Molecular Strategies for Development of Abiotic Stress Tolerance in Plants
<p>It is an established fact that abiotic stresses are a major cause of productivity losses worldwide. Abiotic stresses not only reduce crop yield but also limit the area that can be brought under productive cultivation. Global warming has only added to this already grim scenario. Development of novel, stress resistant genotypes through transgenic technology is seen as a viable solution to this critical problem. However, different abiotic stresses have distinctive primary effects on plants, while all of them have a common denominator in the form of production of reactive oxygen species. Therefore, understanding the biochemical and molecular basis of differential abiotic stress effectors is critically important for the development of transgenic crops. Further, novel gene discovery, annotation and validation is an integral part of this process. The current review highlights various strategies that have been employed to develop transgenic crops resistant to specific abiotic stresses, along with exploring the use of nanotechnology in developing stress tolerance in plants.</p>
Introduction
Plants being sessile in nature are constantly challenged by various environmental stresses. Stress may be defined as an environmental condition that can potentially reduce the growth and consequently the yield of a plant. Environmental stresses include both biotic as well as abiotic stress. Abiotic stress factors are often more damaging than the biotic stresses and are naturally occurring, often intangible, factors such as sunlight, wind, water, nutrients etc. These factors are a major impediment for achieving food and nutritional security. The human population is expected to reach the nine billion mark by 2050 [1]. The seemingly explosive increase in human population is limiting land and other resources for agriculture. Anthropogenic activities are leading to land degradation and cultivation is shifting to more marginal areas and soil types. Additionally, climate change has exacerbated the conditions for agriculture worldwide, with significant yield reductions reported in major cereal species such as wheat, maize, and barley [2]. On top of it, due to global warming in many parts of the world, rainfall has become less predictable, more intense, leading to recurring floods. Further, a continual rise in global temperature has increased the during of dry periods, thereby increasing the chances of drought. For example, in the major crop growing areas of eastern Africa, the predominant rainy season is starting later and ending earlier [3] with longer dry spells in between [4]. The growing food demand and the threat of heavy crop losses due to global climate change, is a compelling factor pushing us to look for viable strategies to substantially Cell & Cellular Life Sciences Journal
improve food availability, thus making stress resistant crops a dire need.
Plant’s Response to Abiotic Stress
It is an established fact that abiotic stress is a major constraint for crop production and productivity. Abiotic stresses such as extreme temperatures, low water availability, high salt concentration and mineral deficiencies or metal toxicities severely diminish productivity of cash crops. This increase in the quantum of environmental stresses poses a serious threat to global agricultural production and food security. It is suggested that approximately 50% of yield reduction is the direct result of abiotic stresses [5]. Despite the presence of large genetic diversity amongst the important food crops, approximately two-third of the maximal yield potential of major crops is lost due to unfavorable growth conditions [6]. The overall effect of abiotic stresses on the yield component of crops is mainly dependent on the magnitude and duration of the imposed stress. The main abiotic stresses, posing threat to the world agricultural output are drought and salinity. Inadequate availability of quality irrigation water, and salt stress caused due to various soil components or the quality of irrigated water, lead to reduction in plant growth and productivity. Depending upon the magnitude of the external stress, plants try to adapt to the environmental conditions, through various morphological, physiological and biochemical changes. Adaptive responses to abiotic stress include closure of stomata which limits water loss and initiation of a series of physiological processes which maintain the integrity of the photosynthetic and carbon fixation apparatus, activation of the antioxidant defense system in the plant etc. Although most of the biochemical factors necessary for stress tolerance acquisition are present in all species, but the degree or strength of the response varies from species to species. The difference in the plant’s response to the imposed stress depends on its ability for signal perception, transduction and potential of the defense machinery to respond to these signals.
