Immunomodulatory Effects of Bio-Clean II on T-Helper 4 and Cytotoxic T-Lymphocytes in Rats Exposed to Purified Bacterial Lipopolysaccharide
The review analyzed data on Immunomodulatory action of cholera toxin B subunit (CT-B) and the synthetic peptide LKEKK that corresponds to residues 16-20 in thymosin-α1 and 131-135 in interferon-α2 on the functional, NO-synthase and guanylate cyclase activity of T and B lymphocytes, of macrophage-like cell line RAW 264.7. According to the data presented, CT-B and the peptide bind to the cholera toxin receptor of the target cell with high affinity and trigger the following cascade of intracellular reactions: activation of inducible NO synthase → increase in NO production → increase in soluble guanylate cyclase activity → increase in the cyclic guanosine-3’,5’-monophosphate level.
Introduction
The use of herbal remedies to cure and manage a variety of diseases has grown increasingly popular as part of African Traditional Medicine (ATM) [1, 2, 3]. In Nigeria, herbal treatments are widely utilized to cure illnesses such as malaria, typhoid, headache, fever, pile, diarrhea, constipation, and arthritis, to name a few [4, 5]. Herbal medicines have found their way into modern medicine due to its antibacterial, antiviral, and anti-inflammatory properties [6, 7, 8, 9]. The herbal remedy “Bio-Clean II” for instance, has been demonstrated to boost immunity and fights viral infections like HIV. It has been shown to induce an increase in CD4 Cell Count and body weight, while decreasing viral load in a group of HIV positive women [10, 11]. It has also been shown to modulate the levels of inflammatory cytokines including Interleukin-6 (IL- 6), Interleukin-10 (IL-10), Tissue Necrosis Factor-Alpha (TNF-α) and Interferon-Gamma (IFN-γ) in experimental lipopolysaccharide (LPS)-induced inflammation using animal model [12]. Many of the preventive and therapeutic effects of Bio-Clean II are directly or indirectly attributable to its immunomodulatory properties [13] due to the presence of some important phytochemicals including resin, alkaloids, saponins, anthraquinones, tannin, and Cardiac Glycoside as reported in our previous studies [9]. Bacterial lipopolysaccharides (LPSs) are a major component of Gram-negative bacteria’s outer membrane. Invading microbes are known to produce glycol conjugates, which are recognized as a molecular signature. LPSs are amphiphilic polymers that have three distinct genetic, structural, and functional regions: the lipid A, the core oligosaccharide, and a polysaccharide component, and the O-chain [14].
Exposure to bacterial lipopolysaccharide (LPS) has been associated with inflammation of body tissues characterized by swelling, redness, pain, and loss of function if the assaulting agent persists [15, 16, 17]. LPS is recognized by the immune system through a complex mechanism including molecules that bind to LPS and pass it on to cell membrane-associated receptors on leukocytes, endothelial cells, and other cells, triggering a pro-inflammatory cascade [15, 18, 19]. T cells, cytokines, catecholamines, and corticosterone, among other things, influence immune system modulation. Lympocyte- activation and proliferation are critical during the assault of infection or other pathological states such as inflammation, as they are the host’s major guardians against disease [20, 21]. Generally speaking, T cells are in charge of the cell- mediated arm of the adaptive immune system, and they are primarily concerned with cellular immune responses to intracellular invaders like viruses (as well as the regulation of B cell responses). They use cell surface receptors (T cell receptors) to identify antigen. Different subsets of T cells exist with specific and diverse immunological functions [22].
T helper cells (Th cells), for example, are a type of T cell that play an important role in the adaptive immune system. They are also known as CD4+ cells because they express CD4 on their surfaces. They “help” the function of other immune cells by releasing cytokines, as their name implies [23, 24, 25]. These cells aid in polarizing the immune response into the appropriate kind, depending on the nature of the immunological attack. They are assumed to be vital for B cell antibody class switching, dendritic cell cross-tolerance, activating and developing cytotoxic T cells, and enhancing the bactericidal activity of phagocytes like macrophages and neutrophils [26]. When MHC class II molecules, which are expressed on the surface of antigen-presenting cells, present them with peptide antigens, they become activated (APCs). They divide quickly after activation and release cytokines that govern or support the immune response. IL-2 and IFN are mainly released by activated CD4+ cells [27]. On the other hand, a cytotoxic T lymphocyte (also known as CD8+ T-cell) is a type of white blood cell that destroys cancer cells, cells infected with intracellular pathogens (such viruses or bacteria), and cells that have been injured in some way [28]. The appearance of the CD8 protein on the cell surface distinguishes these cells. Short peptides (8-11 amino acids in length) associated with MHC class I molecules, which are found on the surface of all nucleated cells, help cytotoxic T lymphocytes recognize their targets.
