Chimeric Antigen Receptor T-Cell Therapy (Car T-Cells) in Solid Tumors, Resistance and Success
CARs are chimeric synthetic antigen receptors that can be introduced into an immune cell to retarget its cytotoxicity toward a specific tumor antigen. CAR T-cells immunotherapy demonstrated significant success in the management of hematologic malignancies. Nevertheless, limited studies are present regarding its efficacy in solid and refractory tumors. It is well known that the major concerns regarding this technique include the risk of relapse and the resistance of tumor cells, in addition to high expenses and limited affordability. Several factors play a crucial role in improving the efficacy of immunotherapy, including tumor mutation burden (TMB), microsatellite instability (MSI), loss of heterozygosity (LOH), the APOBEC Protein Family, tumor microenvironment (TMI), and epigenetics. In this minireview, we address the current and future applications of CAR T-Cells against solid tumors and their measure for factors of resistance and success.
Introduction
Revolutionary discoveries have been made regarding cancer treatment modalities. Chimeric antigen receptor (CAR) T-cell therapy is a challenging method that uses re-engineered lymphocytes expressing specific modular synthetic surface antibodies. The CARs are encoded by specific genes and inserted into the genome of T-cells, which are either autologous or derived from allogeneic donors.
The outcome of obtaining effective autologous T cells from the patient is suboptimal due to the quality and quantity of the harvested cells in addition to being time-consuming and unprofitable.
On the other hand, using allogeneic T cells presents several challenges due to the presence of endogenous Major Histocompatibility Class I (Human Leukocyte Antigen HLA barriers) and T-cell receptors (TCR) on donor’s T lymphocytes, which lead to alloreactivity and graft-versus-host disease (GVHD). Therefore, to overcome the aforementioned HLA barriers, advanced technology was applied to produce modified CAR T-cells by knocking out TCRs and HLA-I in allogeneic T cells. The CARs are separately generated on a case-by-case basis to achieve high personalized effectiveness against tumors [1, 2, 3].
Indicators of Improved Response to CAR T-Cell and Immunotherapy
- The presence of a high extent of tumor-infiltrating lymphocytes (TILs).
- In breast cancer, the quantity and composition of TILs play a major role in determining the response to therapy [4].
- Expression of immune microenvironment markers that affect prognosis and treatment response in several cancer subtypes. For example, the expression of HER2, KI67, PIK3CA, P53, PD1, PDL1, and CTLA4 in breast cancer indicate promising targets for adjuvant therapies [5, 6].
- Detection of other circulating biomarkers including lactate dehydrogenase (LDH), which is associated with poor prognosis in solid neoplasms, particularly in breast, melanoma, prostate, and renal cell carcinomas [7].
- Elevated neutrophil-to-lymphocyte (N/L) ratio or high expression of tumor-associated neutrophils (TAN) could be related to worse prognosis in multiple solid tumors including prostate, bladder, and nasopharyngeal carcinomas. Nevertheless, results are still limited regarding other types of neoplasms [8].
- High tumor mutational burden (TMB) due to an increased level of nonsynonymous somatic mutations, which revealed an improved response to CAR T-cell therapy combined with depletion of tumor cells by chemo or radiation therapy. Therefore, a low tumor mutation burden might worsen the response to immunotherapy despite the expression of immune checkpoint genes [9, 10].
- Homologous recombination repair (HR) deficiency increases genomic instability in tumors and subsequently improves tumors immunogenicity and response to CAR T-cell therapy [11].
Genome Engineering Editing Technologies That Produce CAR T-Cells
Zinc-finger nucleases (ZFNs): ZFNs have been used to modify endogenous genes in a variety of cell types. Genomic alterations including insertions, duplications, and point mutations could be introduced based on this method. ZFNs are used to obstruct gene expression of TCR β and α chain with an increased level of highly-specific cell surface exogenous TCR [12].
Transcription activator-like effector nucleases (TALENs): TALENs are defined as a non-specific DNA-cleaving nuclease connected to a DNA-binding domain. This technique enables modifying any sequence of interest in living cells or organisms. Recently, TALENs have rapidly emerged as an alternative method to ZFNs in introducing targeted double- strand breaks (DSBs) and genome editing [13].
