron microscopy, use of tracers, proteomic are suitable to investigate blood barrier functioning. Basically, each biological cell has ability to tolerate and struggle to survive the physiological and environmental insults. Further investigations will illustrate more on the theoretical and applied aspects of blood organ barrier in the field of pathological conditions and drug designing for remedial aspects.
References
-
Bouffard G (1906) Injection des couleurs de benzidine aux animaux normaux. Annals of Institute Pasteur 20: 539-546.
-
Setchell BP (2008) Blood-Testis Barrier, Junctional and Transport Proteins and Spermatogenesis. Molecular Mechanisms in Spermatogenesis pp: 212-233.
-
Goldmann EE (1909) Die a¨ußere und Innere Sekretion des Gesunden und Kranken Organismus im Lichte der “vitalen Farbung”. Beitr Klin Chir 64: 192-265
-
Easton AS (2014) Regulation of Permeability Across the Blood-Brain Barrier. Biology and Regulation of Blood- Tissue Barriers pp: 1-19.
-
Abbott NJ, Ronnback L, Hansson E (2006) Astrocytes- Endothelial Interactions at the Blood Brain Barrier. Nat Rev Neurosci 7(1): 41-53.
-
Lahir YK (2014) Morphological Aspects of Intercellular Communication. Bionano Frontier 7(1): 3-11.
-
Lahir YK (2015) A Dynamic Component of Tissues-Extra Cellular Matrix: Structural, Functional and Adaptive Approach. Biochem Cell Arch 15(2): 331-347.
-
Lahir YK (2022) Biological Cells Exhibit Intellectual Behavior. World Journal of Pharmaceutical Research 11(4): 2194-2228.
-
Yang BA, Westerhof TM, Sanon K, Merajaver SD, Aguilar CA (2020) Engineered Tools to Study Intercellular Communication. Adv Sci (Weinh) 8(3): 2002825.
-
Qiu Y, Myers DR, Lam WA (2019) The Biophysics and Mechanics of Blood from a Materials Perspective. Nat Rev Mater 4(5): 294-311.
-
Lahir, Harivansh Y (2016) Understanding Basic Role of Glycocalyx during Cancer. Journal Radiation and Cancer Research 7(3): 79-84.
-
Lahir YK (2023) Review: Structural and Functional Aspects of Platelets. World Journal of Pharmacy and Pharmaceutical Sciences 12(4): 886-902.
-
Becker GJ, Hewitson TD (2013) Animal Models of Chronic Kidney Disease: useful but not Perfect. Nephrol Dial Transplant 28(10): 2432-2438.
-
Ebefors K, Lassen E, Anandakrishnan N, Azeloglu EU, Daehn IS (2021) Modeling the Glomerular Filtration Barrier and Intercellular Crosstalk. Front Physiol 12: 689083.
-
Shirinskiĭ VP (2011) Molecular Physiology of the Endothelium and Mechanisms of Vascular Permeability. Usp Fiziol Nauk 42(1): 18-32.
-
Wilhelm I, Fazakas C, Krizbai IA (2011) In Vitro Models of the Blood-Brain Barrier. Acta Neurobiol Exp (Wars) 71(1): 113-128.
-
Gerhardt H, Golding M, Fruttiger M, Ruhrberg C, Lundkvist A, et al. (2003) VEGF Guides Angiogenic Sprouting Utilizing Endothelial Tip Cell Filopodia. J Cell Biol 161(6): 1163-1177.
-
(2024) Cell Membrane. The Editors of Encyclopaedia Britannica.
-
Rucker HK, Wynder HJ, Thomas WE (2000) Cellular Mechanisms of CNS Pericytes. Brain Research Bulletin 51(5): 361-369.
-
Bergers G, Song S (2005) The Role of Pericytes in Blood- Vessel Formation and Maintenance. Neuro Oncol 7(4): 452-464.
-
El-Khoury N, Braun A, Hu F, Pandey M, Nedergaard M, et al. (2006) Astrocyte end-feet in germinal matrix, cerebral cortex, and white matter in developing infants. Paediatric Research 59(5): 673-679.
-
Brown LS, Foster CG, Courtney JM, King NE, Howells DW, et al. (2019) Pericytes and Neurovascular Function in the Healthy and Diseased Brain. Frontiers in Cellular Neuroscience 13: 282.