Cytotoxic T cells could produce some IFN-γ. These cytokines affect the effector activities of other cells, such as macrophages and natural killer cells (NK cells). CD4+ or CD8+ memory T cells that express CD45RO are common [29]. In a rat model of arthritis, cytotoxic T lymphocytes have been linked to the progression of the disease by depleting glycosaminoglycans and other cartilage macromolecules [30]. Several animal investigations have revealed that the primary role of cytotoxic T cells is to promote inflammation. The generation of cytokines by CD8+ cells is thought to speed up the progression of arthritis [31]. For individuals suffering from CD4+ lymphopenia caused by a variety of infectious diseases, medications, or cancer, any treatment (orthodox or traditional) that promotes the proliferation of CD4+ and depletion of CD8+ cells has great therapeutic potential [21]. T cells are frequently affected by the presence of bacterial lipopolysaccharide [32]. To the best of our knowledge, this study is the first to assess the modulation of T-Helper 4 and Cytotoxic T-Lymphocytes as therapeutic mechanisms of Bio- Clean II in purified Bacterial Lipopolysaccharide-Induced Inflammation in Rats. This study was therefore designed to determine the effect of Bio-Clean II on CD4 and CD8 Cell Count in rats exposed to purified bacterial LPS in comparison to controls.
Materials and Methods
Study Design
Animal models are used in this analytic experimental investigation.
Duration of Study
The research work lasted for a period of 3 months (April- July, 2021).
Study Area
The research was conducted at Babcock University’s Experimental Animal House in Ilishan-Remo, Ogun State.
Source of Bio-Clean II
Bio-Clean II was procured from the manufacturer/ maker on demand.
Source of Purified Lipopolysaccharide
As lyophilized powder, purified Lipopolysaccharide® (E. coli 0127:B8, Sigma-Aldrich, St. Louis, MO, USA) was purchased in vials. It was reconstituted in sterile Phosphate Buffer Saline (PBS) and given as a single dose (5 mg/kg) intraperitoneally.
Standard Drugs
Diclofenac (50mg) and Ciprofloxacin (500mg) were purchased from the Pharmacy Unit, Babcock University Teaching Hospital, Ilishan-Remo, Ogun State. Twenty (20) mL of Ciprofloxacin and Diclofenac suspension was prepared daily by dissolving one tablet each in 20 mL of sterile distilled water to obtain a concentration of 25mg/ml of Ciprofloxacin and 2.5mg/ml of Diclofenac.
Experimental Animals
A total of 36 male albino rats weighing 150g±50g (mean±SD) were obtained from the University of Ibadan’s small animal house and were clinically screened for evidence of infection or abnormalities upon arrival. At the Experimental Animal House, Department of Animal Science, School of Agriculture and Industrial Technology, Babcock University, Ilishan-Remo, the animals were housed separately in well-ventilated wire-bottom steel cages under hygienic conditions at 25°C and a relative humidity of 45- 50%. (Ogun State, Nigeria).
The rats were randomly separated into six groups of six rats each and given a regular rat meal (10g/100g body weight) twice daily as well as free tap water. The rats were allowed to acclimate in the Animal House to a consistent 12-hour light-dark cycle for 14 days prior to the commencement of the investigation. All animal experiments were carried out in accordance with the Current Animal Care Regulations and Standards of the Institute for Laboratory Animal Research (ILAR) [33]. The experimental animals were treated as shown in the experimental pharmacological protocol below (Table 1). Group 1 received a single dose of 5mg/Kg of purified LPS intra-peritoneally (Inflammation control). Group 2 received 5mg/Kg of purified LPS, 50mg/Kg/bid Diclofenac and 500mg/kg/bid of Ciprofloxacin (Positive Control). Group 3 received 5mg/Kg of purified LPS and sterile Phosphate Buffer Saline (Negative control). Group 4 was not exposed to purified LPS, rather they received water and feed only throughout the experimental period (Zero Control). Group 5 and 6 were treated with Bio-clean II for 7 and 14 days, respectively, post-exposure to purified LPS. Therapy with Bio-clean II was initiated by day 2 post-LPS challenge (Table 1). The volume of Bio-clean II suspension given to each rat orally through an intragastric tube was determined and recorded prior to the start of the treatment period. After then, the rats were checked daily for any alterations. For the number of days stipulated in the experimental protocol, the study animals were administered a suitable volume of suspension (5ml/kg body weight) twice daily, every 12 hours (between the hours of 6.00-6.30 AM/PM). Before administration, the suspension was gently shaken and delivered via oral gavage. At days 8 and 15, the rats were sacrificed by cervical dislocation, as described by Ochei J, et al. [34], and cardiac blood samples were collected for evaluation of CD4 and CD8 Cell Counts.