Lentiviral vectors: Another possible technique involves transducing CARs into T cells via genetically engineered lentiviruses that represent viral vectors with their ability to infect non-dividing cells [14].
Clustered regulatory interspaced short palindromic repeats/ CAS9: (CRISPR) is a novel genome editing technique that exhibits the insertion of CARs at precise target regions of the genome with relative ease. This technique revealed a robust antitumor activity upon knocking out beta- 2-microglobulin (β2M), in addition to PDCD1 and TRAC, which aid in preventing GVHD in the leukemia mice model [12, 15, 16].
Cas-cLOVER: Cas-CLOVER is a novel gene-editing technology alternative to CRISPR/Cas9, demonstrating high fidelity with no detectable off-targets while maintaining robust editing efficiency. It is functionally similar to CRISPR/Cas9 technologies but uses a different nuclease protein called Clo51 fused to a nuclease-inactivated Cas9 protein.
Cas-CLOVER may represent a combination of the CRISPR–Cas9 and TALEN as applied in deletion of αβTCR and β2M that has been successfully achieved.
Cas-CLOVER revealed greater specificity through the utilization of two guide RNAs as well as a nuclease activity that requires dimerization of subunits associated with each guide RNA [17, 18, 19].
Resistance to CAR T-Cell Therapy
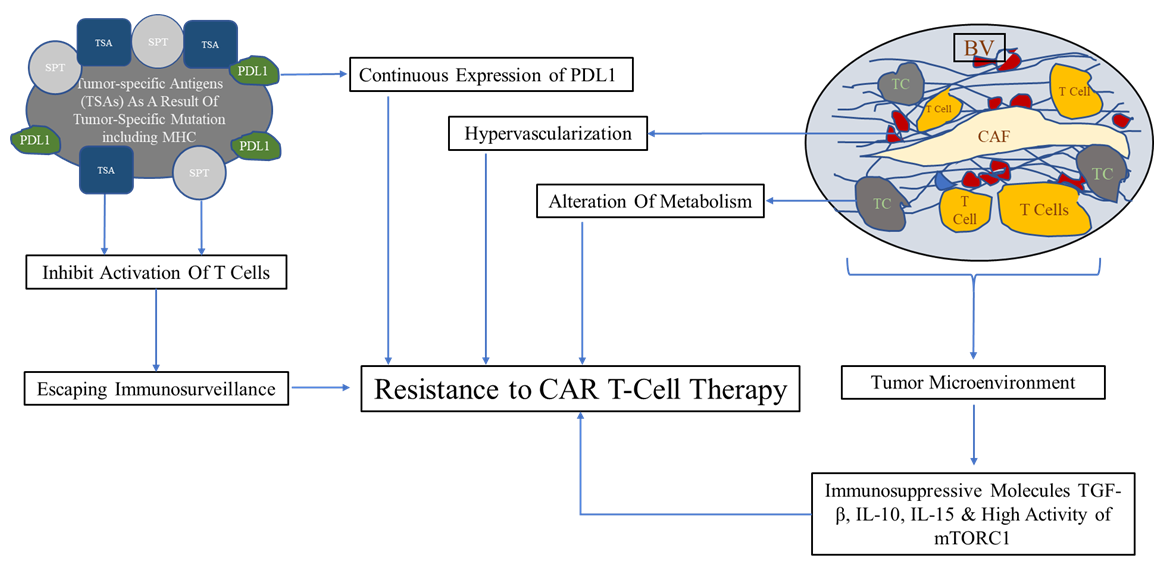
Major challenge for CAR T-Cell application is the resistance and the side effects to therapy Figure 1
- Constant modification of surface proteins of tumor cells facilitates their ability to escape from the immune system [20].
- Abnormal expression of tumor-specific antigens on the surface of tumor cells. These antigens are presented by MHC class I or II molecules on the surface of tumor cells. They are called tumor-specific antigens (TSAs) and typically result from a tumor-specific mutation [20].