-
Luissint AC, Federici C, Guillonneau F, Chretien F, Camoin L, et al. (2012) Guanine nucleotide-binding protein Galphai2: a new partner of claudin-5 that regulates tight junction integrity in human brain endothelial cells. Journal of Cerebral Blood Flow and metabolism 32(5): 860-873.
-
Vorbrodt AW, Dobrogowska DH (2003) Molecular anatomy of intercellular junctions in brain endothelial and epithelial barriers: electron microscopist’s view. Brain Research Reviews 42(3): 221-242.
-
Stamatovic SM, Johnson AM, Keep RF, Andjelkovic AV (2016) Junctional proteins of the blood-brain barrier: New insights into function and dysfunction. Tissue Barriers 4(1): e1154641.
-
Abbott NJ, Patabendige AA, Dolman DE, Yusof SR, Begley DJ (2010) Structure and function of the blood-brain barrier. Neurobiol dis 37(1): 13-25.
-
Zhao Y, Gan L, Ren L, Lin Y, Ma C, et al. (2022) Factors influencing the blood-brain barrier permeability. Brain Research 1788: 147937.
-
Ali J, Liao F, Martens E, Muller WA (1997) Vascular endothelial cadherin (VE-cadherin): cloning and role in endothelial cell-cell adhesion. Microcirculation 4(2): 267-277.
-
Rho SS, Ando K, Fukuhara S (2017) Dynamic regulation of vascular permeability by vascular endothelial cadherin- mediated endothelial cell-cell Junctions. Journal of Nippon Medical School 84(4): 148-159.
-
Kuo WT, Odenwald MA, Turner JR, Zuo L (2022) Tight junction proteins occluding and ZO-1 as regulators of epithelial proliferation and survival. Annals of New York Academy of Science 1514(1): 21-33.
-
Agarwal V, Stephen T, Johnson SA, Freund JM, Siu BF, et al. (2011) Recruitment of progenitor cells, by an extracellular matrix cryptic peptide, in mouse model, of digit amputation. Tissue Engineering Part-A 17(19-20): 2435-2443.
-
Turowski P, Bridget-Ann K (2015) The blood brain barrier and methamphetamine: open sesame?. Frontiers in Neuroscience 9: 156.
-
Setchell BP, Voglmayer JK, Waites GM (1969) A blood- testis barrier restricting passage from blood into rete testis fluid but not into lymph. J. Physiology 200(1): 73- 85.
-
Franca LR, Hess RA, Dufore JM, Hofmann MC, Griswold MD (2016) The Sertoli cells: one hundred fifty years of beauty and plasticity. Andrology 4(2): 189-212.
-
Setchell BP, Main SJ (1975) The blood-testis barrier and steroids. Current Topics in Molecular Endocrinology 2: 223-233.
-
Setchell BP, Davies RV, Gladwell RT, Hinton BT, Main SJ, et al. (1978) The movement of fluid in the seminiferous tubules and rete testis. Annals of Biology Animal Biochemistry and Biophysics 18(2B): 623-632.
-
Fröhlich E (2002) [Structure and function of blood- tissue barriers]. Deutsche Medicinische Wöchenschrift 127(49): 2629-2634.
-
Green KJ, Getsios S, Troyanovsky S, Godsel LM (2010) Intercellular junction assembly, dynamics, and homeostasis. Cold Spring Harbor Perspective Biology 2(2): a000125.
-
Cavallaro U, Dejana E (2011) Adhesion molecule signalling: not always a sticky Business. Nature Reviews Molecular Cell Biology 12(3): 189-197.
-
Krutovskikh VA, Piccoli C, Yamasaki H (2002) Gap junction intercellular communication propagates cell death in cancerous cells. Oncogene 21(13): 1989-1999.
-
Dupont M, Souriant S, Lugo-Villarino G, Maridonneau- Parini I, Vérollet C (2018) Tunnelling Nanotubes: Intimate Communication between Myeloid Cells. Frontiers in Immunology 9: 43.
-
Bloemendal S, Kück U (2013) Cell-to-cell communication in plants, animals, and fungi: a comparative review. Naturwissenschaften 100(1): 3-19.
-
Harjunpaa H, Llort AM, Guenther C, Fagerholm SC (2019) Cell adhesion molecules and their roles and regulation in the Immune and tumor Microenvironment. Front Immunol 10: 107.
-
Nguyen TA, Pang KC, Masters SL (2017) intercellular communication for innate immunity. Mol Immunol 86: 16-22.