| Groups | Treatments |
|---|---|
| G1(n=6) | Received 5mg/Kg of Purified Lipopolysaccharide (Inflammation control) |
| G2(n=6) | Received 5mg/Kg of Purified Lipopolysaccharide + 50mg/Kg/bid Diclofenac+500mg/Kg/bid of Ciprofloxacin (Positive control). |
| G3(n=6) | Received 5mg/Kg of purified Lipopolysaccharide + sterile Phosphate Buffer Saline (Negative control). |
| G4(n=6) | Received water and feed only (Zero control) |
| G5(n=6) | Received 5mg/Kg of Purified Lipopolysaccharide+ Bio-clean II for 7 days |
| G6(n=6) | Received 5mg/Kg of Purified Lipopolysaccharide+ Bio-clean II for 14 days |
Table 1: Experimental pharmacological protocol Where “n” = the number of rats in each group.
Collection of Blood Specimens
Cardiac blood specimen was collected from each experimental animal by terminal bleeding from the heart. The dead rat was placed on its back on a cork board and strapped across the forelegs and hind legs with two adhesive tapes. A 40 X 0.8mm needle was inserted in the center line at the tip of the sternum and pushed forward at 45oC until it punctured the heart. The needle was moved forward until blood began to flow into the syringe. The needle was rapidly withdrawn after collecting roughly 4ml of heart blood. Two milliliters (2 ml) of the blood specimen collected was put into a clean container with anticoagulant (EDTA) in preparation for CD4 cell count. The remaining two milliliters (2 ml) was transferred into a plain bottle to clot. The non- anticoagulated blood specimens were made to stand for about an hour to clot, retract and centrifuge afterwards at 3,500 rpm for 10 minutes at room temperature. The yielded serum was transferred to another clean sterile plain bottle and used for CD8 estimation. The used needle was disposed of in a disinfected jar and then autoclaved.
Laboratory Analyses
Evaluation of CD4+ Cell Counts
The CD4 cell count was evaluated using the cartridge BD FACSPresto Near-patient CD4 Counter according to the manufacturer’s instructions as described by Moon S, et al. [35].
Test principle
Fluorescence photomicroscopy and light absorbance detection on a multicolor platform are used to operate the instrument. LEDS illuminate on or through a sample, and the resulting signal is captured on a digital camera chip through a microscope lens and emission filter, or spectrometer equipment. Each test was performed with a cartridge containing dried reagents. An entrance port to the cartridge receives the whole blood from a capillary or venipuncture specimen. CD4 absolute count and the proportion of CD4+ T cells among lymphocytes (percent CD4) are all determined using the same cartridge. The software is built into the instrument, which is controlled via a touchscreen interface. The absolute CD4 count, percent CD4, and Hb concentration are all calculated automatically by the software algorithms. The findings can be viewed on the screen, printed, and exported.
Evaluation of CD8+ Cell Counts
CD8 cell count was estimated using Elabscience Enzyme linked immunosorbent assay (ELISA) CD8 test kit according to the manufacturer’s instruction as described by Kannangai R, et al. [36].
Test Principle
This ELISA CD8 test kit uses the sandwich-ELISA principle. This kit includes a micro ELISA plate that has been precoated with a Rat CD8 antibody. Samples or standards are placed in micro ELISA plate wells, which are then mixed with the relevant antibody. Each microplate well was treated with a biotinylated detection antibody specific for Rat CD8 and Avidin-Horserad Peroxidase (HRP) and incubated. Each well receives a free substrate solution. Only the wells containing Rat CD8 biotinylated detection antibody and Avidin-HRP conjugate will turn blue. The addition of stop solution stops the enzyme-substrate reaction, and the color changes to yellow. At a wavelength of 450nm, the optical density (00) was determined spectrophotometrically. The OD value is proportional to the sample’s CD8 concentration. The ELISA kit has a detection range of 1.25-80 ng/ml and a sensitivity of 0.75 ng/ml. This kit detects the presence of Rat CD8 in samples. There was no evidence of considerable cross- reactivity or interference between Rat CDS and analogues. The Coefficient of variation in terms of repeatability is ten percent.
Elabscience CD8 Reagent Preparation Procedure
Briefly, the kit was taken out of the refrigerator 20 minutes ahead of time before usage. The samples were also brought to room temperature and mixed properly, avoiding foaming in the process. A 1x working solution was made from the 25x concentrated wash buffer. Different concentrations of the reference standard were prepared and used. The 100x Biotinylated detection Ab/Ag and 100x concentrated HRP conjugate were both diluted to 1x working solution before use (15 minutes earlier).