Approaches for Abiotic Stress Tolerance
Various approaches (Figure 1) have long been employed to develop stress tolerant varieties such as conventional breeding and molecular breeding approaches; but due to limitations such as restricted availability of germplasm, intensive labor requirements, extended time and resource requirements, these approaches have not yielded the desired results. In recent decades, molecular approaches have taken a quantum jump, in devising strategies for the development of new germplasm. One of the most promising and practical approaches for development of new stress tolerant varieties is genetic engineering. Through the use of transgenic technology, one can produce plants with desired traits such as tolerance to various abiotic stresses that includes water stress (flood and drought), temperature stress (high and low), and salt stress more precisely. Development of plant transformation technology is impacting crop improvement options in unprecedented ways. Different stresses elicit differential response in plants, and hence each kind of stress has its own set of responsive genes. Although, several common genes may be participate in response to different abiotic stresses, yet each type of stress also induces its characteristic gene expression profile. The key to the tolerance of plants to abiotic stresses is a complex network of transcription factors and other regulatory genes that control multiple defense enzymes, proteins, and pathways [6, 7].
Figure 1: Techniques employed for developing stress tolerant plants. Apart from recombinant DNA approaches, nanotechnological interventions have also shown promising results in the recent past. As of now nanotechnology is relatively nascent, but nevertheless it has shown strong potential in solving the problem of abiotic stresses. Nanotechnological interventions are currently being explored and tested in selected plant species, and there are also concerns about the environmental safety aspects of this technology. However, any new technology is like a double edged sword and we have to fine tune the technology to reap maximum benefit from it. Specifically in agriculture, technical innovation is essentially required to solve the global challenges such as Cell & Cellular Life Sciences Journal
producing food for the burgeoning population, climate change, declining nutrient use efficiency etc. Nanotechnology applied to food and agricultural sector can be a real game changer, in terms of harnessing the untapped potential of the current breed of crop plants. In the recent past, engineered nanoparticles have received particular attention as potential candidates for improving crop yield [8, 9, 10]. As of now, nanotechnology has the potential to bring in a nano-revolution, in the field of food and agriculture, although as mentioned earlier a few issues are needed to be addressed.
Genetic Engineering for Abiotic Stress Tolerance
Lot of efforts worldwide have been made to develop abiotic stress tolerant crops through the application of recombinant DNA technology, employing a spectrum of different gene, encoding osmoprotectants, ion channel proteins, molecular chaperons and other stress-related functional proteins. The results obtained have evoked a mixed response. In comparison to other genes, biosynthetic accumulation of glycine betaine, proline and other osmoprotectant genes, in several transgenic crop plants, have shown some improvement in abiotic stress tolerance. Irrespective of the degree of successful gene transfer products developed through of this technology, it has opened up new opportunities to improve tolerance to abiotic stresses by incorporating genes involved in stress protection from any source into agriculturally important plants [11]. Quest for novel genes, required to be transferred to field crops is an integral part of this ongoing struggle against abiotic stresses (Figure 2). The transgenic approach also allows scientists to study and validate the mechanism governing stress tolerance by over-expressing the gene into a model plant species and to monitor the phenotypical and biochemical changes before and after a specific abiotic stress treatment [12, 13]. Reasonable degree of success has been achieved, and several varieties of stress tolerant crops have been released in recent years. A number of new experimental varieties are also being developed for further testing under field conditions [14]. Though resource intensive, this approach seems a viable option for developing genotypes that can perform better under sub-optimal growth conditions, given the complex nature of abiotic stress tolerance response and other environmental constraints.
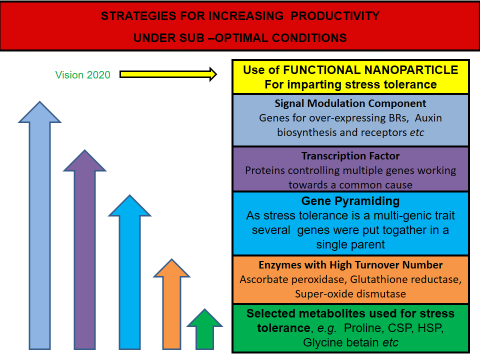
Genes Involved in Abiotic Stress Tolerance
The current success in plant abiotic stress tolerance can be significantly enhanced by characterizing individual genes and evaluating their contribution to the overall stress tolerance potential, knowledge of basic biochemical pathways, and identification of key regulatory genes of stress response pathways. The genes implicated in abiotic stresses are broadly classified in two groups. 1. Proteins that act directly. For example, osmoprotectants, detoxifying enzymes, late embryogenesis abundant (LEA) proteins and molecular chaperones. 2. Proteins involved in regulating stress responsive genes.
For example, transcription factors, protein kinases, enzymes involved in phosphor-inositol metabolism
Proteins that act directly
Engineering osmolyte biosynthesis has emerged as a viable approach in producing plant transgenics for enhanced tolerance to osmotic stresses. Osmoregulation or osmoprotection is one of the most efficient methods, developed by the stress-tolerant plants, to maintain osmoticum as well as to conserve cellular resources under unfavorable stressful regimes. However, most crop plants lack the ability to synthesize sufficient quantities of the osmo-protectants, so as to provide a meaningful degree of protection. Therefore, genes regulating the synthesis of osmo-protectants have been transformed into transgenic Cell & Cellular Life Sciences Journal
plants to confer stress tolerance, through over expression. The engineering of plants with higher concentrations of proline began with the over expression of genes encoding the biosynthetic enzymes Pyrroline-5-carboxylate synthase (P5CS) and Pyrroline-5- carboxylate reductase (P5CR), catalyzing the two steps between the substrate glutamic acid and the product proline. P5CS over expression in tobacco considerably elevated free proline levels in transgenic tobacco plants [15] and the transgenic plants were successfully tested under abiotic stress conditions. A common secondary response of various abiotic stresses is the over-production of reactive oxygen species (ROS) in plants. These ROS are inherently reactive and result in induction of oxidative stress. ROS may cause damage to proteins, lipids, carbohydrates and DNA which ultimately results in plant death. The ROS comprises both free radical (O2.-, superoxide radicals; OH., hydroxyl radical; HO2., perhydroxy radical and RO., alkoxy radicals) and non-radical (molecular) forms (H2O2, hydrogen peroxide). In chloroplasts, photosystems (PSI and PSII) are the major sites for the production of 1O2 and O2.-. In mitochondria, complex I, ubiquinone and complex III of electron transport chain are the major sites for generation of O2.-. The antioxidant defense machinery protects plants against such oxidative stress damages. Plants possess an efficient enzymatic (superoxide dismutase, SOD; catalase, CAT; ascorbate peroxidase, APX; glutathione reductase, GR; monodehydroascorbate reductase, MDHAR; dehydroascorbate reductase, DHAR; glutathione peroxidase, GPX; guaicol peroxidase, GPX and glutathione- S- transferase, GST) and non-enzymatic (ascorbic acid, ASH; glutathione, GSH; phenolic compounds, alkaloids, non-protein amino acids and a-tocopherols) antioxidant defense molecules that work in a concerted manner, to regulate the cascades of uncontrolled oxidation reactions and protect plant cells from oxidative damage. For detailed analysis of contribution of these antioxidant enzymes to stress tolerance, a large number of experiments have been conducted with transgenic model plants overproducing these antioxidant enzymes [16]. By constitutively expressing Mn-SOD into chloroplast and mitochondria, Bowler, et al. [17] observed reduced cellular damage in response to induction of oxidative stress. Among the vast majority of stress associated proteins, HSPs, CSPs and LEA type proteins are the major stress- induced proteins that accumulate upon water, salinity and extreme temperature stress. They have been shown to act as molecular chaperones, which are responsible for protecting the cellular machinery in a broad array of cellular processes. Basically, the molecular chaperones assist in stabilization of proteins and membranes and in protein refolding under stress conditions [18]. LEA-type proteins have been divided into a number of families, with diverse structures and functions [19]. The enhanced expression of transcription factors that regulate the expression of LEA type proteins was found to correlate with cold, drought, or salt stress tolerance in plants [20, 21, 22]. Xu et al. [22] reported that the expression of a group 3 protein HVA1 in barley confers tolerance to water deficiency and salt stress in transgenic rice plants. Apart from the LEA proteins, HSPs and CSPs also play important role in stress metabolism. These proteins have been divided into various sub-classes, based on their molecular mass. Genetic engineering for increased thermo tolerance by enhancing HSP synthesis in plants has been achieved in a number of plant species. For instance, expression of HSP DnaK1 from a halo-tolerant cynobacterium A. halophytica has resulted in improving salt tolerance of transgenic tobacco plants [23].
Proteins involved in regulating stress responsive genes
The gene product of this group includes transcription factors, protein kinases and enzymes involved in phosphoinositide metabolism. This group of gene products regulates gene expression and signal transduction pathways. Far beyond the manipulation of single functional gene, engineering certain regulatory genes has emerged as an effective strategy now for controlling the expression of related stress-responsive genes. Transcription factors (TFs) have emerged as effective candidates for genetic engineering to develop broad spectrum stress-tolerant crops, because of their role as master regulators of many stress-regulatory pathways. Many TFs belonging to families AP2/EREBP, MYB, WRKY, NAC, bZIP have been found to be involved in tolerance to various abiotic stresses and some TF genes have also been engineered to improve stress tolerance in model and crop plants. As one of the most important regulators, TFs function as terminal transducers and directly regulate the expression of an array of downstream genes by interacting with the specific cis- elements in their promoter region [24]. Over-expression of a constitutively active form of AtDREB2A from Arabidopsis has been reported to improve the tolerance to drought and osmotic stresses [25]. Plant growth regulators are also known to manipulate a number of physiological processes involved in stress tolerance. The phytohormone abscisic acid (ABA) plays a crucial role in the activation of cellular defense responses Cell & Cellular Life Sciences Journal
under drought as well as salinity stresses [26, 27]. Abscisic acid signals are perceived by different cellular receptors and a concept of activation of specific cellular ABA mediated responses by signal perception in distinct cellular compartments is currently accepted. The nucleo- cytoplasmic receptors PYR/PYL/RCARs (PYRABACTIN RESISTANCE/ PYRABACTIN RESISTANCE- LIKE/REGULATORY COMPONENT OF ABA RECEPTORS) bind ABA and inhibit type 2C protein phosphatases (PP2Cs) such as ABI1 and ABI2.The inactivation of PP2Cs activates accumulation of active SNF1-RELATED PROTEIN KINASES (SnRK2s) [28, 29]. In addition to two-component histidine kinases, receptor-like protein kinases have also been implicated in abiotic stress responses. A transcript of Nicotiana tabacum NtC7, a transmembrane protein, was found to accumulate rapidly and transiently within one hour of salt and osmotic stress treatment. Transgenic plants overexpressing NtC7 exhibited a marked tolerance to 12-h mannitol treatment, which suggests that NtC7 functions to maintain osmotic adjustment [30, 31, 32, 33, 34, 35, 36]. However, the regulatory mechanisms involving signaling components is very complex and further studies are needed to find out the exact role of these components, and their downstream targets in a signaling cascade. In particular, information on protein kinase substrates, which may be transcription factors, as well as cytoplasmic or membrane proteins, is very limited. Further studies, combining genetic and molecular approaches, will enrich the knowledge of the complex networks of stress signaling in plants. How the signals interact to confer cross-tolerance and developmental traits, and how this interaction influences crop productivity, are surely important questions in the research area aimed at improving agricultural productivity.
Conclusion
Development of stable transgenic crops that can withstand increased pressure of abiotic stresses and can sustain productivity under sub-optimal growth conditions is one of the foremost challenges in agricultural biotechnology. Transgenic technology has so far produced mixed results in this arena. However, there is no doubt that we have come a long way in our quest to understand and regulate plant metabolic processes under stress. This forms the bedrock of our efforts towards ensuring food and nutritional security for the masses. Several technological advances in the tools and techniques of developing transgenic crops have strengthened our efforts. An array of new concepts, starting from individual gene manipulation to signaling cascade interventions showcase the enormous efforts that have gone into perfecting this technology. Our understanding of the basic gene regulatory concepts has enabled us to use novel methods for optimizing the stress responsiveness of crop plants. Nanotechnology has the potential to augment agricultural productivity through designing and fabrication of materials and systems at the nano-scale that could leverage the exceptional properties of matter at this scale. Nanotechnological developments have opened broader avenues in the advancement of fortified crops through site directed delivery of various macromolecules including genes and drugs, smart delivery systems of agrochemicals, early detection of diseases and pathogens and precision farming techniques.
References
-
UNCTAD W (2011) Price Volatility in Food and Agricultural Markets: Policy Responses. Seeds and growth of spinach Biological trace element research 104(1): 83-91.
-
Lobell DB, Field CB (2007) Global scale climate-crop yield relationships and the impacts of recent warming. Environ Res Lett 2(1): 1-8.
-
Segele ZT, Lamb PJ (2005) Characterization and variability of Kiremt rainy season over Ethiopia. Meteorol Atmos Phys 89(1-4):153-180.
-
Seleshi Y, Camberlin P (2006) Recent changes in dry spell and extreme rainfall events in Ethiopia. Theor Appl Clim 83(1-4): 181-191.
-
Haggag WM, Abouziena HF, Abd-El-Kreem F, El Habbasha S (2015) Agriculture biotechnology for management of multiple biotic and abiotic environmental stress in crops. J Chem Pharm Res 7(10): 882-889.
-
Bray EA, Bailey-Serres J, Weretilnyk E (2000) Responses to abiotic stresses. In Gruissem W, et al. (Eds.), Biochemistry and Molecular Biology of Plants. American Society of Plant Physiologists, Rockville, MD, pp: 1158-1249.
-
Cushman JC, Bohnert HJ (2000) Genomic approaches to plant stress tolerance. Curr Opin Plant Biol 3(2): 117-124.
-
Barik TK, Sahu B, Swain V (2008) Nanosilica-from medicine to pest control. Parasitology research 103(2): 253-258. Cell & Cellular Life Sciences Journal
-
Nayan R, Rawat M, Negi B, Pande A, Arora S (2016) Zinc sulfide nanoparticle mediated alterations in growth and anti-oxidant status of Brassica juncea. Biologia 71(8): 896-902.
-
Rawat M, Nayan R, Negi B, Zaidi MGH, Arora S (2017) Physio-biochemical basis of iron-sulfide nanoparticle induced growth and seed yield enhancement in B. juncea. Plant Physiol Biochem 118: 274-284.
-
Bajaj S, Mohanty A (2005) Recent advances in rice biotechnology-towards genetically superior transgenic rice. Plant Biotech J 3(3): 275-307.
-
Bartels D, Phillips J (2010) Genetic modification of plants. Biotechnol Agric Forestry 64(2): 139-157.
-
Mantri N, Patade V, Penna S, Pang E (2012) Abiotic stress response in plants _ present and future. In: Ahmad P, Prasad MNV, editors. Abiotic stress responses in plants: metabolism to productivity. New York:Springer 1-19.
-
Atkinson NJ, Urwin PE (2012) The interaction of plant biotic and abiotic stresses: from genes to field. J Exp Bot 63(10): 3523-3543.
-
Kishor P, Hong Z, Miao GH, Hu C, Verma D (1995) Over expression of [D]-pyrroline-5-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants. Plant Physiol 108(4): 1387-1394.
-
Wang W, Vinocur B, Altman A (2003) Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta 218(1): 1-14.
-
Bowler C, Slooten L, Vandenbranden S, DeRycke R, Botterman J, Sybesma C, VanMontagu M, Inze D (1991) Manganese superoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants. EMBO J 10(7): 1723_1732.
-
Boston RS, Viitanen PV, Vierling E (1996) Molecular chaperones and protein folding in plants. Plant Mol Biol 32(1-2): 191-222.