-
Niel G, Angelo G, Raposo G (2018) Shedding light on the cell biology of extracellular vesicles. Nature Reviews Molecular Cell Biology 19(4): 213-228.
-
Grecco HE, Schmick M, Bastiaens PI (2011) Signaling from the living plasma Membrane. Cell 144(6): 897-909.
-
Janmay PA, Miller RT (2011) Mechanisms of mechanical signaling in development and disease. J Cell Sci 124(Pt 1): 9-18.
-
Helvert S, Storm C, Friedl P (2018) Mechanoreciprocity in cell migration. Nature Cell Biology 20(1): 8-20.
-
Swartz MA, Tschumperlin DJ, Kamm RD, Drazen JM (2001) Mechanical stress is communicated between different cell types to elicit matrix remodeling. PNAS 98(11): 6180-6185.
-
Winer JP, Oake S, Janmey PA, (2009) Non-linear elasticity of extracellular matrices enable contractile cells to communicate local position and orientation. PLoS One 4(7): e6382.
-
Reinhart KCA, Dembo M, Hammer DA (2008) Cell- cell mechanical communication through compliant substrates. Biophys J 95(12): 6044-6051.
-
Kaji H, Unal CG, Langer R, Khademhosseini A (2011) Engineering systems for the generation of patterned co- cultures for controlling cell-cell interactions. Biochim Biophys Acta 1810(3): 239-250.
-
Hernandez V, Quiros M, Nusrat A (2017) Intestinal epithelial claudins: expression and regulation in homeostasis and inflammation. Ann N Y Acad Sci 1397(1): 66-79.
-
Balda MS, Matter K (2023) tight junctions. Curr Biol 33(21): R1135-R1140.
-
Peng YJ, Zhang H, Wong G, Feng MG, Ying SH (2024) MARVEL family Proteins contribute to vegetative growth, development and virulence of insect fungal pathogen Beauveria bassiana. J Invertebr Pathol 203: 108076.
-
Hartsock A, Nelson WJ (2008) Adherens and tight junctions: structure, function and connections to the actin cytoskeleton. Biochim Biophys Acta 1778(3):660- 669.
-
Cording J, Berg J, Käding N, Bellmann C, Tscheik C, et al. (2013) In tight junctions, claudins regulate the interactions between occludin, tricellulin and marvelD3, which, inversely, modulate claudins oligomerization. Journal of Cell Science 126(2): 554-564.
-
Saitou M, Furuse M, Sasaki H, Schulzke JD, Fromm M, et al. (2000) Complex phenotype of mice lacking occludin, a component of tight junction strands. Molecular Biology of the Cell 11(12): 4131-4142.
-
Traweger A, Fang D, Liu YC, et al. (2002) the tight junction-specific protein occludin is a functional target of the E3 ubiquitin-protein ligase itch. J Biol Chem 277(12): 10201-10208.
-
Bauer H, Mayer JZ, Steinbacher P, Lametschwandtner AH, Bauer HC (2010) the dual role of _zona occludens_ (ZO) proteins. Biomedical and Biotechnology 2010: 402593.
-
Cui X, Zhang X, Yin Q, Meng A, Su S, et al. (2014) F actin cytoskeleton reorganization is associated with hepatic stellate cell activation. Mol Med Rep 9(5): 1641-1647.
-
Schorey CD (2005) disassembling adherens junctions: Breaking up is hard to do. Trends Cell Biol 15(1): 19-26.
-
Yeh TH, Krauland L, Singh V, Zou B, Devaraj P, et al. (2010) Liver-specific β-catenin knockout mice have Bile canalicular abnormalities, bile secretory defect, and intrahepatic cholestasis. Hepatology 52(4): 1410-1419.
-
Pradhan-Sundd T, Monga SP (2019) Blood-Bile Barrier: Morphology,Regulation, and Pathophysiology. Gene Expression 19(2): 69-87.
-
Pradhan-Sundd T, Zhou L, Vats R, Jiang A, Molina L, et al. (2018) Dual catenin loss in murine liver causes tight junctional deregulation and progressive intrahepatic cholestasis. Hepatology 67(6): 2320-2337.
-
Behari J, Yeh TH, Krauland L, Otruba W, Cieply B, et al. (2010) Liver-specific beta-catenin knockout mice exhibit defective bile acid and cholesterol homeostasis and increased susceptibility to diet-induced steatohepatitis. American Journal of Pathology 176(2): 744-753.