Assay Procedure
Briefly, each well was filled with 100uL of standard (or sample) and incubated for 90 minutes at 37°C. The liquid was then removed and 10uL of Biotinylated detection Ab/ Ag was added, and incubated for 1 hour at 37oC. It was then extracted and washed three (3) times. Ninety (90) microliters of substrate reagent was added, and the mixture was incubated at 37°C for 15 minutes. The optical density (OD) value was immediately read at 450nm using a KC-100 Microplate ELISA reader after adding 50 microliters of stop solution. The outcomes were calculated.
Data Analyses
Data were evaluated using one-way analysis of variance (ANOVA) and the Turkey-Kramer Multiple Comparison Test with the SPSS-20.0 (Statistical Package for Social Scientists- version 20.0) statistical tool. P values of less than 0.05 were adjudged to be significant [37]. The mean of Six (6) rats were used to show CD4 and CD8 data using charts and Tables.
Results
This present study assessed the effects of “Bio-Clean II” herbal remedy on CD4 and CD8 cell counts in experimental LPS-induced inflammation using an animal model. Post exposure to LPS, physical examination of the rats in the inflammation control group, revealed the presence of swelling and redness especially in the feet of the rats.
Effect on CD4 Count
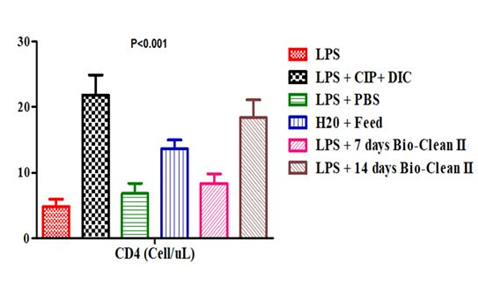
The CD4 count of the test and control rats posttreatment with Bio-clean II is represented using a bar chart (Figure 1). The CD4 count of the 7 days (Group 5: 8.40±1.44 Cells/ μl) and 14 days (Group 6: 18.40±2.66 Cells/μl) Bio-clean II treated rats was found to be significantly higher (p=0.040 and p=0.032, respectively) when compared to the inflammation control (Group 1: 4.80±1.16 Cells/μl). On the one hand, it was found to be non-significantly lower (p=0.117 and p=1.000, respectively) when compared to the positive control (Group 2: 21.80±3.06 Cells/μl), but on the other, it was found to be significantly higher (p=0.016) when compared to the negative control (Group 3: 6.80±1.59 Cells/μl). Furthermore; the CD4 count of the 7 days Bio-clean treated rats (Group 5: 8.40±1.44 Cells/μl) was also observed to be non-significantly lower (p=0.371) when compared to the zero control (Group 4: 13.60±1.36); but that of the 14 days (Group 6: 18.40±2.66 Cells/μl) was found to be significantly higher (p=0.026). In addition, the CD4 count of the 14 days Bio-clean treated rats (Group 6: 18.40±2.66 Cells/μl) when compared with that of the 7 days Bio-clean treated rats (Group 5: 8.40±1.44 Cells/μl) was found to be significantly higher (p=0.010). Meanwhile, the CD4 count of the positive control (Group 2: 21.80±3.06 Cells/μl) was found to be significantly higher (p=0.019) than of the negative control (Group 3: 6.80±1.59 Cells/μl).