- Alterations in the signaling of antigen-presenting pathways, including mutations interfering with the proteasome, and modifications in the structure of MHC. These changes inhibit the correct activation of T cells and encourage escaping immunosurveillance by tumor cells. In addition, evolving of new mutations in tumor cells themselves represent an additional factor in increasing resistance to therapy [21].
- Tumor microenvironment, including cancer-associated fibroblasts CAFs, fibroblast, regulatory T cells, extracellular collagen matrix, and blood vascular network. These factors facilitate tumor growth, invasion, and relapse after therapy. Also, alteration of metabolism in tumor cells as a response to the environmental metabolic changes represent an additional factor [21].
- Immature vascularization in tumors promotes the expression of hypoxia-inducible factor-1 (HIF-1), and subsequently induces the expression of vascular endothelial growth factor (VEGF), epidermal growth factor, fibroblast growth factor (FGF), hepatocyte growth factor (HGF), platelet-derived growth factor (PDGF), and other angiogenic proteins. This cascade stimulates hypervascularization that contributes to resistance to immunotherapy [22].
- Exhaustion of T cells due to continuous expression of PDL1 by tumor cells and the severity of exhaustion correlated with the level of antigen stimulation. Interestingly, epigenomic alteration is most likely result in exhaustion phenotype in T cells [23].
- Secretion of immunosuppressive molecules including TGF-β, IL-10, in addition to the increase in mTORC1 activity mediated by IL15 which participates in resistance to therapy [9, 24].
CAR T-Cells and Loss of Heterozygosity (LOH)
The link between CAR T Cells therapy and LOH was not studied long enough in solid tumors, however, irreversible loss of heterozygosity in CD19 on Chromosome 16q could be used as a biomarker for an outcome prediction after the CAR T cells therapy for B cell acute lymphoblastic leukemia (B-ALL) [25].
CAR T-Cells and Microsatellite Instability (MSI)
Studies on patients with gastric carcinoma who expressed high MSI were found to have higher expression levels of MET gene than normal tissue, and this result provided strong support for the selection of c-Met as the target of CAR-T.
However, the presence of the immunosuppressive microenvironment will reduce the efficacy of CAR-T in solid tumors. The PD1/CD28 chimeric-switch receptor (CSR) improves immunosuppression by fusing the extracellular domain of PD-1 with the transmembrane and intracellular domains of CD28, thus transforming the inhibitory signal of PD-1 into the activation signal of CD28. Therefore, CAR- Ts with PD1/CD28 CSR had a better anti-tumor effect than CAR-T combined with PD-1 antibody alone [26].
CAR-T cells and APOBAEC
The APOBEC (Apolipoprotein B mRNA Editing Catalytic Polypeptide-like) is a family of proteins that are characterized by their ability to bind to RNA and single-stranded DNA. Several factors regulate their activity, including genetic alterations, changes in transcription, and interactions with intracellular macromolecules. Loss of cellular control of APOBEC activities is associated with cancer progression and treatment resistance [27, 28].
CAR-T cells and Immune Checkpoint Inhibitors
Cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) and programmed death1(PD-1) are defined as checkpoint receptors that are expressed by activated T cells to negatively control their cytotoxic activities. Overexpression of these receptors leads to excessive activation and exhaustion of T-cells. In contrast, blocking and inhibition of these products improve the effect of CAR T cells [29].