-
Reed MA, Zhou C, Muller CJ, Burgin TP, Tour JM (1997) Conductance of a molecular junction. Science 278(5336): 252-254.
-
Kojima T, Murata M, Yamamoto T, Lan M, Imamura M, et al. (2009) Tight junction protein and signal transduction pathway in hepatocytes. Histology Histopathology 24(11): 1463-1472.
-
Luissint AC, Artus C, Glacial F, Ganeshamoorthy K, Couraud PO (2012) Tight junctions at the blood brain barrier: physiological architecture and disease- associated dysregulation. Fluids Barriers CNS 9(1): 23.
-
Luissint AC, Federici C, Guillonneau F, Chretien F, Camoin L, et al. (2012) Guanine nucleotide-binding protein Galphai2: a new partner of claudin-5 that regulates tight junction integrity in human brain endothelial cells. Journal of Cerebral Blood Flow metabolism 32(5): 860- 873.
-
Patel SJ, Milwid JM, King KR, Bohr S, Iracheta-Vellve A, et al. (2012) Gap junction inhibition prevents drug- induced liver toxicity and fulminant hepatic failure. Nature Biotechnology 30(2):179-183.
-
Bhattacharya A, Kaushik DK, Lozinski BM, Yong VW (2020) Beyond barrier functions: Roles of pericytes in homeostasis and regulation of neuroinflammation. Journal of Neuroscience Research 98(12): 2390-2405.
-
Wu D, Chen Q, Chen X, Han F, Chen Z, et al. (2023) The blood–brain barrier: structure, regulation, and drug delivery. Signal Transduction Targeted Therapy 8(1): 217.
-
Harilal S, Jose J, Parambi DGT, Kumar R, Unnikrishnan MK, et al. (2020) Revisiting the blood-brain barrier: A hard nut to crack in the transportation of drug molecules. Brain Research Bulletin 160: 121-140.
-
Paradis T, Bègue H, Basmaciyan L, Dalle F, Bon F (2021) Tight Junctions as a Key for Pathogens Invasion in Intestinal Epithelial Cells, International. Journal of Molecular Sciences 22(5): 2506.
-
Greene G, Campbell M (2016) Tight junction modulation of the blood brain barrier: CNS delivery of small molecules. Tissue Barriers 4(1): e1138017.
-
Oldendorf WH, Cornford ME, Brown WJ (1977) The large apparent work capability of the blood-brain barrier: a study of the mitochondrial content of capillary endothelial cells in brain and other tissues of the rat. Annals of Neurology 1(5): 409-417.
-
Daneman R, Prat A (2015) Blood brain barrier. Cold Spring Harbor Perspectives in Biology 7(1): a020412.
-
Tietz S, Engelhardt B (2015) Brain barriers: Crosstalk between complex tight junctions and adherens junctions. Journal of Cell Biology 209(4): 493-506.
-
Santa-Maria AR, Walter FR, Figueiredo R, Kincses A, Vigh JP, et al. (2021) Flow induces barrier and glycocalyx- related genes and negative surface charge in a lab-on- a-chip human blood-brain barrier model. Journal of Cerebral Blood Flow and Metabolism 41(9): 2201-2215.
-
Sofroniew MV, Vinters HV (2010) Astrocytes: biology and pathology. Acta Neuropathology 119(1): 7-35.
-
Yang BA, Westerhof TM, Sanon K, Merajaver SD, Aguilar CA (2020) Engineered tools to study intercellular communication. Advanced Sci (Weinh) 8(3): 2002825.
-
McConnell HL, Mishra A (2022) Cells of the Blood-Brain Barrier: An Overview of the neurovascular Unit in Health and Disease. Methods Mol Bioliology 2492: 3-24.
-
Mazumder AG, Julé AM, Cullen PF, Sun D (2022) Astrocyte heterogeneity within white matter tracts and a unique subpopulation of optic nerve head astrocytes. iScience 25(12): 105568.
-
Lin A, Peiris NJ, Dhaliwal H, Hakim M, Li W, et al. (2021) Mural Cells: Potential Therapeutic Targets to Bridge Cardiovascular Disease and Neurodegeneration. Cells 10(3): 593.
-
Hartmann DA, Berthiaume AA, Grant RI, Harrill SA, Koski T, et al. (2021) Brain capillary pericytes exert a substantial but slow influence on blood flow. Nature Neurosciences 24(5): 633-645.
-
Dong X (2018) Current Strategies for Brain Drug Delivery. Theranostics 8(6): 1481-1493.