-
Thomashow MF (1998) Role of cold-responsive genes in plant freezing tolerance. Plant Physiol 118(1): 1-8.
-
Kasuga M, Liu Q, Miura S, Yamaguchi-Shinozaki K, Shinozaki K (1999) Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress inducible transcription factor. Nat Biotech 17(3): 287-291.
-
Jaglo KR, Kleff S, Amundsen KL, Zhang X, Haake V, et al. (2001) Components of the Arabidopsis C- repeat/dehydration responsive element binding factor cold-response pathway are conserved in Brassica napus and other plant species. Plant Physiol 127(3): 910-917.
-
Xu D, Duan X, Wang B, Hong B, Ho T, et al. (1996) Expression of a late embryogenesis abundant protein gene, HVA1, from barley confers tolerance to water deficit and salt stress in transgenic rice. Plant Physiol 110(1): 249-257.
-
Sugino M, Hibino T, Tanaka Y, Nii N, Takabe T (1999) Overexpression of DnaK from a halotolerant cyanobacterium Aphanothece halophytica acquires resistance to salt stress in transgenic tobacco plants. Plant Sci 146(2): 81-88.
-
Yamaguchi-Shinozaki K, Shinozaki K (2006) Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu Rev Plant Biol 57: 781-803.
-
Sakuma Y, Maruyama K, Osakabe Y, Qin F, Seki M, et al. (2006a) Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought- responsive gene expression. Plant Cell 18(5): 1292- 1309.
-
Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR (2010) Abscisic acid: emergence of a core signaling network. Annu Rev Plant Biol 61: 651-679.
-
Raghavendra AS, Gonugunta VK, Christmann A, Grill E (2010) ABA perception and signalling. Trends Plant Sci 15(7): 395-401.
-
Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y, et al. (2009) Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 324(5930): 1064-1068.
-
Park SY, Fung P, Nishimura N, Jensen DR, Fujii H, et al. (2009) Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 324(5930): 1068-1071. Cell & Cellular Life Sciences Journal
-
Tamura T, Hara K, Yamaguchi Y, Koizumi N, Sano H (2003) Osmotic stress tolerance of transgenic tobacco expressing a gene encoding a membrane-located receptor-like protein from tobacco plants. Plant Physiol 131(2): 454-462.
-
Acquaah G (2007) Principles of plant genetics and breeding. Blackwell, Oxford, UK
-
Chen TH, Murata N (2002) Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr Opin Plant Biol 5(3): 250-257.
-
Pinheiro C, Chaves MM (2011) Photosynthesis and drought: can we make metabolic connections from available data? J Exp Bot 62(3): 869-882.
-
Takahashi S, Seki M, Ishida J, Satou M, Sakurai T, et al. (2004) Monitoring the expression profiles of genes induced by hyperosmotic, high salinity, and oxidative stress and abscisic acid treatment in Arabidopsis cell culture using a full-length cDNA microarray. Plant Mol Biol 56(1): 29-55.
-
Tattersall EA, Grimplet J, Deluc L, Wheatley MD, Vincent D, et al. (2007) Transcript abundance profiles reveal larger and more complex responses of grapevine to chilling compared to osmotic and salinity stress. Funct Integr Genomics 7(4): 317-333.
-
Yancey PH (2001) Water stress, osmolytes and proteins. Amer Zool 41: 699-709.
- The Muculent Bleb-Mucinous Cystic Neoplasm-Hepatobiliary Region
- Insulin Sensitizers as Anti-Aging Agents: Unveiling Synergies with Albumin, GLP-1RA, Klotho Protein, and Metformin in the Quest to Combat Aging
- Reprogramming of GLP-1 Response at Prediabetes for the Prevention of Type 2 Diabetes: The Role of Albumin and GLP-1 Receptor Agonists
- The Mingled Allies-Combined Hepatocellular Carcinoma and Cholangiocarcinoma
- Compilation and Embodiment-Leydig Cell Tumour Testis
- Glucolipotoxicity: A Novel Different Perspective on the Causes of Cancer