Effect on CD8 Count
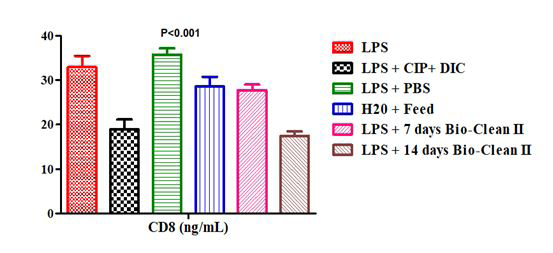
The CD8 count of the test and control rats post treatment with Bio-clean II is presented using a bar chart in Figure 2. The CD8 count of 7 days Bio-clean treated rats (Group 5: 27.67±1.32ng/mL) was found to be non-significantly lower (p=0.835) in comparison to the inflammation control (Group 1: 32.93±2.52ng/mL); while that of the 14 days Bio-clean II treated rats (Group 6: 17.44±1.00ng/mL) was significantly lower (p=0.029). In comparison to the positive control (Group 2: 18.92±2.22ng/mL), the CD8 count of 7 days (Group 5: 27.67±1.32ng/mL) and 14 days (Group 6: 17.44±1.00ng/ mL) Bio-clean treated rats were found to be non-significantly higher and lower, p=0.179 and p=1.000, respectively. Still, in comparison to the negative control (Group 3: 35.72±1.47ng/ mL), the CD8 count of the 7 days Bio-clean treated rats (Group
5: 27.67±1.32ng/mL) was found to be non-significantly lower, but that of the 14 days Bio-clean treated rats (Group 6: 17.44±1.00ng/mL) was significantly lower (p=0.000). Also, the CD8 count of the 7 days (Group 5: 27.67±1.32ng/mL) and 14 days (Group 6: 17.44±1.00ng/mL) Bio-clean treated rats was non-significantly lower (p=1.000 and p=0.068, respectively) when compared to the zero control (Group 4: 28.56±2.23ng/mL). Meanwhile, the CD8 count of the 14 days Bio-clean treated rats (Group 6: 17.44±1.00ng/mL) when compared with that of the 7 days Bio-clean treated rats (Group 5: 27.67±1.32ng/mL) was found to be significantly lower (p=0.005). Lastly, the CD8 count of the positive control (Group 2: 18.92±2.22ng/mL) when compared with that of the negative control (Group 3: 35.72±1.47ng/mL) was found to be significantly lower (p=0.006).
Effect on Body Weight
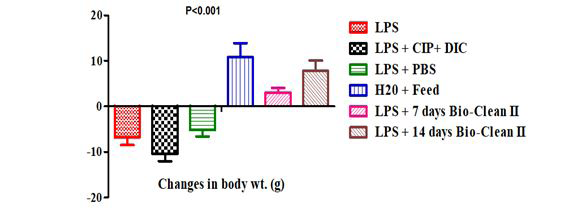
The change in weight of the control and test rats posttreatment with Bio-clean II is represented with a bar chart in Figure 3. The changes in body weight of the 7 days (Group 5: 3.00±1.05g) and 14 days (Group 6: 7.80±2.24g) Bio-clean treated rats were found to be significantly higher when compared to the inflammation control group (Group 1: -6.80±1.69g, p=0.029 and p=0.016, respectively), positive control group (Group 2: -10.40±1.63g, p=0.004, each) and negative control group (Group 3: -5.20±1.39g, p=0.028 and p=0.029, respectively). Furthermore, the variation in body weight of 7 days (Group 5: 3.00±1.05g) and 14 days (Group 6: 7.80±2.24g) Bio-clean treated rats was found to be non- significantly lower (p=0.636 and p=1.000, respectively) when compared to the zero control group (Group 4: 10.80±3.12g). Meanwhile, the changes in body weight of the 14 days Bio- clean treated rats (Group 6: 7.80±2.24g) was found to be non- significantly higher (p=0.806) than that of the 7 days Bio- clean treated rats (Group 5: 3.00±1.05g). The change in body weight of the positive control rats (Group 2: -10.40±1.63g) when compared with that of the negative control (Group 3: -5.20±1.39g) was found to be non-significantly lower (p=0.477).
Discussion
This current research was designed to assess the immunomodulatory effect of Bio-clean II on T-Helper 4 and Cytotoxic T-Lymphocytes in male Wistar rats exposed to purified Lipopolysaccharide® (from E. coli 0127: B8). The outcome of this study shows that Bio-clean II significantly increased CD4 Cell count, but significantly decreased CD8 Cell count when compared to the inflammation control in a duration-dependent manner.
This is in support of our previous study12 in which there was a significant (p<0.001) increase in the serum level of IL- 10 and a nonsignificant (p>0.05) decrease in the serum level of IFN-γ and TNF-α when compared to the inflammation control group in rats treated with Bio-Clean II post-exposure to purified LPS. It also agrees with the previous work of Ibeh IN, et al. [10] in which CD4 Cell count was significantly elevated in HIV positive patients treated with Bio-Clean II for 30days compared to the controls who received standard antiretroviral therapy. In this study, the herbal remedy Bio- clean II was shown to cause alterations in the levels of immune cells. The alteration is characterized by the increase in the initial depleted CD4+ Cells and decrease in the initial inflated CD8+ cells count in LPS-induced rats; thus demonstrating the immunomodulatory properties of the herbal remedy. Post-therapy, clinical cure of LPS-induced inflammation were defined as absence of signs and symptoms (including pain, swelling, twitching etc) in the treated rats compared to the inflammation control. The herbal remedy significantly improved and even cleared the symptoms of inflammation in the experimental animals.