-
Ahlawat J, Guillama Barroso G, Masoudi Asil S, Alvarado M, Armendariz I, et al. (2020) Nanocarriers as Potential Drug Delivery Candidates for Overcoming the Blood- Brain Barrier: Challenges and Possibilities. ACS Omega 5(22): 12583-12595.
-
Gumbiner B (1987) Structure, biochemistry, and assembly of epithelial tight junctions. American Journal of Physiology 253(6 Pt 1): C749-758.
-
Tsukita S, Furuse M, Itoh M (2001) Multifunctional strands in tight junctions. Nature Reviews Molecular Cell Biology 2(4): 285-293.
-
Haseloff RF, Dithmer S, Winkler L, Wolburg H, Blasig IE (2015) Transmembran proteins of the tight junctions at the blood-brain barrier: structural and functional aspects. Seminars in Cell Developmental Biology 38: 16- 25.
-
Ikenouchi J, Furuse M, Furuse K, Sasaki H, Tsukita S, et al. (2005) Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. Journal of Cell Biology 171(6): 939-945.
-
Furuse M, Hirase T, Itoh M, Nagafuchi A, Yonemura S, et al. (1993) Occludin: a novel integral membrane protein localizing at tight junctions. Journal of Cell Biology 123(6 Pt 2): 1777-1788.
-
Tajes M, Ramos-Fernández E, Weng-Jiang X, Bosch- Morató M, Guivernau B, et al. (2014) The blood-brain barrier: structure, function and therapeutic approaches to cross it. Molecular Membrane Biology 31(5): 152-167.
-
Liebner S, Dijkhuizen RM, Reiss Y, Plate KH, Agalliu D, et al. (2018) Functional morphology of the blood-brain barrier in health and disease. Acta Neuropathology 135(3): 311-336.
-
Willis CL, Meske DS, Davis TP (2010) Protein kinase C activation modulates reversible increase in cortical blood-brain barrier permeability and tight junction protein expression during hypoxia and posthypoxic reoxygenation. Journal of Cerebral Blood Flow Metabolism 30(11): 1847-1859.
-
Malinova TS, Huveneers S (2018) Sensing of cytoskeletal forces by asymmetric adherens junctions. Trends in Cell Biology 28(4): 328-341.
-
Dash S, Duraivelan K, Samanta D (2021) Cadherin- mediated host-pathogen interactions. Cell Microbiology 23(5): e13316.
-
Kwak M, Southard KM, Kim WR, Lin A, Kim NH, et al. (2022) Adherens junctions organize size-selective proteolytic hotspots critical for Notch signalling. Nature Cell Biology 24(12): 1739-1753.
-
Hess AR, Franco LR (2005) Structure of the Sertoli cell. Sertoli cell Biology, Academic Press Elsevier Press, pp: 19-40.
-
Griswold MD (2016) Spermatogenesis: The Commitment to Meiosis. Physiology Rev 96(1): 1-17.
-
Gawdi R, Shumway KR, Emmady PD (2023) Physiology, Blood Brain Barrier. Treasure Island (FL): Stat-Pearls Publishing, USA.
-
Cheng CY, Mruk DD (2010) The biology of spermatogenesis: the past, present and future. Philosophical Transaction of Royal Society 365: 1459- 1463.
-
Meinhardt A, Hedger MP (2011) Immunological, paracrine and endocrine aspects of testicular immune privilege. Molecular Cell Endocrinology 335(1): 60-68.
-
Lara NLEM, Costa GMJ, Figueiredo AFA, de França LR (2020) The Sertoli cell: what can we learn from different vertebrate models?. Animal Reproduction 16(1): 81-92.
-
Barrett KE, Barman SM, Boitano S, Brooks H (2012) Ganong’s Review of Medical Physiology. In: 24th edition (Edn.), McGraw-Hill Education, pp: 412-420.
-
Sharma S, Hanukoglu A, Hanukoglu I (2018) Localization of epithelial sodium channel (ENaC) and CFTR in the germinal epithelium of the testis Sertoli Cells and spermatozoa. Journal of Molecular Histology 49(2): 195-208.
-
Gerhardt H, Golding M, Fruttiger M, Ruhrberg C, Lundkvist A, et al. (2003) VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. Journal of Cell Biology 161(6): 1163-1177.