CAR T Cells and Bispecific Antibody
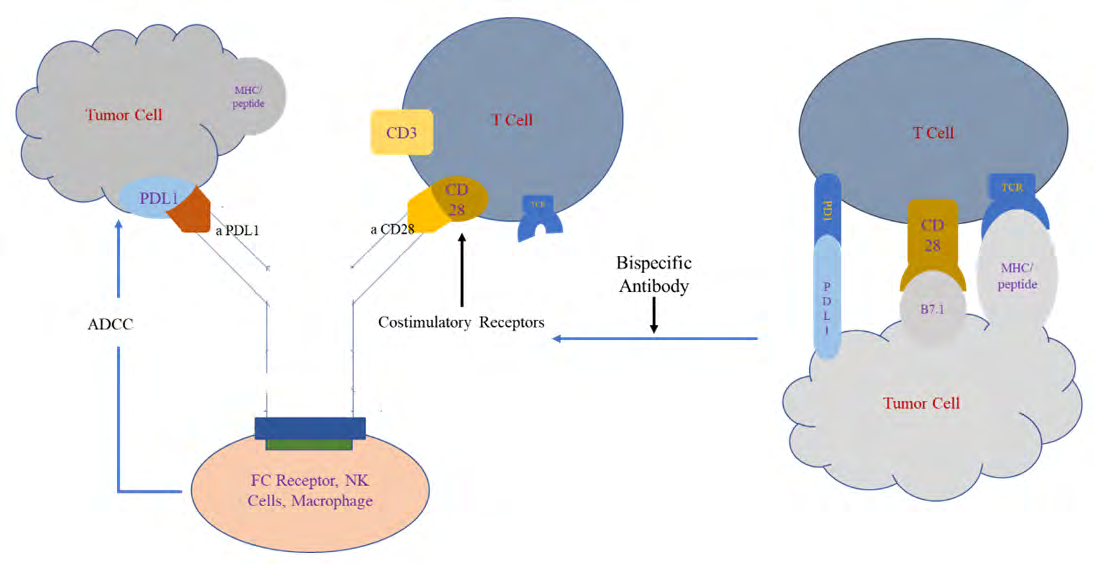
Because of several mechanisms of Cart T Cells resistance, recently, a new approach has been proposed and advanced to overcome such resistance by producing Bispecific Antibody Armed T cells (BATs), which revealed encouraging clinical results in breast, prostate, and pancreatic cancer [30].
The bispecific monoclonal antibody (BsMAb, BsAb) is an artificial protein that can simultaneously bind to two different antigens or two different epitopes on the same antigen [31].
bispecific antibodies can be engineered by nonbinding two epitopes of different antibodies such as, anti-HER2 and CD3, anti-HER2/HER3, anti-CD19/CD3, EpCAM/CD3TF2, CD28/PDL1, and Her2/PDL1 (Figure 2) [32].
In one study, BsMAb targets CD28 homolog (CD28H) (a newly identified B7 family receptor), which is constitutively expressed on T and natural killer (NK) cells with a PD- L1 receptors on tumor cells to potentiate tumor-specific immune responses.
The best CD28H–PD-L1 bispecific antibody which retained equivalent binding of both antigens to their parental IgG, (anti-CD28H) as the Fab and (anti–PD-L1) as an scFv appended to the C-terminus of the human IgG4P Fc domain.
On the other hand, CD28H–PD-L1 bispecific antibody perform more functions than the two separate antibodies (anti PDL1 and anti CD28H) because binding in one arm to the PDL-1 receptor results in T-cell costimulation, inducing NK-cell cytotoxicity of PD-L1–expressing tumor cells and activated tissue-resident memory CD8+ T cells that may lead to induction of durable, therapeutic antitumor responses.
Mechanistically, the CD28H agonistic arm of the bispecific antibody reduced PD-L1/PD-1–induced SHP2 phosphorylation while simultaneously augmenting T-cell receptor signaling by activating the MAPK, AKT and MTOR pathways [33].
CAR T-cells and Epigenetic Factor
Despite the high preservation of DNA methylation patterns, several studies highlighted a significant association between epigenetic factors and various cancer subtypes, which present epigenetics as a major determiner in fetal cell development and differentiation.
The chromatin might represent a signaling platform to integrate signals to control gene expression. These changes are demonstrated by different chromatin appearances characterized by histone post-translational modifications and DNA modifications, which affect the gene expression pattern in T cells. Subsequently, epigenetic modifications could ultimately improve the efficacy of CAR-T cell therapy [34]. However, the epigenetic regulation (DNA methylation and histone modifications) plays an important role in the differentiation of T cells.
It has been reported that acquired Cancer Cell Resistance to T cell bispecific antibodies and car T targeting her2 in Vivo was conducted through Jak Down-Modulation and Due to Epigenetic Modification as well [35].