The result of this present study is partly in agreement with that of a previous study by Ahmad W, et al. [38] in which the immunostimulatory effect of the extract of Tinospora crispa (a traditional medication with indications for several illnesses) on innate immune responses in Wistar Kyoto rats was demonstrated. Animals treated with extract of Tinospora crispa (21days) had enhanced expression of CD4+ (87.745±4.09Cells/μl), and CD8+ cytotoxic T-cell (49.345±2.76ng/mL) in comparison with the control (49.085±2.88Cells/μl, and 20.255±1.61ng/mL, respectively). The outcome of this study is also consistent with the work of Tatfeng and Enitan [39] who evaluated the effects of Allium cepa and Allium sativum on some immunological cells in rats. Treatment with onion and garlic extracts (28 days) significantly increased the CD4 cells Count (600±11Cells/ μl and 684±11Cells/μl, respectively) when compared to the zero control (200±11Cells/μl) which received no herbal treatment.
Furthermore, in comparison with an earlier study carried out by Sandikci M, et al. [40] on the effects of formaldehyde and xylene on CD4 and CD8 positive T cells in bronchus-associated lymphoid tissue in rats treated for 21 days, the experimental animals were challenged with formaldehyde and Xylene, elevated values of CD4 and CD8 cells were observed. While in this study, the CD4 counts of the inflammation control group (4.80±1.16Cells/μl) when compared to the test group (25.70 ± 2.39 Cells/μl) of the previous study were significantly increased. The case was not the same for the CD8 count which experienced non- significant depletion (32.93±2.52ng/mL), when compared to the test group (21.76 ± 0.94 ng/mL) of the previous study. CD8 T-cells respond to viral infections in general, but they also help to fight bacterial and protozoal diseases. New methods for reliably quantifying and characterizing pathogen-specific CD8 T-cells have been accessible in recent years, and our understanding of in vivo T-cell responses has greatly improved. Pathogen-specific T cells, formerly thought to be uncommon after infection, are now seen in high numbers, particularly in non-lymphoid tissues in the periphery. The complex link between microbial-induced innate inflammatory responses and the speed, size, and character of long-term T-cell responses is also highlighted by comparing CD8 T-cell responses to various infections [21].
Another observation worthy of mentioning is the effect of Bio-clean II on the weight of the Rats. There was a significant (p=0.000) increase in the weights of the rats administered with Bio-clean II for 7 and 14 days (3.00±1.05g and 7.80±2.24g, respectively), when compared to those treated with antibiotic combined with standard anti-inflammatory drug (-10.40±1.63g), as well as the inflammation control (-6.80±1.69g). Largely, the animals in the inflammation control, positive control, and negative control groups lost weight, while those in the zero control, 7 days, and 14 days Bio-clean treatment groups gained significant weight. This is consistent with the findings made by Ibeh IN, et al. [10, 11] who reported a significant increase in body weight of HIV positive patients treated with Bio-Clean, suggesting the weight gaining potential of the herbal remedy in immunosuppressed and malnourished individuals.
Conclusion
Bio-Clean Il caused slight alterations in the CD4 and CD8 counts of rats exposed to purified lipopolysaccharide. The outcome of this study, underscores the potential role of Bio- Clean Il in the treatment of bacterial inflammatory diseases if attention should be given to duration of therapy. The outcome of this present study gave further credence to the immunomodulatory potential of Bio-Clean II in LPS-induced inflammation. Moreover, since the immunomodulatory effect of the herbal remedy (especially at 14-day duration therapy) was comparable to that of ciprofloxacin (a conventional antibiotic for treating bacterial infection) and diclofenac (anti-inflammatory drug) combined, in view of the continuous emergence of antibiotic resistance, the herbal remedy may therefore serve as a better alternative for the treatment of LPS-induced inflammation; which is both cheap and readily available especially in resource-limited settings with little or no access to healthcare services. However, the signaling pathways involved in the immunomodulatory potential of the herbal remedy would require further investigation.
Acknowledgments
All authors made substantial contributions to this study. We are grateful to the Staff of Babcock University Animal House involved in this research.
Disclosure
The authors report no conflicts of interest in this work.
Ethical Approval
Ethical approval for this animal experimental study was granted by the Babcock University Health Research Ethics Committee (BUHREC) with ethical approval registration number: BUHREC 513/21. Adherence to Animal Care Regulations and Standards The animal research adheres to the ARRIVE guidelines (https://arriveguidelines.org/arrive-guidelines).
Data Availability
The authors shall make available the data underlying this study on request.
Funding
This study was self-funded. No financial support was received from any funding agency.
References
-
Balick MJ, Paul AC (1996) Plants that heal. In their plants, people, and culture; the science of ethnobotany. Scientific American Library, New York, pp: 25-61.
-
(2001) Legal Status of Traditional Medicine and Complementary/ Alternative Medicine: Worldwide Review. World Health Organization.
-
(2008) Herbal Medicine. World Health Organization.
-
Shulz V, Rudolf H, Mark B, Tyler VE (2001) Medicinal plants, Phytomedicines and Phytotherapy. 4th (Edn), In their rational phytotherapy; a physician’s guide to herbal medicine. Berlin, New York, Springer, pp: 1-42.
-
Enitan SS, Olley M, Uhunmwangho SE, Akele YR, Ehiaghe AF, et al. (2014) Antibacterial Activity of Methanolic Leaf Extract of Plukenetia conophora Mull. arg. against Selected Bacteria Isolated from Urinary Tract Infection, International Journal of Microbiology and Application, pp: 1-10.
-
Ngcobo M, Gqaleni N (2016) Evaluation of the immunomodulatory effects of a South African commercial traditional immune booster in human peripheral blood mononuclear cells. BMC Complement Altern Med 16(1): 300.
-
Maqbool M, Dar MA, Gani I, Mir SA, Khan M (2019) Herbal Medicines as an Alternative Source of Therapy: A Review. World Journal of Pharmacy and Pharmaceutical Sciences 8(2): 374-380.
-
Wu D, Lewis ED, Pae M, Meydani SN (2019) Nutritional Modulation of Immune Function: Analysis of Evidence, Mechanisms, and Clinical Relevance. Front Immunol 9: 3160.
-
Enitan SS, Ibeh IN, Akele RY, Isitua CC, Idris PO (2022) Phytochemical Screening, Trace Element Analysis, Bacteriological Quality and Efficacy of Bio-Clean II Herbal Remedy. Traditional Medicine and Modern Medicine.
-
Ibeh IN, Akanu N, Mkpa AM, Isitua CC, Ogefere HO, et al. (2013) Evaluation of the Anti-Human Immunodeficiency Virus (HIV) Properties of DXL (Decoction X-Liquid- Bioclean li). J Clin Toxicol 12(4): 1-7.
-
Ibeh IN, Okungbowa MA, Ibeh NI, Adejumo BI (2016) Comparative studies on the effects of Zidovudine Nevirapine Lamivudine and Bio-clean II on female HIV/ AIDS cases in Nigeria. JAMPS 9(3): 1-7.
-
Enitan SS, Ibeh IN, Akele RY, Isitua CC, Jegede OO, et al. (2022) Immunomodulatory Effects of Bio-Clean II on some Inflammatory Cytokines in Rats Exposed to Purified Bacterial Lipopolysaccharide. Immunotoxicology.
-
Ibeh IN, Okungbowa MA, Ibeh NI, Isitua CC (2016) Studies on HIV Resistance to Multiple Drug Therapy (Lamivudine, Zidovudine and Nevirapine) in Benin Metropolis, Nigeria. International Journal of Scientific Research in Environmental Science and Toxicology 1(1): 1-3.
-
De Castro C, Antonio M (2010) Methods in Enzymology Book series. In: Fukuda M, et al. (Eds.), pp: 2-581.
-
Mayer H, Tharanathan RN, Weckesser J (1985) Analysis of Lipopolysaccharides of Gram-negative bacteria. Meth Microbiol 18: 157-207.
-
Levi M, Keller TT, Van-Gorp E, Ten-Cate H (2003) Infection and inflammation and the coagulation system. Cardiovas Res 60(1): 26-39.
-
Ferrero-Miliani L, Nielsen OH, Andersen PS, Girardin SE (2007) Chronic inflammation: importance of NOD2 and NALP3 in interleukin-1beta generation. Clin Exp Immunol 147(2): 227-235.
-
Abbas AB, Lichtman AH (2009) Chapter 2 Innate Immunity. In: Saunders (Eds.), Basic Immunology. Functions and disorders of the immune system. 3rd (Edn), Elsevier, UK, pp: 239-243.
-
Dube MP, Sattler FR (2010) Inflammation and complication of HIV disease. Journal of Infectious disease 201(12): 1783-1785.
-
Wong P, Pamer EG (2003) CD8 T cell responses to infectious pathogens. Annu Rev Immunol 21(1): 29-70.
-
Khan S, Malik F, Suri KA, Singh J (2009) Molecular insight into the immune up-regulatory properties of the leaf extract of Ashwagandha and identification of Th1 immunostimulatory chemical entity. Vaccine 27(43): 6080-6087.
-
Kassim M (2009) Immune Regulation. Essential Clinical Immunology. United States of America by Cambridge University Press, New York, pp: 33-44.
-
Lederman S, Yellin MJ, Krichevsky A, Belko J, Lee JJ, et al. (1992) Identification of a novel surface protein on activated CD4+ T cells that induces contact-dependent B cell differentiation (help). J Exp Med 175(4): 1091-1101.
-
Luckheeram RV, Zhou R, Verma AD, Xia B (2012) CD4+T Cells: Differentiation and Functions. Clin Dev Immunol 2012: 925135.
-
Hariyanto AD, Permata TBM, Gondhowiardjo SA (2021) Role of CD4+CD25+FOXP3+ TReg cells on tumor immunity. Immunol Med, pp: 1-14.
-
Delves PJ, Martin SJ, Burton DR, Roitt IM (2011) Lymphocyte activation. Roitt’s Essential Immunology. 12th (Edn.), Wiley, West Sussex, UK, pp: 560.
-
Lees CJ, Apostolopoulos V, McKenzie IF (1999) Cytokine production from murine CD4 and CD8 cells after mannan-MUC1 immunization. J Interferon Cytokine Res 19(12): 1373-1379.
-
Al-Shura AN (2020) Lymphocytes. Advanced Hematology in Integrated Cardiovascular Chinese Medicine. Elsevier, pp: 41-46.
-
Valentine M, Song K, Maresh GA, Mack H, Huaman MC, et al. (2013) Expression of the Memory Marker CD45RO on Helper T Cells in Macaques. PLoS ONE 8(9): e73969.
-
Subramanian S, Ramalingam K (2005) Electron microscopic evidence on the participation of Cytotoxic T Lymphocytes and Macrophages in Mtb adjuvant induced connective tissue inflammation and arthritogenesis in Rattus norvegicus. Asian Journal of Microbiology, Biotechnology and Environmental Sciences 7(2): 227- 233.
-
Carvalheiro H, da Silva JAP, Souto-Carneiro MM (2013) Potential roles for CD8+ T cells in rheumatoid arthritis. Autoimmun Rev 12(3): 401-409.
-
McAleer JP, Vella AT (2008) Understanding how lipopolysaccharide impacts CD4 T-cell immunity. Crit rev immunol 28(4): 281-299.
-
(1996) Guide for the care and use of laboratory animals. Institute for Laboratory Animal Research Council, Division on Earth and Life Studies, American Academy of Sciences, National Research Council of the National Academies, The National Academies Press 500 Fifth Street, NW Washington 11(8): 131.
-
Ochei J, Kolhatkar A (2006) Euthanasia of Animals by Cervical Dislocation. In: Ochei J et al. (Eds.), Theory and Practice of Medical Laboratory Science. Tata McGraw- Hill publishing Company Limited, New Delhi, pp: 1213- 1230.
-
Moon S, Gurkan UA, Blander J, Fawzi WW, Aboud S, et al. (2011) Enumeration of CD4+ T-Cells Using a Portable Microchip Count Platform in Tanzanian HIV-Infected Patients. PLoS ONE 6(7): e21409.
-
Kannangai R, Prakash KJ, Ramalingam S, Abraham OC, Mathews KP, et al. (2000) Peripheral CD4+/CD8+ T-lymphocyte counts estimated by an immunocapture method in the normal healthy south Indian adults and HIV seropositive individuals. J Clin Virol 17(2): 101-118.
-
Shott S (1990) Statistics for health professionals. Saunders WB, Co Philadelphia, pp: 313-336.
-
Ahmad W, Jantan I, Kumolosasi E, Bukhari SN (2015) Immunostimulatory effects of the standardized extract of Tinospora crispa on innate immune responses in Wistar Kyoto rats. Drug Des Devel Ther 9: 2961-2973.
-
Tatfeng YM, Enitan SS (2012) Effect of Allium cepa and Allium sativum on some immunological cells in rats. Afr J Tradit Complement Altern Med 9(3): 374-379.
-
Sandikci M, Eren U, Kum S (2007) Effects of formaldehyde and xylene on CD4- and CD8-positive T cells in bronchus- associated lymphoid tissue in rats. Toxicol Ind Health 23(8): 471-477.
- Are the Vaccines the Only Solution to Prevent the COVID-19 Pandemic? Part Two
- Clinical Characteristics of Women in this New Global Immunodeficiency
- Cell Dynamics in HIV Pathogenesis: Insights and Implications
- Determination of the CDR (CDR1, CDR2) « Complementary- Determining Region Invertebrate Primitive Antibody from Sea Star »
- Prioritizing Care for High-Risk COVID-19 Patients in the EU: 10 Civic Recommendations to the Institutions
- Comprehensive Insights into ModRNA Vaccines: Persistent PP-Spike Recombinant Protein, Hyperimmune/Inflammatory Reactions, Thrombotic Vasculopathy, Chronic Organ Complications and Excess Deaths