-
Goldmann EE (1909) The external and internal secretion of the healthy and sick organism in the light of the “vital coloration. Beitr Klin Chir 64: 192-265
-
de Kretser DM (1990) Germ cells-Sertoli cell interactions. Reproduction, Fertility and Development 2(3): 225-235.
-
Bergmann M, Schindelneisser J (1987) Development of blood-testis barrier in domestic fowl (Gallus domesticus). International Journal of Andrology 10(2): 481-488.
-
Leal MC, deWaal PP, Garcia-Lopez A, Chen SX, Bogerd J, et al. (2009) Zebrafish primary testis tissue culture: an approach to study testis function ex-vivo. General and Comparative Endocrinology 162(2): 134-138.
-
Cheng CY, Mruk DD (2012) The blood-testis barrier and its implications for male contraception. Pharmacological Reviews 64(1): 16-64.
-
Ahmed N, Yufei H, Yang P, Muhammad Yasir W, Zhang Q, et al. (2016) Cytological study on Sertoli cells and their interactions with germ cells during annual reproductive cycle in turtle. Ecology and Evolution 6(12): 4050-4064.
-
Schweigert FJ, Gericke B, Wolfram W, Kaisers U, Dudenhausen JW (2006) Peptide and protein profiles in serum and follicular fluid of women undergoing IVF. Human Reproduction 21(11): 2960-2968.
-
Sui Michelle, Cheng CY (2013) The blood-follicle barrier (BFB) in disease and in ovarian function. Advances in Experimental Medicine and Biology 763: 186-192.
-
Mariana M, Frances G, Andrea C, Giorgia M, Alessandro T (2022) Reverse electrodialysis heat engine (REDHE) in Salinity Gradient Heat Engins.
-
Garcia-ascual A, Labadia A, Triguero D, Costa G (1996) Local regulation of oviductal blood flow. General Pharmacology 27(8): 1303-1310.
-
Coy P, García-Vázquez FA, Visconti PE, Avilés M (2012) Roles of the oviduct in mammalian fertilization. Reproduction 144(6): 649-660.
-
Eder S, Müller K, Chen S, Schoen J (2022) Long-term culture of feline oviduct epithelial cells on permeable filter supports. Cytotechnology 74(5): 531-538.
-
Powell DW (1981) Barrier function of epithelium. American Journal of Physiology 244(4): G275-G288.
-
Chen S, Einspanier R, Schoen J (2015) Transepithelial electrical resistance (TEER): a functional parameter to monitor the quality of oviduct epithelial cells cultured on filter supports. Histochemistry and Cell Biology 144(5): 509-515.
-
Erlebacher A (2013) Immunology of the maternal- foetal interface. Annual Review of Immunology 31(1): 387-411.
-
McRae AC (1988) The blood-uterine lumen barrier and exchange between extracellular fluids. Journal of Reproduction and Fertility 82(2): 857-873.
-
Ayala FJ (2007) Darwin’s greatest discovery design: design without designer. Proceedings of National Academy of Science 104(1): 8567-8573.
-
Slater M, Aburto MR, Acker-Palmer A, (2021) Blood- Brain Barrier Dynamics to Maintain Brain Homeostasis. Trends in Neurosciences 44(5): 393-405.
-
Tizard IR (2023) The primates: humans and their relatives. Comparative mammalian immunology, Academic Press, pp:375-395.
-
Reznick DN, Mateos M, Springer MS (2002) Independent origins and rapid evolution of the placenta in the fish genus Poeciliopsis. Science 298(5595): 1018- 1020.
-
Blackburn DG, Avanzati AM, Paulesu L (2015) Classics revisited. History of reptile placentology: Studiati’s early account of placentation in a viviparous lizard. Journal of Placenta 36(11): 1207-1211.
-
Blundell C, Tess ER, Schanzer AS, Coutifaris C, Su EJ, et al. (2016) A microphysiological model of the human placental barrier. Lab Chip 16(16): 3065-3073.
-
Jensen OE, Hernyavsky IL (2019) Blood flow and transport in the human placenta. Annu Rev Fluid Mech 51: 25-47.
-
Sadler TW (2012) Langman’s Medical Embryology. In: 12_th_ Edition (Edn.), Wolters Kluwer, Lippincot William &Wilkins, Philadelphia, USA.
-
Griffiths SK, Campbell JP (2015) Placental structure, function and drug transfer. Continuing Education in Anaesthesia Critical Care & Pain 15(2): 84-89.
-
Krishna U, Bhalerao S (2011) Placental insufficiency and fetal growth restriction. Journal of Obstetrics and Gynaecology of India 61(5): 505-511.