Conclusion
CAR-T cell therapy is a challenging method that represents a promising tool in assessing better outcomes in cancer patients. This technique faces several challenges in solid tumors, including tumor mutational burden and tumor microenvironment. In our manuscript, we aimed to present challenges, innovations, and improvements to this revolutionary treatment method.
References
-
Wilkins O, Keeler AM, Flotte TR (2017) CAR T-Cell Therapy: Progress and Prospects. Hum Gene Ther Methods 28(2): 61-66.
-
Filley AC, Henriquez M, Dey M (2018) CART Immunotherapy: Development, Success, and Translation to Malignant Gliomas and Other Solid Tumors. Front Oncol 8: 453.
-
Wang X, Rivière I (2016) Clinical manufacturing of CAR T cells: foundation of a promising therapy. Mol Ther Oncolytics 3: 16015.
-
Gatti-Mays ME, Balko JM, Gameiro SR, Bear HD, Prabhakaran S, et al. (2019) If we build it they will come: targeting the immune response to breast cancer. NPJ Breast Cancer 5: 37.
-
Hendricks WPD, Briones N, Halperin RF, Facista S, Heatonet PR, et al. (2019) PD-1-Associated Gene Expression Signature of Neoadjuvant Trastuzumab- Treated Tumors Correlates with Patient Survival in HER2-Positive Breast Cancer. Cancers (Basel) 11(10): 1566.
-
Buchbinder EI, Desai A (2016) CTLA-4 and PD-1 Pathways: Similarities, Differences, and Implications of Their Inhibition. Am J Clin Oncol 39(1): 98-106.
-
Zhang J, Yao YH, Li BG, Yang Q, Zhang PY, et al. (2015) Prognostic value of pretreatment serum lactate dehydrogenase level in patients with solid tumors: a systematic review and meta-analysis. Sci Rep 5: 9800.
-
Cupp MA, Cariolou M, Tzoulaki I, Aune D, Evangelou E, et al. (2020) Neutrophil to lymphocyte ratio and cancer prognosis: an umbrella review of systematic reviews and meta-analyses of observational studies. BMC Med 18(1): 360.
-
Yadav RK, Ali A, Kumar S, Sharma A, Baghchi B, et al. (2020) CAR T cell therapy: newer approaches to counter resistance and cost. Heliyon 6(4): e03779.
-
Hu FF, Liu CJ, Liu LL, Zhang Q, Guo AY (2021) Expression profile of immune checkpoint genes and their roles in predicting immunotherapy response. Brief Bioinform 22(3).
-
van Wilpe S, Tolmeijer SH, Koornstra RHT, de Vries IJM, Gerritsen WR, et al. (2021) Homologous Recombination Repair Deficiency and Implications for Tumor Immunogenicity. Cancers (Basel) 13(9): 2249.
-
Azangou-Khyavy M, Ghasemi M, Khanali J, Boroomand- Saboor M, Jamalkhah M, et al. (2020) CRISPR/Cas: From Tumor Gene Editing to T Cell-Based Immunotherapy of Cancer. Front Immunol 11: 2062.
-
Joung JK, Sander JD (2013) TALENs: a widely applicable technology for targeted genome editing. Nat Rev Mol Cell Biol 14(1): 49-55.
-
Labbé RP, Vessillier S, Rafiq QA (2021) Lentiviral Vectors for T Cell Engineering: Clinical Applications, Bioprocessing and Future Perspectives. Viruses 13(8): 1528.
-
Kato-Inui T, Takahashi G, Hsu S, Miyaoka Y (2018) Clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 with improved proof-reading enhances homology-directed repair. Nucleic Acids Res 46(9): 4677-4688.
-
Ren J, Liu X, Fang C, Jiang S, June CH, et al. (2017) Multiplex genome editing to generate universal CAR T cells resistant to PD1 inhibition. Clin Cancer Res 23(9): 2255-2266.
-
Rafiq S, Hackett CS, Brentjens RJ (2020) Engineering strategies to overcome the current roadblocks in CAR T cell therapy. Nat Rev Clin Oncol 17(3): 147-167.
-
Barnett B (2018) Manufacture of allogeneic Universal Donor CAR-T therapies using piggyBac™ and Cas- CLOVER in CAR-TCR Summit, Boston MA.
-
Cas-CLOVER Nucleases – The Clean Alternative To CRISPR/Cas9.
-
Vina y DS, Ryan EP, Pawelec G, Talib WH, Stagg J, et al. (2015) Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin Cancer Biol 35(Suppl): S185-S198.
-
Bai R, Chen N, Li L, Du N, Bai L, et al. (2020) Mechanisms of Cancer Resistance to Immunotherapy. Front Oncol 10: 1290.
-
Katayama Y, Uchino J, Chihara Y, Tamiya N, Kaneko Y, et al. (2019) Tumor Neovascularization and Developments in Therapeutics. Cancers (Basel) 11(3): 316.
-
Wherry EJ, Kurachi M (2015) Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol (8): 486-499.
-
Xia A, Zhang Y, Xu J, Yin T, Lu XJ (2019) T Cell Dysfunction in Cancer Immunity and Immunotherapy. Front Immunol 10: 1719.
-
Zhang X, Sjöblom T (2021) Targeting Loss of Heterozygosity: A Novel Paradigm for Cancer Therapy. Pharmaceuticals (Basel) 14(1): 57.
-
Chen C, Gu YM, Zhang F, Zhang ZC, Zhang YT, et al. (2021) Construction of PD1/CD28 chimeric-switch receptor enhances anti-tumor ability of c-Met CAR-T in gastric cancer. Oncoimmunology 10(1): 1901434.
-
Salter JD, Bennett RP, Smith HC (2016) The APOBEC Protein Family: United by Structure, Divergent in Function. Trends Biochem Sci 41(7): 578-594.
-
Georgiadis C, Rasaiyaah J, Gkazi SA, Preece R, Etuk A, et al. (2021) Base-edited CAR T cells for combinational therapy against T cell malignancies. Leukemia 35(12): 3466-3481.
-
Shen SH, Woroniecka K, Barbour AB, Fecci PE, Sanchez- Perez L, et al. (2020) CAR T cells and checkpoint inhibition for the treatment of glioblastoma. Expert Opin Biol Ther 20(6): 579-591.
-
Thakur A, Scholler J, Kubicka E, Bliemeister ET, Schalk DL, et al. (2021) Bispecific Antibody Armed Metabolically Enhanced Headless CAR T Cells. Front Immunol 12: 690437.
-
Bispecific monoclonal antibody.
-
Corsiero E, Colombo F (2021) Bispecific Antibodies and Trispecific Antibodies. Queen Mary University of London, London, England.
-
Ramaswamy M, Kim T, Jones DC, Ghadially H, Mahmoud TI, et al. (2022) Immunomodulation of T- and NK-cell Responses by a Bispecific Antibody Targeting CD28 Homolog and PD-L1. Cancer Immunol Res 10 (2): 200- 214.
-
Akbari B, Ghahri-Saremi N, Soltantoyeh T, Hadjati J, Ghassemi S, et al. (2021) Epigenetic strategies to boost CAR T cell therapy. Mol Ther 29(9): 2640-2659.
-
Arenas EJ, Sabadell AM, Ruiz IR, Alonso MR, Escorihuela M, et al. (2021) Acquired cancer cell resistance to T cell bispecific antibodies and CAR T targeting HER2 through JAK2 down-modulation. Nature Communications 12(1): 1237.
- Effects of 5-HTP and Melatonin on the Sleep Cycle of Medical Students
- Adsorption of Bisphenol A on NH4OH- Modified Rice Husk and Sugar Cane Bagasse Biochar
- Comparative Assessment of the Reinforcement Efficiency of Palm Fruit Fibre and Coconut Fibre in High Density Polyethylene (HDPE) Matrix Composite
- Importance of Bio Compounds Naturally Present in Food with Functionality in Animal Metabolism
- Sub-Acute Study on the Cardiotoxic Effects of Monosodium Glutamate Ingestion in Albino Rat
- Weight Management and Its Natural Solutions: A Review