-
Arumugasaamy N, Rock KD, Kuo CY, Bale TL, Fisher JP (2020) Microphysiological systems of the placental barrier. Advances in Drug Deliveries Reviews 161-162: 161-175.
-
Sastry BV (1999) Techniques to study human placental transport. Advanced Drug Delivery Reviews 38(1): 17-39.
-
Kitano T, Iizasa H, Hwang IW, Hirose Y, Morita T, et al. (2004) Conditionally immortalized syncytiotrophoblast cell lines as new tools for study of the blood-placenta barrier. Biological and Pharmaceutical Bulletin 27(6): 753-759.
-
Costa MA (2016) The endocrine function of human placenta: an overview. Reproductive Biomedicine Online 32(1): 14-43.
-
Brown LS, Foster CG, Courtney JM, King NE, Howells DW, et al. (2019) Pericytes and Neurovascular Function in the Healthy and Diseased Brain. Frontiers in Cellular Neuroscience 13: 282.
-
Nishiguchi A, Gilmore C, Sood A, Matsusaki M, Collett G, et al. (2019) In vitro placenta barrier model using primary human trophoblasts, underlying connective tissue and vascular endothelium. Biomaterials 192: 140- 148.
-
Pemathilaka RL, Reynolds DE, Hashemi NN (2019) Drug transport across the human placenta: review of placenta-on-a-chip and previous approaches. Interface Focus 9(5): 20190031.
-
Kappen C, Kruger C, MacGowan J, Salbaum JM (2012) Maternal diet modulations placenta growth and gene expression in a mouse model diabetic pregnancy. PLOS ONE 7(6): e38445.
-
Dotiwala AK, McCausland C, Samra NS (2023) Anatomy, Head and Neck: Blood Brain Barrier. StatPearls [Internet].
-
Madhusoodanan J (2018) Break on through: how some viruses infect placenta. Knowable Magazine.
-
Pereira L (2018) Congenital viral infection: traversing the uterine-placental interface. Annual Review of Virology 5(1): 273-299.
-
Uhlén M, Fagerberg L, Hallström BM, Lindskog C, Oksvold P, et al. (2015) Proteomics: Tissue-based map of the human proteome. Science 347(6220): 1260419.
-
Andrews S, Krueger C, Mellado-Lopez M, Hemberger M, Dean W, et al. (2023) Mechanisms and function of de novo DNA methylation in placental development reveals an essential role for DNMT3B. Nature Communications 14(1): 371.
-
Tietz PS, Larusso NF (2006) Cholangiocyte biology. Current Opinion in Gastroenterology 22(3): 279-287.
-
Kojima T, Yamamoto T, Murata M, Chiba H, Kokai Y, et al. (2003) Regulation of the blood-biliary barrier: Interaction between gap and tight junctions in hepatocytes. Medical Electron Microscopy 36(3): 157- 164.
-
Rao RK, Samak G (2013) Bile duct epithelial tight junctions and barrier function. Tissue Barriers 1(4): e25718.
-
Renuke HG, Cotran RS, Venkatachalam MA (1975) Role of molecular charge in glomerular permeability. Tracer studies with cationized ferritins. Journal of Cell Biology 67(3): 638-646.
-
Johansen KL, Chertow GM, Foley RN, Gilbertson DT, Herzog CA, et al. (2020) 2020 USRDS Annual Data Report: Epidemiology of Kidney Disease in the United States. Am J Kidney Dis 77(4S1): A7-A8.
-
Daehn IS, Duffield JS (2021) The glomerular filtration barrier: a structural target for novel kidney therapies. Nature Review Drug Discovery 20(10): 770-788.
-
Dalal R, Bruss ZS, Sahdev JA (2023) Physiology, Renal blood flow and filtration. Statpearls (internet).
-
Menon MC, Chuang PY, He CJ (2012) The glomerular filtration barrier: components and crosstalk. Int J Nephrol 2012: 749010.
-
Scott RP, Quaggin SE (2015) Review series: The cell Biology of renal filtration. J cell Biology 209: 199-210.
-
Grahammer F, Wigge C, Schell C, Kretz O, Patrakka J, et al. (2016) A flexible, multilayered protein scaffold maintains the slit in between glomerular podocytes. Journal of Clinical Investigation Insight 1(9): e86177.
-
Leiby KL, Raredon MSB, Niklason LE (2020) Bioengineering the Blood-gas Barrier. Comprehensive Physiology 10(2): 415-452.
-
Aros CJ, Pantoja CJ, Gomperts BN (2021) Wnt signalling in lung development, regeneration and disease progression. Communication Biology.
-
Crapo JD, Barry BE, Gehr P, Bachofan M, Weibei ER (1982) Cell numbers and cell characterization of normal human lung. American Review of Respiratory Disease 126(2): 332-337.
-
Beers MF, Morrisey EE (2011) The three R’s of lung health and disease: repair, remodelling, and regeneration. Journal of Clinical Investigations 121(6): 2065-2073.
-
Dorrello NV, Guenthart BA, O’Neill JD, Kim J, Cunningham K, et al. (2017) Functional vascularized lung grafts for lung bioengineering. Science Advances 3(8): e1700521.
-
Lefèvre-Utile A, Braun C, Haftek M, Aubin F (2021) Five Functional Aspects of the Epidermal Barrier. International Journal of Molecular Science 22(21): 11676.
-
Elias PM (2008) Skin barrier function. Current Allergy Asthma Reports 8(4): 299-305.
-
Feingold KR (2012) Lamellar bodies: The key to cutaneous barrier function. Journal of Investigative Dermatology 132(8): 1951-1953.
-
Brown SJ, McLean WH (2012) One remarkable molecule: Filaggrin. Journal of Investigative Dermatology 132(3 P2): 751-762.
-
Briot J, Simon M, Méchin MC (2020) Deimination, Intermediate Filaments and Associated Proteins. International Journal of Molecular Science 21(22): 8746.
-
Quiroz FG, Fiore VF, Levorse J, Polak L, Wong E, et al. (2020) Liquid-liquid phase separation derives skin barrier. Science 367(6483): eaax9554.
-
Bäsler K, Bergmann S, Heisig M, Naegel A, Zorn- Kruppa M, et al. (2016) The role of tight junctions in skin barrier function and dermal absorption. Journal of Controlled Diseases 242: 105-118.
-
Churko JM, Laird DW (2013) Gap junction remodeling in skin repair following wounding and disease. Physiology (Bethesda) 28(3): 190-198.
-
Johnson JL, Najor NA, Green KJ (2014) Desmosomes: Regulators of cellular signaling and adhesion in epidermal health and disease. Cold Spring Harbor Perspectives in Medicine 4(11): a015297.
-
Li MW, Mruk DD, Cheng CY (2012) Gap junctions and blood-tissue barriers. Advances in Experimental Medicine and Biology 763: 260-280.
-
Chen MS, Hou PK, Tai TY, Lin BJ (2008) Blood ocular barrier. Tzu Chi Medical Journal 20(1): 25-34.
-
Cunha-Vaz JG, Maurice DM (1967) The active transport of fluorescein by the retinal vessels and retina. Journal of Physiology 191(3): 467-486.
-
Giguère S, Prescott JF, Desmond BJ (2006) Antimicrobial therapy in veterinary medicine. Wiley Blackwell pp: 366.
-
Maggs DJ, Miller PE, Ofri R, Slatter DH (2008) Slatter’s fundamentals of veterinary ophthalmology. Elsevier Health Sciences pp: 455.
-
Coscas G, Cunha-Vaz J, Loewenstein A, Soubrane G (2010) Macular Edema: A Practical Approach. Karger Publishers, pp: 59.
-
Ribbatti D (2015) The discovery of blood-thymus barrier. Immunology Letters 168(2): 325-328.
-
Pearse G (2006) Normal structure, function and histology of thymus. Toxicologic Pathology 34(5): 504- 514.
-
Bruss DM, Ely S (2023) Anatomy, Head and Neck: Blood Thymus barrier.
-
Nagatake T, Zhao YC, Ito T, Itoh M, Kometani K, et al. (2021) Selective expression of claudin-5 in thymic endothelial cells regulates the blood-thymus barrier and T cell export. Int Immunol 33(3): 171-182.
-
Lkhagvasuren B, Mee-Inta O, Zhao ZW, Hiramoto T, Boldbaatar D, et al. (2021) Pancreas-Brain Crosstalk. Frontiers in Neuroanatomy 15: 691777.
- Evaluation of Skin Aging Preventive Effects of Cherry Blossom Petal Extracts Through Antioxidant and Anti-Glycation Activities
- Is Cell Death Responsible for False Positive Results of In Vivo Comet Assay?
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism