Avoidance of Whitening of Brownadipose Tissue (BAT/Beige Adipose Tissue (BeAT)-as an Innovative Target of Obesity/Ageing Prevention-A Mini Review
Whereas we have been working regarding the browning of WAT for practically last 2 decades in obesity research, whitening of BAT as an obesity-correlated complication possessing metabolic along with health aspects has not got considerable importance. Here our aim has to be focusing on why there is an escalation of BAT whitening which by itself becomes a considerable cause of obesity and its complications. Here we have paid attention to Natural factors, High Fat Diet, Ageing, part of nicotine, thermo neutrality, Serotonin, Glucocorticoids obesogens, Dechlorane plus endocrine disrupting chemicals (EDCs), like wide spectrum of consumer products, like tobacco, flame retardants, as well as pesticides which we are exposed in common day to day life apart from consumption of nicotine in the prenatal or lactation exposure to the offspring as well as the origin of chronic diseases beyond infancy. Furthermore our concentration has been over how autophagy, inflammation, along with dysfunctional angiogenesis “vascularization” are central events involved in BAT/beige AT whitening of these we have detailed role of VEGF-A besides role of Bevacizumab its antibody , capillary rarefaction as well as hypoxia in a little detail. Ageing is a natural event. When BAT starts receding specifically in females direct association of BAT with reproduction is seen & apparently FSH antibody is acknowledged to revert post menopause and post-andropause diminished metabolic rate that is associated with escalated visceral adiposity in addition to escalate longevity. Moreover, recently the group of Vatner, et al. have discovered that interference with the Regulator of G protein signaling14 (RGS14) in mice escalates healthy lifespan, modulation by escalated BAT. Thus RGS14 seems to be a target that’s hampering apparently escalated longevity in addition to BAT. Capillary rarefaction impacts numerous organs .Hence avoidance of smoking in pregnancy might be a significant long term factor in preventing chronic diseases beyond infancy specifically in males in view of females get protected by estrogen. Thus greater focus is needed to avoidance of whitening of BAT to combat the epidemic of obesity. Too much misuse of dexamethasone prescription needs to be avoided since post covid era.
Introduction
With the escalating obesity along with type2 diabetes mellitus (T2DM) worldwide it has now been decided to treat the condition as disability. Hence the choice of the oral hypoglycaemics has to be the one which aid in weight reduction. We have reviewed the different oral hypoglycaemics along with insulin, Sodium–glucose transporters (SGLTs) along with observed SGLTs were the most promising being cardio & renoprotective besides having weight reduction actions contrary to weight gain action of glitazones in particular pioglitazones. Earlier we had further reviewed the part of browning of White Adipose tissue(WAT) to Beige adipocytes along with classes of drugs specifically role of β-3 adrenergic receptors agonists like mirabegron. Further we summarized the mechanism of weight reduction caused by SGLT hampering agents besides their glycosuria facilitating action along with role of macrophage Polarization on Insulin Resistance (IR). In case of SGLT2 hampering agents along with the resurgence of emphasis over browning of WAT to form Beige adipocytes and update over role of exercise, irisin cold exposure for weight. Nevertheless, whereas we have been working regarding the browning of WAT for practically last 2 decades in obesity research, whitening of BAT as an obesity- correlated complication possessing metabolic along with health aspects has not got considerable importance. Thus here we lay emphasis on this latter aspect why whitening of BAT might be the initiator of obesity and complications.
Adipose tissue(AT) inclusive of White Adipose tissue(WAT),brown adipose tissue (BAT), beige adipose tissues (Be AT) carry out necessary functions regarding sustenance of whole –body energy homeostasis [1]. BAT reflects a fat tissue that has become specialized regarding adaptive non shivering thermogenesis for heat production on exposure to cold stress. BAT takes part in primary metabolism as well as energy expenditure (EE) along with can be promptly stimulated with utilization of thermal in addition to dietary stimuli [2]. As per a recent study it was illustrated that an escalation in body mass along with function might work in the form of an efficacious therapeutic target in the context of obesity in addition to other associated metabolic diseases in such cases [3]. BAT depots got accidentally discovered in inter scapular areas of smaller mammals and human infants by the utilization of 2‒deoxo‒2‒Fluoro‒D– Glucose (18F‒FDG)/ positron emission tomography (PET)/ computed tomography (CT) scanning methods [4, 5, 6].
Detection of BAT was further feasible by these scanning methods in particular in supraclavicular, cervical and paravertebral areas of adults in neck area (biopsy proven) [4, 5]. These scanning methods of PET-CT illustrated a robust positive association amongst BAT action along with basal metabolic rate (BMR). Additionally, BAT activation possessed an inverse association with age, body mass index (BMI) [7], as well as adiposity in adults. The young along with lean females have greater metabolically active BAT [8].
![Figure 1: Coutesy A schematic representation of BAT occurrence in the interscapular region in an infant and in the supraclavicular, cervical, and paravertebral regions in an adult human [7].](/fulltextimages/10324/fig_1.png)
Brown adipocytes possess individual characteristics, like multiloculated small lipid droplets, with up regulated uncoupling protein1 (UCP1), enrichment of mitochondria, capillaries [3]. BAT thermogenesis is basically based on UCP1, a (biomarker for BAT) on the mitochondrial inner membrane for energy dispersal [9, 10]. UCP1 results in uncoupling of adenosine triphosphate (ATP) generation from oxidation of energy substrates, thereby facilitating non generational EE via escalated mitochondrial uncoupling[5, 8, 9].The action as well as formation of brown adipocytes are controlled by the sympathetic nervous system(SNS) thermogenesis is regulated by them leptin melanocortin pathway [11, 12]. See (Figure 1) [13].
Coutesy Schematic overview showing the characteristics of brown, white, and beige/brite adipocytes, including their localization, morphology, physiological function, and marker genes [14]. In brief, the anatomical location of these adipocytes differs, except for white and beige adipocytes which are found within WAT depots. In terms of morphology and function, brown adipocytes are comprised of small multilocular lipid droplets, high mitochondrial content, and increased UCP1 expression, which can be also induced in beige adipocytes to promote thermogenesis and enhance energy expenditure, features that are important for the treatment of obesity. In sharp contrast, white adipocytes have fewer mitochondria and large unilocular lipid droplets to facilitate the storage of energy in a form of fats. Of note, these adipocytes have similar and distinct marker genes which could play an important role in tracking metabolic complications linked with obesity. Cidea, cell death-inducing DNA fragmentation factor-like effector A; CD137, tumour necrosis factor receptor superfamily, member 9; Dio2, Iodothyronine deiodinase 2; Lhx8, LIM home box protein 8; Pgc1A, peroxisome proliferator-activated receptor coactivator 1 alpha; Prdm16, PR domain-containing 16; Tcf21, Transcription factor 21; TMEM26, trans membrane protein 26; UCP1, uncoupling protein 1; WAT, white adipose tissue; Rb1, RB transcriptional corepressor 1; Zic1, zinc finger protein of the cerebellum.
Earlier we had reviewed different classifications of Brown or “Beige” Adipocyte Physiology and treatment strategies like β3 adrenergic agonists, and other 4 classes targeted inc of Mirabegron, roles of PRDM, BMP7, MED axis in addition to role of SGLT inhibitors [14, 15, 16, 17, 18, 19]. Here we considered how this knowledge might be applied of browning of WAT in treating PCOS.
Methods
Thus a narrative mini review was carried out using the PubMed, Web of Science , Medline, Embase, Cochrane reviews, and Google Scholar, Search engine with the MeSH Terms; impaired lipid metabolism; oxidative stress; inflammation; T2DM); Type 1 diabetes (T1D);SGLT2 inhibitors; empagliflozin; cangliflozin; NAFLD ;Obesity ; weight reduction; whitening of Brown Adipose tissue (BAT); White Adipose tissue(WAT); Macrophage Polarization; Brown Adipose tissue (BAT); CVS protection; renoprotectionin T2D from 2001 till date up to 2023to update part of agents whitening BAT on treatment of VARIOUS diseases.
Results
We found a total of 750 articles, out of which we selected 104 articles for this review. No meta-analysis, were done.
Natural Factors Stimulating BAT/Beigeadipose Tissue Whitening
Whereas we have been working regarding the browning of WAT for practically last 2 decades in obesity research, whitening of BAT as an obesity-correlated complication possessing metabolic along with health aspects has not got considerable importance. Regarding the latter, there is escalating proof in the context of BAT whitening, along with this incident numerous factors are implicated in its origin. Noticeably, it has long been deciphered that risk factors like diet, age, genetics demonstrated certain chemicals possessed the capacity of negatively influencing the health of the general population [19]. Escalated ingestion of unhealthy diet can directly aid in adiposity or weight accrual [20], while aging is an undeniable a major consequence that has long been associated with inimical health [21]. Despite, the information that it significantly affects metabolic health, exact pathological modes implicated in this event require clarity. Intriguingly, experimental validation pointed that factors like diet, thermo neutrality, age, as well as genetics can stimulate whitening of BAT, along with exaggerate obesity.
High Fat Diet Stimulated BAT/Beige Adipose Tissue Whitening by Repressing Angiogenesis along with Escalating Inflammation
We possess the information that intake of a diet having enrichment of fat, cholesterol, as well as sugar is the maximum frequent etiology of obesity in addition to metabolic syndrome (MetS) [22, 23, 24]. Actually, animal models are good models for evaluation of this type of a diet, with validation pointing that feeding rats two distinct constituents of a western-style diet 60%HFD along with 55% high fructose diet can result in fat accrual, drive a state of insulin resistance(IR) along with other metabolic complications [25, 26]. Dependent on enhancing growing experimental validation, exposure of rodents to HFD-feeding can result in variable pathological aberrations that are inclusive of adipocyte hyperplasia along with hypertrophy [25]. The latter is correlated with the state of inflammation, whereby AT expansion causes the liberation of proangiogenic cytokines, like leptin, adiponectin, VEGF, TNF-a, as well as transforming growth factorbeta (TGF-β) angiopoietin; already reviewed extensively [27, 28]. Amongst other cytokines, VEGF-A is a main proangiogenic factor that is frequently downregulated in obesity (Figure 2) [29, 30, 31].
![Figure 2: Courtesy ref no -27-Regulation of vascular endothelial growth factor A (VEGFA)/ vascular endothelial growth factor receptor 2 (VEGFR2) signaling. (a) Placental growth factor (PlGF), VEGFB and VEGFA compete for binding to VEGFR1. VEGFA has ten times higher binding affinity to VEGFR1 than to VEGFR2 ((b) Binding of VEGFB to VEGFR1 displaces VEGFA and increases its bioavailability for binding to VEGFR2, which then induces angiogenic signaling via VEGFR2 activation.(c) Binding of PlGF to VEGFR1 increases angiogenesis both by displacing VEGFA and making it available for VEGFR2-binding and by activating PlGF’s own, albeit weaker, angiogenic signaling viaVEGFR1 (d) Neuropilin-1 (NRP1) binds to VEGFA and presents it to VEGFR2, which enhances VEGFA/VEGFR2 signaling (reviewed in Reference [78]). (e) Binding of Angiopoietin 1 (ANGPT1) to its receptor TIE2 on endothelial cell (EC) surface activates signaling complex formation and recruits vascular endothelial protein tyrosine phosphatase (VE-PTP) to EC-EC junctions, which reduces VEGFR2activity.](/fulltextimages/10324/fig_2.png)
Figure 2: Courtesy ref no -27-Regulation of vascular endothelial growth factor A (VEGFA)/ vascular endothelial growth factor receptor 2 (VEGFR2) signaling. (a) Placental growth factor (PlGF), VEGFB and VEGFA compete for binding to VEGFR1. VEGFA has ten times higher binding affinity to VEGFR1 than to VEGFR2 ((b) Binding of VEGFB to VEGFR1 displaces VEGFA and increases its bioavailability for binding to VEGFR2, which then induces angiogenic signaling via VEGFR2 activation.(c) Binding of PlGF to VEGFR1 increases angiogenesis both by displacing VEGFA and making it available for VEGFR2-binding and by activating PlGF’s own, albeit weaker, angiogenic signaling viaVEGFR1 (d) Neuropilin-1 (NRP1) binds to VEGFA and presents it to VEGFR2, which enhances VEGFA/VEGFR2 signaling (reviewed in Reference [78]). (e) Binding of Angiopoietin 1 (ANGPT1) to its receptor TIE2 on endothelial cell (EC) surface activates signaling complex formation and recruits vascular endothelial protein tyrosine phosphatase (VE-PTP) to EC-EC junctions, which reduces VEGFR2activity.
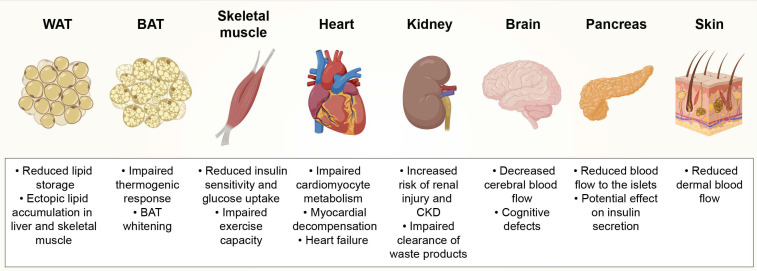
Actually, AT vascularization controls chronic inflammation in addition to systemic insulin sensitivity [32]. In obese subjects, AT displayed capillary rarefaction as well as hypoxia, which are in agreement with the infiltration of macrophages along with the liberation of inflammatory cytokines [29, 33]. Obesity along with its correlated metabolic diseases is associated with dysfunction to vascular impairment as well as along with elimination of capillaries known as “capillary rarefaction” (Figures 3 & 4).
![Figure 4: Endothelial regulation of adipose tissue (AT) phenotype. Factors such as diet-induced obesity reduced vascular density due to a loss of Vegfa or an increase in the anti-angiogenic is of orm VEGFA165b may cause whitening of AT. On the other hand, increased vascular density due to Vegfa or Vegfb overexpression or Vegfr1 ablation] can induce browning of WAT in mice. Interestingly, the suppression of Vegfa expression in mice also led to a browning phenotype in WAT, suggesting that further studies are required to conclude the exact role of VEGFA in this phenomenon. Recently, endothelial protein argonaute-1 (AGO1) suppression was also shown to increase vascular density and AT browning.](/fulltextimages/10324/fig_4.png)
Figure 4: Endothelial regulation of adipose tissue (AT) phenotype. Factors such as diet-induced obesity reduced vascular density due to a loss of Vegfa or an increase in the anti-angiogenic is of orm VEGFA165b may cause whitening of AT. On the other hand, increased vascular density due to Vegfa or Vegfb overexpression or Vegfr1 ablation] can induce browning of WAT in mice. Interestingly, the suppression of Vegfa expression in mice also led to a browning phenotype in WAT, suggesting that further studies are required to conclude the exact role of VEGFA in this phenomenon. Recently, endothelial protein argonaute-1 (AGO1) suppression was also shown to increase vascular density and AT browning.
Various lines of proof revealed that HFD as well as high-fat high-sugar diet stimulate BAT/beige adipocytes impairment along with whitening via vascular rarefaction correlated with the state of inflammation [34, 35, 36, 37, 38, 39]. Despite, the whitening has been lesser studied in beige AT in contrast to BAT, the (whitening of beige adipocytes was found in HFD-fed C57BL/6 mice, as well as it possessed remarkably escalated adipocyte size along with diminished expression of UCP1 [35]. In canonical BAT, HFD-feeding progressively stimulated fat deposition in BAT, which caused the expansion in addition to whitening of BAT in female CD-1 mice [40]. Akin to that chronic HFD feeding in mice led to progressive BAT whitening, which was correlated with lesser energy expenditure (EE), in addition to down-regulation of gene expression of Bmp8b, Nrg4, Vegfa implicated in controlling vascularization, along with up regulation of inflammasome along with endoplasmic reticulum stress [41]. Furthermore, BAT from HFD-fed C57BL/6J mice revealed lipid accrual along with IR, which were associated with diminished glucose as well as triglyceride- obtained fatty acids uptake [36]. Noticeably, Ucp1 gene expression was not changed, while the expression of PGC1α along with protein kinase B (PKB) was repressed, pointing that dysfunctional mitochondrial biogeneration as well as insulin sensitivity [36]. Greater understanding regarding the mode illustrated that HFD-induced iBAT whitening in C57BL/6 mice, which were associated with diminished expression of the genes facilitating vascularization, thermogenesis, fatty acids oxidation (FAO) inclusive of VEGF-A, Ucp1, β3-AR, C idea, along with carnitinepalmitoyltransferase (CPT) [37]. In the same study, they illustrated that counteracting BAT whitening using PPARα agonists can aid to abrogate the complications associated with obesity [37]. The combination of HFD and high-sugar diet resulted in dysfunctional BAT function in Wistar rats [38]. The proof of whitening in these animals was considerable by the escalated adipocytes region along with diminished expression of BAT markers, like FGF21, PPARγ, SIRT1, along with CPT1, in addition to the genes implicated in the insulin signaling inclusive of insulin receptor substrate 2 (IRS-2), as well as glucose transporter (GLUT)-4 [38]. On observing an association amongst the pathogenesis of obesity along with its risk factors, maternal nutrition has assumed to be a target regarding gaining insight into the generation of obesity and further [42]. These nutrition aids significantly (to the generational origins of metabolic complications on growth till adulthood [43]. Despite, playing a key part in new-borns as controller of energy homeostasis as well as thermogenesis [44, 45], clarification is required regarding how maternal nutrition impacts offspring AT function. Recently, Savva, et al. [39], evaluated the impact of maternal HFD on AT programming in male as well as female C57Bl6/J mice offspring.
Intriguingly, only male offspring illustrated a whitening of BAT along with resulted in dysfunctional metabolic profile while female counterparts presented with enhanced thermogenesis as well as cell differentiation in BAT [39], which can be in view of the presence of estrogen in females [46]. Conversely, the whitening of BAT in male offspring was associated with the upregulation of the genes implicated in lipid metabolisms like Cd36, Cpt1, Cidea, along with DIO2, UCP1, PGC1a as well as PRDM16 [39].
Thermoneutrality- Stimulated BAT Whitening via Stimulating Mitophagy along with SNS Obtained Diminished Signals
Surrounding temperature possesses a robust influence on body metabolism as well as EE, which in turn impacts the morphology along with thermogenesis function of BAT. Despite, the thermo neutral situation of about 22°C is the standard living milieu’s situation for humans, it is acknowledged to result in BAT involution as well as adiposity in rodents [47], and still no clarification exists regarding the reversed action of seasonal alterations on BAT thermogenesis along with plasticity in humans. Apparently at the time of winter season, human subcutaneous(s/c) WAT revealed escalated mRNA expression of UCP1, PGC1a, in addition to trans membrane protein 26 (TMEM26), along with other genes implicated in energy utilization as well as lipolysis, like adiponectin, acetyl CoA carboxylase (ACC), and HSL [48]. Nevertheless, this action was repressed by escalated lipid accrual along with inflammation in obesity [48]. In healthy men, whole-body EE as well as cold-stimulated thermogenesis was evaluated in both summer along with winter, using fluorodeoxy glucose (FDG)-positron emission tomography (PET) combined with computed tomography (CT) [49]. Cold- stimulated thermogenesis was escalated in winter in contrast to summer in a BAT-based way, pointing that the metabolic activity of human BAT is maximal in winter [49]. Regarding glucose metabolism, it was revealed that winter swimmers illustrated no BAT glucose uptake at a thermal comfort zone whereas winter swimmers have higher cold- stimulated thermogenesis in contrast to control subjects in young healthy men [50]. In mice, a thermo neutral zone of 30°C is used for thermally humanizing mice BAT, which illustrates a considerable similarity with human BAT [51]. It has been displayed that thermo neutral housing of mice in association with or without a high- calorie diet, robustly diminishes metabolic capacity along with escalates lipid accrual in BAT, resulting in a “white- like” appearance [52, 53, 54]. (Actually, A/J mice housed at 30°C with HFD revealed paler and larger brown or beige adipocytes [53]. This was associated with diminished SNS and thermo genic program which were apparent by reduced tyrosine hydroxylase and norepinephrine turnover, as well as the decreased mRNA expression of UCP1, PGC1a, DIO2, elongation of very long chain fatty acids protein 3 (Elovl3) and cyclooxygenase (Cox)-1 [53]. Other proof illustrated that BAT whitening can be correlated with the enrollment of immune cells implicated in pro-inflammation as well as mitophagy. An environmental temperature of 28 illustrated that interscapular and mediastinal BAT from C57Bl/6j mice acquired a white-like unilocular adipocyte phenotype, which involved escalated macrophage infiltration, formation of crown-like structures, along with degenerating mitochondria, marked by adipose triglyceride lipase (Atgl)-deficiency [52].
A study by Sass, et al. [54], illustrated that thermo neutral adaptation at 30 induced BAT whitening which possessed properties of escalated unilocular adipocytes as well as mitochondrial breakdown in Wild-type mice. This was also associated with the diminished gene expression of thermogenic markers, inclusive of UCP1, PGC1a,cytochrome coxidase subunit 4 isoform 1 (Cox4i1), cytochrome c oxidase subunit 7A (Cox7a), along with cytochrome c oxidase subunit 8B (Cox8b), whereas the expression quantities of the autophagy-controlling transcription factor EB (TFEB) was progressively escalated [54]. Moreover, the hampering of autophagy reversed the whitening in BAT [54]. This agrees with the prior proof pointing that metabolism in part gets controlled by mitochondrial turnover in mice [55].
Aging-Induced BAT/beige Adipose Tissue Whitening by EScalating lncRNAs Expression along with Interferon-γ Liberation
Aging has long been believed to be responsible for AT impairment as well as escalated risk of obesity [56]. A remarkable decrease in BAT as well as beige AT with advancing years along with escalating body fat percentage has been estimated [57]. In general, aging is intricately correlated with low-grade systemic inflammation, as well as alterations in endocrine signals [58]; modes that are associated with BAT impairment upon aging [57]. Numerous modes that might aid in the age- associated reduced BAT activity have been studied in animals along with humans. Like Berry, et al. [59], have revealed that mouse along with human beige progenitor cells undergo an age- associated senescence-like phenotype which explains age-based beiging failure; nevertheless, genetic or pharmacological activation of p38/Ink4a-Arf pathway rejuvenated beige progenitors and restored beiging potential. Tajima, et al. [60] isolated mitochondria lipoylation as a molecular event underlying the age- associated decrease in BAT thermogenesis of mice, pointing that an aberration in mitochondrial lipoylation along with fuel oxidation in BAT, resulted in glucose intolerance as well as obesity upon aging. On the other hand α-lipoic acid supplementation escalated mitochondrial lipoylation which in turn restored BAT function in aged mice [60]. A noticeable finding was made by the reduced BAT quantities as well as action associated with the whitening at the time of adiposity in rodents as well as rabbits [61]. In female C57BL/6 mice aged (6-12 months old), BAT illustrated a morphological alteration toward the fat storage phenotype with escalated lipid droplet size as well as area [62]. This was associated with the elimination of clusters of beige adipocytes from subcutaneous as well as visceral WAT [62]. To unearth the underlying mode, a recent study by Pan et al. [63], demonstrated an escalated unilocular lipid droplets in addition to senescent T cells infiltration which induces BAT whitening via interferon (IFN)-γ in 18-month-old and
3-month-old mice. Furthermore, IFN- γ liberation resulted in the hampering of brown pre-adipocyte differentiation that aided in AT remodeling in aged mice [63]. This was further verified using stromal vascular fraction cells isolated from BAT and T cells co-culture, which revealed diminished UCP1, PPARγ, PGC1α, Plin1, in addition to adiponectin gene expression [63]. Despite, the subcutaneous as well as visceral WAT probably constituted of beige adipocytes were escalated in size, UCP1 was poorly determined in young mice as well as it was not determined in old mice, pointing that there was a total elimination of thermo genic capacity upon aging. A shift from brown/beige- to white-like adipocytes was observed in FVB mice subsequent to 12- months of age [64]. Moreover, the transcriptional profile of adipose-derived stromal cells portray these alterations of both at mRNA in addition to microRNA transcriptional quantities via E2F transcription factor 1 (E2F1) as well as nuclear factor kappa B (NFκB) [64]. In 2 years old (aged) rabbits, BAT presented large unilocular lipid droplets with a drastically diminished transcriptional copy numbers of the mitochondrial genes, inclusive of cytochrome B,COX2, as well as NADH dehydrogenase 1 [65]. Intriguingly long non-coding RNAs (lncRNAs) were highly expressed in the BAT of aged rabbits. On assessment utilizing in vitro model, these lncRNAs apparently resulted in dysfunctional brown adipocyte differentiation. Presumably, lncRNAs repressed the expression of brown adipocyte transcriptional factors, and nevertheless, this needs further assessment [65]. Recently, Huang, et al. [66] have illustrated that rabbit BAT involutes in a way akin to humans with the analogous progenitor hierarchy. Continuous whitening with adipocyte hypertrophy along with elimination of UCP1 expression in the interscapular, subscapular, as well as suprascapular BAT depots of the rabbits was observed from 3 weeks of age and full transformation to WAT-like tissues at 12 weeks [66]. This BAT whitening was correlated with the limited adipogenic capability of follistatin-like 1 (Fstl1+) progenitors [66]. Furthermore elimination of the Fstl1 gene or ablation of Fstl1+ progenitors in mice induced BAT scarcity [66]. This pointed that lnc RNAs can be one of the molecular drivers of BAT whitening upon aging. Nevertheless, this needs validation in human studies.
When maximally stimulated, BAT tunes on the anabolism in addition to catabolism of macromolecules. This takes place in view of BAT distinctly dissociates oxidative phosphorylation as well as ATP development for heat production. In view of BAT’s distinct metabolic phenotype to work in the form of metabolic sink, researchers have shifted to BAT to overcome the escalating obesity along with diabetes epidemics plaguing the developed world. The overlap of obesity in addition to aging is well , acknowledged as well as thus there has been further incentive in BAT’s part in postponing the underlying events of aging (summarized in Figure 4) [67]. Specifically, activating BAT apparently is a feasible approach for reverting the post menopause and post- andropause diminished metabolic rate that is associated with escalated visceral adiposity. Whereas a direct correlation amongst the escalated BAT activity in addition to longevity can be acknowledged (that been illustrated in various studies), it is significant to not lose sight of the endocrine function of BAT as well (Figure 5).
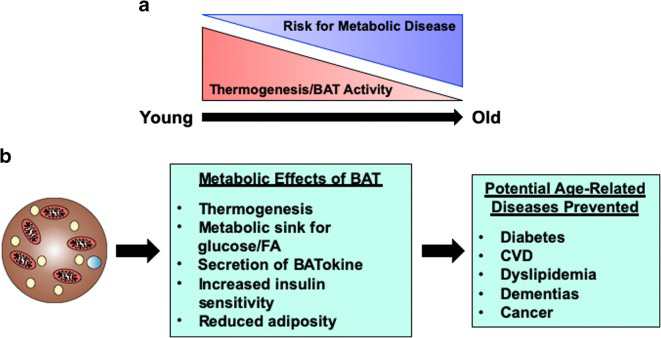
Figure 5: Courtesy ref no -67-Brown adipose tissue and aging. (A) Brown adipose tissue (BAT) mass and activity decreases throughout life. This decrease is associated with an increased risk of developing metabolic syndrome, especially in the post menopause and post-andropause life stages. (B) BAT has major metabolic functions including thermogenesis, secreting batokines, and acting as a metabolic sink for glucose and lipids. BAT reduces obesity and increases insulin sensitivity, which help prevent age-related diseases such as diabetes, dementias, and CVD. Thus, BAT may potentially be a therapeutic option to treat aging, as well as treating obesity.
Escalated exercise ability properties apart from being a characteristic of healthful aging, further works in the form of a therapy for aging patients in addition to patients with cardiovascular disease (CVD). Interference with the Regulator of G protein signaling14 (RGS14) in mice escalates healthy lifespan, modulation by escalated BAT. Very recently Vatner, et al. [68], as per that estimated if RGS14 knockout (KO) mice illustrated escalated exercise ability in addition to part of BAT with regards to modulating exercise. As per that, they estimated if RGS14 KO mice illustrated escalated exercise ability along with the part of BATin modulating exercise ability. Exercise was conducted on a treadmill as well as exercise ability by maximal running distance in addition work till exhausted. Estimation of exercise ability in RGS14 KO mice as well as their wilder type (WT) counterparts along with WT mice with BAT as well as 154±6% transplantation from RGS14 KO mice or from other mice. RGS14 KO mice illustrated 160 ± 9% escalated maximal running distance along with their WT escalated work till exhausted in contrast to WT mice. RGS14 KO mice BAT transplantation to WT mice led to phenotype getting reversed with the WT mice in receipt of BAT transplant from RGS14 KO mice illustrating 151±5% escalated maximal running distance as well as 158±7% escalated work till exhausted, at3 days subsequent to BAT transplantation in contrast to RGS14 KO donor mice. Transplanting BAT from WT to WT mice further caused escalated exercise being performed, however, not at3 days, however just at 8wks following transplantation. BAT escalated exercise ability got modulated by i) mitochondrial biogeneration as well as SIRT3;II)antioxidant defense as well as MEK/ERK pathway in addition to hind limb perfusion. Hence BAT modulated escalated exercise ability ,a mode that is more robust with RGS interference [68]. Earlier Vatner, et al. [69], had illustrated how interfering with the Regulator of G protein signaling14 (RGS14) escalated longevity and metabolism by BAT, hence RGS14 interference becomes a good target for ageing &obesity.
Genetic Models of Type 2 Diabetes Presented BAT Whitening
The belief of the genetic change as the etiology of obesity has been progressively assessed in the past two decades [70]. Dependent on the years of discoveries, the genetic etiologies of obesity can be widely classified into polygenic as well as monogenic [70]. In particular, polygenic obesity (or common obesity) is caused by multiple gene mutations or polymorphisms that facilitate weight accrual [71]. Conversely, monogenic obesity which is inherited in a Mendelian pattern is canonically scarce having the properties of early-onset, robust, along with a single gene mutation in the leptin melanocortin pathway [70, 71]. In view of these, maximum people with obesity have some mutated genes that make them prone to gain escalated weight. For a better acquisition of insights of genetic obesity, animal models of obesity as well as type 2 diabetes like leptin receptor deficient db/ db mice, as well as Zucker fatty fa/fa rats have been widely utilized [61]. Over the past few years, these models have been further utilized to study the impact of genetic obesity on the morphology along with function of BAT. Nevertheless, the particular concentration herein is on BAT whitening. A study by Lapa, et al. [72], revealed that BAT from Zucker diabetic fatty fa/fa rats, showed a ubiquitous white adipose- like tissue phenotype, with large unilocular lipid droplets as well as dysfunctional glucose utilization at 14 weeks of age. This BAT involution was correlated with the escalated synthesis as well as accrual of intracellular fatty acids, as well as the decreased expression of UCP1 [72]. Although the underlying molecular modes have not been established thus far, iBAT from db/db mice reveales whitening as well as crown-like structure generation at 13weeks of age [52]. In a non-obese model of T2D, which shares a predisposed locus on chromosome 10 like in humans, lower cell density as well as higher adipocyte area was recorded in Goto-Kakizaki rats [38]. Intriguingly, glucose uptake in BAT was dysfunctional in both baseline as well as subsequent to 30 min of stimulation with 1 mg/kg CL316,243, a β3-adrenergic agonist as well [38]. Subsequently, there was an escalated expression of genes implicated in FAO (CPT1 andCPT2), BAT metabolism (Sirt1 and PGC1α), however diminished gene expression of GLUT- 1 occurred in contrast to other experimental groups [38]. Possibly, dysfunction of the β3adrenergic response could indicate an escalated expression of the above-mentioned genes that work in the form of a compensatory mode.
Chemically Stimulated BAT/Beige Adipose Tissue Whitening
In parallel with other risk factors, chemicals like endocrine disrupting chemicals (EDCs) are acknowledged significantly in aiding in the greater prevalence of obesity [70]. These chemicals are observed in a wide spectrum of consumer products, like tobacco, flame retardants, as well as pesticides that people possess great probability of being exposed to in their daily life, via consumption, inhalation, or direct dermal contact [73], The probable targets for EDCs are the glucocorticoid along with mineralocorticoid receptors, that are members of the steroid receptor subfamily that mediate the effects of glucocorticoids as well as mineralocorticoids, the main classes of corticosteroids [74]. These chemicals can act directly on AT to stimulate hypertrophy along with impairment of this tissue [75]. Various proof have validated that exposure to some chemicals can negatively influence the phenotype along with physiological functions of BAT as well as beige AT by stimulating whitening .
Bevacizumab Results in Dysfunctional Vascular Network along with Stimulates Whitening in BAT
Bevacizumab, a recombinant humanized anti-vascular endothelial growth factor (VEGF) antibody, is trailed in retinopathy of premature infants [76], Although anti-VEGF agents are the first-line treatment for various angiogenesis- associated retinal diseases, it is not clear how the anti-VEGF antibody can aggravate the risks of systemic complications subsequent to intravitreal injection in premature infants [77], A study by Jo, et al. [78], demonstrated that intravitreally injection with anti-VEGF antibody (1 mg/eye) escalated lipid droplet accrual as well as stimulates the elimination of vascular network in neonatal C57BL/6 mice. Moreover diminished VEGF quantities were associated with the down regulation of mitochondria- associated genes PGC1α as well as UCP1 [78]. As BAT is a highly vascularized tissue, it is clear that anti-VEGF agents disturb BAT vascularization, which in turn stimulates BAT whitening along with impairment.
Nicotine Exposure During the Prenatal and Lactation Period Stimulates BAT/Beige Adipose Tissue Whitening in Offspring
Nicotine is a chemical that is widely observed in tobacco, and it has been correlated with, many health problems [79]. Accordingly, epidemiological studies have displayed that maternal smoking at the time of pregnancy might be a serious risk factor for childhood obesity [80]. Of major bother, maternal nicotine exposure has become a growing risk factor for the health of the offspring as well as the origin of chronic diseases beyond infancy. For example, prenatal or lactation exposure to nicotine is associated with increased gonadal and inguinal subcutaneous WAT depots as well as impairment in offspring [81]. Nevertheless, clarification is required regarding the influence of nicotine on BAT structure along with function. Earlier, it was revealed that nicotine exposure in adult mice escalated BAT thermogenesis as well as facilitated weight elimination through the inactivation of hypothalamic AMPK [82], along with stimulated WAT browning via the hypothalamic κ opioid receptor [83]. Paradoxically, maternal nicotine exposure stimulated BAT impairment as well as weight accrual in both male offspring [84, 85].
In male rat offspring exposed to 1 mg/kg nicotine at the time of pregnancy as well lactation; BAT revealed a whitening phenotype with the properties of lipid droplet accrual in addition to led to dysfunctional mitochondrial structure [84]. Furthermore, the expression of BAT structure as well as function- associated genes like PRDM16, PGC1α, UCP1, along with CPT2 was decreased. Similarly [85], female rat offspring exposed to 1 mg/kg nicotine during pregnancy along with lactation presented white-like adipocytes, dysfunctional angiogenesis, along with aberrant mitochondrial structure in iBAT. This was associated with a down-regulation of brown-like genes PGC1α, UCP1, Prdm16, along with C idea, as well as a decrease in BAT liberation of pro-angiogenic factors including VEGF, VEGF receptor 2,hepatocyte growth factor (Hgf), neuropeptide Y (Npy), as well as resistin [85]. This was further validated in vitro using C3H10T1/2 cells, revealing a diminished beige “brown-like” phenotype along with angiogenesis, as well as brown-like gene expression of PGC1α along with UCP1. Into to, this validation pointed that nicotine interferes with angiogenesis in the early generation stage as well as causes dysfunctional blood vessel generation to stimulate BAT whitening via down regulation of the PGC1α–UCP1 signals.
Dechlorane plus Disrupted Mitochondrialucp1 along with Stimulates BAT Whitening Dechlorane plus, an endocrine-disrupting chemical found in inflame retardants, is a potential obesogen [86]. Apparently, dechlorane plus can induce adiposity by promoting adipogenesis in cultured adipocytes via PPARγ independent mode [84]. In agreement, Peshdary, et al. [87], illustrated that dechlorane plus induces a WAT-like phenotype as well as interferes with thefunction of BAT in part by down regulation of UCP1 mRNA expression in C57BL/6 mice. These outcomes are in line with UCP1 gene knockout in mice pointing that elimination of UCP1 could lead to the whitening of BAT [88, 89]. Nevertheless, greater research on the influence of dechlorane plus on BAT as well as beige AT function is required to better gain insight over its impact on obesity as well as other associated diseases.
Serotonin-Results in Dysfunctional Brown Adipocytes Differentiation along with stimulates Beige Adipocyte Whitening
Endogenous chemicals like neurotransmitters like serotonin are acknowledged to control adipogenesis [90]. A study by Rozenblit Susan, et al. [91], illustrated serotonin (10 mM) stimulates whitening in palmitic acid- exposed HIB1B adipocytes by shifting their metabolism to lipogenesis instead of lipid oxidation in part by, repressing brown adipocytes differentiation as well as stimulating beige adipocytes trans differentiation into white adipocytes. Furthermore, this was validated by the down regulation of brown adipocyte differentiation markers, like Prdm16, Bmp7, along with Pparg [91], Consistently, expression of BAT markers like UCP1, FGF21, as well as pAMPK/AMPK ratio were down regulated while genes regulating lipogenesis fatty acid synthase (FASN), leptin, as well as adiponectin were upregulated [91], The observed actions of serotonin warrant further assessment in human AT, presumably akin actions acquired in humans could have major repercussions on obesity decreasing WAT and escalating BAT activity.
Glucocorticoids stimulated BAT Whitening by Stimulating Autophagy
The class of steroid hormones glucocorticoids and corticosterone are acknowledged to modulate glucose homeostasis in humans as well as rodents, respectively [92]. Hypercortisolism caused by either endogenous over generation of glucocorticoids or exogenous delivery of glucocorticoids as anti-inflammatory medication can stimulate the generation of obesity. A study by Gasparini, et al. [93], reported that corticosterone (250 mg/day) stimulates intracellular lipid accrual along with reduces diminished UCP1 expression in BAT of CD1 mice. Intriguingly, no significant alterations were found in PGC1α, FASN, or acetyl-coa carboxylase alpha (Acaca) expression, suggesting that alteration in lipid accrual did not directly implicate mitochondrial biogeneration , adipogenesis, or lipogenesis in BAT [94], Various robust synthetic glucocorticoids including dexamethasone have been generated for pharmacological use [89]. Like for instance, dexamethasone has been believed to be responsible in the study of adipogenesis [94]. Nevertheless, Deng, et al. [95], illustrated that exposing C57BL/6 mice to 5mg/kg dexamethasone for one week stimulated autophagy lipid droplet expansion in iBAT. Akin outcomes were observed in vitro using brown adipocytes ATG7 expression, in part by escalating the expression of B cell translocation gene 1 (BTG1) that induces the activity of CREB [96]. Consistently, UCP1 expression was down regulated, while the expression of WAT marker genes RB Despite, the remodelling of BAT as well as beige AT via whitening apparently is more frequent in obesity; clarification is required regarding how this maladaptive event takes place. Various investigations have revealed that the BAT/beige AT whitening has numerous factors involved in its origin. Actually, several factors like diet, age, genetics, thermo neutrality, along with chemical exposure have been illustrated to greatly impact the whitening of AT by targeting various modes (Figure 6).
transcriptional corepressor 1(Rb1), nuclear-receptor- interacting protein 1 (Nrip1), as well as Rbl1/p107 (RB transcriptional corepressor like 1) were up regulated [96]. Other proof revealed that chronic exposure to corticosterone can stimulate the whitening of BAT in C57BL/6 mice, was clarified by escalated adipocyte area along with escalated expressions of UCP1 in BAT [97]. Noticeably, the whitened phenotype has not been earlier correlated with scalated uncoupling proteins under chronic stress; nevertheless Bel, et al. [96], suggested that the increased UCP1 expression could be a compensatory mechanism under some stress.
Conclusions along with Future Directions
![Figure 6: Coutesy Summary of the proposed mechanisms by which multiple factors induce the whitening of brown (BAT) and beige adipose tissue. Factors such as high fat diet (HFD), bevacizumab, and nicotine induce the whitening by suppressing pro-angiogenic cytokines such as VEGF and VEGF-A, which inhibit angiogenesis and impair vascularization in BAT and beige adipose tissue, whereas thermoneutral zone and dexamethasone (Dex) exposure increase ATG7 and TFEB, which stimulate autophagy or mitophagy. Other factors such as aging, and elevated serotonin induce BAT/beige adipose tissue whitening by increasing INF-γ, and LncRNAs, and decreasing BMP-7, which in turn inhibits brown adipocyte differentiation. Moreover, dechlorane plus (DP) contributes to whitening in part by inhibiting UCP1 expression. ATG7, autophagy-related 7; LncRNAs, long non-coding RNAs;BMP-7, bone morphogenetic protein 7; BTG1, B cell translocation gene 1; IFN-g, interferon-gamma; UCP1, uncoupling protein 1; TFEB, transcription factor EB; VEGF, vascular endothelial growth factor [13].](/fulltextimages/10324/fig_6.png)
Figure 6: Coutesy Summary of the proposed mechanisms by which multiple factors induce the whitening of brown (BAT) and beige adipose tissue. Factors such as high fat diet (HFD), bevacizumab, and nicotine induce the whitening by suppressing pro-angiogenic cytokines such as VEGF and VEGF-A, which inhibit angiogenesis and impair vascularization in BAT and beige adipose tissue, whereas thermoneutral zone and dexamethasone (Dex) exposure increase ATG7 and TFEB, which stimulate autophagy or mitophagy. Other factors such as aging, and elevated serotonin induce BAT/beige adipose tissue whitening by increasing INF-γ, and LncRNAs, and decreasing BMP-7, which in turn inhibits brown adipocyte differentiation. Moreover, dechlorane plus (DP) contributes to whitening in part by inhibiting UCP1 expression. ATG7, autophagy-related 7; LncRNAs, long non-coding RNAs;BMP-7, bone morphogenetic protein 7; BTG1, B cell translocation gene 1; IFN-g, interferon-gamma; UCP1, uncoupling protein 1; TFEB, transcription factor EB; VEGF, vascular endothelial growth factor [13].
Besides, these factors, chronic exposure to greater quantities of particulate matter, a complicated mixture of solid along with liquid particles obtained from human activities as well as natural sources further facilitate the whitening of BAT, as reviewed by Guardia and Shin, et al. [98]. Despite, the activity of BAT has been robustly associated with the protection against obesity, fatty liver, along with T2D [99], the impairment or whitening of BAT in obesity, might aid in accelerating other metabolic complications [99]. For instance, recent proof has illustrated that robust hyperprolactinemia can further facilitate BAT whitening along with further aggravates HFD-induced metabolic dyscontrolling [100, 101]. This indicates that the whitening might portray one of the complications responsible for the pathogenesis of obesity; along with it can result in other secondary complications. Noticeably, experimental proof detailed here points that autophagy, inflammation, along with dysfunctional angiogenesis “vascularization” are central events involved in BAT/beige AT whitening [78, 85, 102, 103]. For instance, the most frequent mode believed to be responsible in AT whitening include the hampering of VEGFs, PGC1α, along with BMP-7 which result in dysfunctional vascularization, mitochondrial bio generation, along with brown adipocyte differentiation, respectively. This might result in greater sophisticated events like infiltration of INF-γ secreting T cells, escalated autophagy as well as, dysfunctional substrate metabolism. This proof is in agreement with the observation from gene knockout models, which pointed that the whitening of BAT is under the control of β-AR [52], BMP [101], mitochondrial transcription factor A [104, 105], as well as other genes controlling BAT function [106, 107, 108, 109, 110], as well as miRNAs that control multiple events including the differentiation as well as and function of brown adipocytes [111, 112], Dependent on the present validation, there are various probable marker genes that are implicated in controlling BAT/beige AT whitening . Nevertheless, this requires further assessment in both non-clinical as well as clinical settings. Specific attention must be placed regarding identification of probable therapeutic strategies to avoid or reverse AT whitening in obesity. This is inclusive of Fa evaluating in addition to gaining insight ins with regards to the therapeutic effects of prominent agents like metformin in targeting the AT to manage obesity- correlated complications [113].
References
-
Stern JH, Rutkowski JM, Scherer PR (2016) Adiponectin, leptin, and fatty acid in the maintenance of metabolic homeostasis through adipose tissue crosstalk. Cell Metab 23(5): 770-784.
-
Symonds ME, Pope M, Budge H (2015) The ontogeny of brown adipose tissue. Annu Rev Nutr 35: 295-320.
-
Yang J, Zhang H, Parhat K, Xu H, Li M, et al (2021) Molecular imaging of brown adipose tissue mass. Int J Mol Sci 22(17): 9436.
-
Virtanen KA, Icidell ME, Orava J (2009) Functional brown adipose tissue in healthy adults. N Engl J Med 360(15): 1518-1525.
-
Cypess AM, Lehman S, Williams G (2009) Identification and importance of brown adipose tissue in adult humans. N Engl J Med 360(15): 1509-1517.
-
Richard D, Picard F (2011) Brown fat biology and thermogenesis. Front Biosci 16(4): 1233-1260.
-
Wang GX, Zhao XY, Lin JD (2015) The brown fat secretome: molecular function beyond thermogenesis. Trends Endocrinol Metab 26(5): 231-237.
-
Saito M, Okamatsu Ogura Y, Matsushita M, Watanabe K, Yoneshiro T, et al. (2019) High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and obesity. Diabetes 58(7): 1526-1531.
-
Cannon B, Nedergaard J (2004) Brown adipose tissue: Function and physiological significance. Physiol Rev 84(1): 277-359.
-
Madden CJ, Morrison SF (2010) Endogenous activation of spinal 5hydroxy tryptamine receptors contributes to the thermoregulatory activation of brown adipose tissue. Am J Physiol Reg Integr Comp Physiol 298(3): R776-R783.
-
Villaroya F, Cereijo R, Villaroya J, Giralt M (2017) Brown adipose tissue as a secretory organ. Nat Rev Endocrinol 13(1): 26-35.
-
Stanford KI, Middlebeek RJW, Townsend KL (2013) Brown adipose tissue regulates glucose homeostasis and insuln sensitivity. J Clin Invest 123(1): 215-223.
-
Ziquby K, Dludia PV, Mithembu SXH, Nkambule BB, Mabhida SE, et al. (2023) An insight into Brown/ beige adipose tissue whitening, a metabolic complication of obesity. Front Endocrinol (Lausanne) 14: 1114767.
-
Kochar Kaur k, Allahbadia GN, Singh M (2015) Therapeutic Applications of the Recent Understanding of Brown or Beige Adipocyte Physiology. Adv Tech Biol Med 3(2).
-
Kaur KK, Allahbadia GN, Singh M (2018) Advances in BAT physiology for understanding and translating into Pharmacotherapies for obesity and comorbidities. MOJ Drug Des Develop Ther 2(5): 166-176.
-
Kaur KK, Allahbadia GN, Singh M (2015) An Update on Microrna’s and Metabolic Regulation with Future Therapeutic Potentials Regarding Diagnosis and Treatment of Obesity, Metabolic Syndrome and Other Related Disorders. J Health Med Informat 6(2): 1-16.
-
Kaur KK, Allahbadia GN, Singh M (2018) Utilization of β3 Adrenergic Receptors as Targets for Treating Diabesity-Mirabegron and Beyond A Systematic Review. International Journal of Innovations in Biological and Chemical Sciences 11: 1-13.
-
Kaur KK, Allahbadia GN, Singh M (2013) SGLT2 hampering agents possess effectiveness not only by their renal glycosuria action but possess ant obesity along with anti-insulin Resistance along with: Concentration on browning along with Macrophage Polarization: A Narrative Review. Accepted for publication.
-
Kaur KK, Allahbadia GN, Singh M (2013) Brown Adipose Tissue in the form of Innovative Approach for Polycystic ovary syndrome Treatment-still long time to reach Clinical Arena: A Narrative review under review.
-
Pillon NJ, Loos RJF, Marshall SM, Zierath JR (2021) Metabolic consequences of obesity and type 2 diabetes: Balancing genes and environment for personalized care. Cell 184(6): 1530-1544.
-
Nascimento GG, Peres MA, Mittinty MN, Peres KG, Do LG, et al. (2017) Diet-induced overweight and obesity and periodontitis risk: An application of the parametric g-formula in the 1982 pelotas birth cohort. Am J Epidemiol 185(6): 442-451.
-
Fabbri E, Zoli M, Gonzalez Freire M, Salive ME, Studenski SA, et al. (2015) Aging and multimorbidity: New tasks, priorities, and frontiers for integrated gerontological and clinical research. J Am Med Dir Assoc 16(8): 640- 647.
-
Konda PY, Poondla VK, Jaiswal KK, Dasari S, Uyyala R, et al. (2020) Pathophysiology of high fat diet induced obesity: impact of probiotic banana juice on obesity associated complications and hepatosteatosis. Sci Rep 10: 16894.
-
Kayode OO (2021) Diet and obesity. In: Psychology and pathophysiological outcomes of eating Intech Open.
-
Moura E Dias MD, Dos Reis SA, Da Conceição LL, Oliveira Sediyama CMN, Pereira SS, et al. (2021) Diet-induced obesity in animal models: points to consider and influence on metabolic markers. Diabetol Metab Syndr 13(1):32.
-
Lang P, Hasselwander S, Li H, Xia N (2019) Effects of different diets used in diet-induced obesity models on insulin resistance and vascular dysfunction in C57BL/6 mice. Sci Rep 9(1): 19556.
-
Herold J, Kalucka J (2021) Angiogenesis in adipose tissue: The interplay between adipose and endothelial cells. Front Physiol 11: 624903.
-
Corvera S, Gealekman O (2014) Adipose tissue angiogenesis: impact on obesity and type-2 diabetes. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 1842(3): 463-472.
-
Paavonsalo S, Hariharan S, Lackman MH, Karaman S (2020) Capillary rarefaction in obesity and metabolic diseases–Organ-Specificity and possible mechanisms. Cells 9(12): 2683.
-
Elias I, Franckhauser S, Bosch F (2013) New insights into adipose tissue VEGF-a actions in the control of obesity and insulin resistance. Adipocyte 2(2): 109-112.
-
Pellegata NS, Berriel Diaz M, Rohm M, Herzig S (2022) Obesity and cancer– extracellular matrix, angiogenesis, and adrenergic signaling as unusual suspects linking the two diseases. Cancer Metastasis Rev 41(3): 517-547.
-
Ye J (2011) Adipose tissue vascularization: its role in chronic inflammation. Curr Diabetes Rep 11(3): 203- 210.
-
Fujisaka S, Usui I, Ikutani M, Aminuddin A, Takikawa A, et al. (2013) Adipose tissue hypoxia induces inflammatory M1 polarity of macrophages in an HIF-1a-dependent and HIF-1a-independent manner in obese mice. Diabetologia 56(6): 1403-1412.
-
Gao M, Ma Y, Liu D (2015) High-fat diet-induced adiposity, adipose inflammation, hepatic steatosis and hyperinsulinemia in outbred CD-1 mice. PloS One 10(3): e0119784.
-
Dobner J, Ress C, Rufinatscha K, Salzmann K, Salvenmoser W, et al. (2017) Fat-enriched rather than high-fructose diets promote whitening of adipose tissue in a sex- dependent manner. J Nutr Biochem 49: 22-29.
-
Kuipers EN, Held NM, Het Panhuis WI, Modder M, Ruppert PMM, et al. (2019) A single day of high-fat diet feeding induces lipid accumulation and insulin resistance in brown adipose tissue in mice. Am J Physiol Endocrinol Metab 317(5): E820-E830.
-
Miranda CS, Silva-Veiga F, Martins FF, Rachid TL, Mandarim-De-Lacerda CA, et al. (2020) PPAR- activation counters brown adipose tissue whitening: acomparative study between high-fat– and high- fructose–fed mice. Nutrition 78: 110791.
-
Serdan TDA, Masi LN, Pereira JNB, Rodrigues LE, Alecrim AL, et al. (2021) Impaired brown adipose tissue is differentially modulated in insulin-resistant obese wistar and type 2 diabetic goto-kakizaki rats. Biomed Pharmacother 142: 112019.
-
Savva C, Helguero LA, González-Granillo M, Melo T, Couto D, et al. (2022) Maternal high-fat diet programs white and brown adipose tissue lipidome and transcriptome in offspring in a sex- and tissue-dependent manner in mice. International Journal of Obesity 46: 831-42.
-
Song NJ, Chang SH, Li DY, Villanueva CJ, Park KW (2017) Induction of thermogenic adipocytes: Molecular targets and thermogenic small molecules. Exp Mol Med 49(7): e353.
-
Rangel Azevedo C, Santana Oliveira DA, Miranda CS, Martins FF, Mandarimde Lacerda CA, et al. (2022) Progressive brown adipocyte dysfunction: Whitening and impaired no shivering thermogenesis as long-term obesity complications. J Nutr Biochem 105: 109002.
-
Parlee SD, MacDougald OA 92014) Maternal nutrition and risk of obesity in off spring: The Trojan horse of developmental plasticity. Biochim Biophys Acta Mol Basis Dis 1842(3): 495-506.
-
Sarker G, Litwan K, Kastli R, Peleg Raibstein D (2019) Maternal over nutrition during critical developmental periods leads to different health adversities in the off spring: relevance of obesity, addiction and schizophrenia. Sci Rep 9: 17322.
-
Lidell ME (2019) Brown adipose tissue in human infants. Handb Exp Pharmacol 251: 107-123.
-
Quesada López T, Gavaldà Navarro A, Morón Ros S, Campderrós L, IglesiasR, Giralt M, et al. (2019) GPR120 controls neonatal brown adipose tissue thermogenic induction. Am J Physiol Endocrinol Metab 317(5): 742- 750.
-
González Garcı́a I, Contreras C, Estévez Salguero Á, Ruı́z Pino F, Colsh B, et al. (2018) Estradiol regulates energy balance by ameliorating hypothalamic ceramide- induced ER stress. Cell Rep 25(2): 413-423.425.
-
Johnson F, Mavrogianni A, Ucci M, Vidal Puig A, Wardle J (2011) Could increased time spent in a thermal comfort zone contribute to population increases in obesity?. Obes Rev 12(7): 543-551.
-
Kern PA, Finlin BS, Zhu B, Rasouli N, McGehee RE, et al. (2014) The effects of temperature and seasons on subcutaneous white adipose tissue in humans: evidence for thermogenic gene induction. J Clin Endocrinol Metab 99(12): 2772-2779.
-
Yoneshiro T, Matsushita M, Nakae S, Kameya T, Sugie H, et al. (2016) Brown adipose tissue is involved in the seasonal variation of cold-induced thermogenesis in humans. Am J Physiol Regul Integr Comp Physiol 310(10): 999-1009.
-
Søberg S, Löfgren J, Philipsen FE, Jensen M, Hansen AE, et al. (2021) Altered brown fat thermoregulation and enhanced cold-induced thermogenesis inyoung, healthy, winter-swimming men. Cell Rep Med 2(10): 100408.
-
Fischer AW, Cannon B, Nedergaard J (2018) Optimal housing temperatures for miceto mimic the thermal environment of humans: An experimental study. Mol Metab 7: 161-170.
-
Kotzbeck P, Giordano A, Mondini E, Murano I, Severi I, et al. (2018) Brown adipose tissue whitening leads to brown adipocyte death and adipose tissue inflammation. J Lipid Res 59(5): 784-794.
-
Cui X, Nguyen NLT, Zarebidaki E, Cao Q, Li F, et al. (2016) Thermoneutralitydecreases thermogenic program and promotes adiposity in high-fat diet-fed mice.Physiol Rep 4(10): e12799.
-
Sass F, Schlein C, Jaeckstein MY, Pertzborn P, Schweizer M, et al. (2021) TFEBdeficiency attenuates mitochondrial degradation upon brown adipose tissue whitening at thermoneutrality. Mol Metab 47: 101173.
-
Kim D, Kim JH, Kang YH, Kim JS, Yun SC, et al. (2019) Suppression ofbrown adipocyte autophagy improves energy metabolism by regulating mitochondrial turnover. Int J Mol Sci 20(14): 3520.
-
Mancuso P, Bouchard B (2019) The impact of aging on adipose function and adipokine synthesis. Front Endocrinol (Lausanne) 10: 137.
-
Zoico E, Rubele S, de Caro A, Nori N, Mazzali G, et al. (2019) Brown and beige adipose tissue and aging. Front Endocrinol (Lausanne) 10: 368.
-
Mogilenko DA, Shchukina I, Artyomov MN (2021) Immune ageing at single-cell resolution. Nat Rev Immunol 22(8): 484-98.
-
Berry DC, Jiang Y, Arpke RW, Close EL, Uchida A, et al. (2017) Cellular aging contributes to failure of cold- induced beige adipocyte formation in old mice and humans. Cell Metab 25(1): 166-181.
-
Tajima K, Ikeda K, Chang HY, Chang CH, Yoneshiro T, et al. (2019) Mitochondrial lipoylation integrates age- associated decline in brown fat thermogenesis. Nat Metab 1: 886-898.
-
Lutz TA, Woods SC (2012) Overview of animal models of obesity. Curr Protoc Pharmacol 5: 5.6.1.
-
Gonçalves LF, Machado TQ, Castro Pinheiro C, de Souza NG, Oliveira KJ, et al. (2018) Ageing is associated with brown adipose tissue remodelling and loss of white fat browning in female C57BL/6 mice. Int J Exp Pathol 98(2): 100-108.
-
Pan XX, Yao KL, Yang YF, Ge Q, Zhang R, et al. (2021) Senescent T cell induces brown adipose tissue Whitening via secreting IFN-g. Front Cell Dev Biol 9: 637424.
-
Scambi I, Peroni D, Nodari A, Merigo F, Benati D, et al. (2022) The transcriptional profile of adipose-derived stromal cells (ASC) mirrors the whitening of adipose tissue with age. Eur J Cell Biol 101(2): 151206.
-
Du K, Bai X, Yang L, Shi Y, Chen L, et al. (2021) De novo reconstruction of transcriptome identified long non- coding RNA regulator of aging-related brown adipose tissue whitening in rabbits. Biol (Basel) 10(11): 1176.
-
Huang Z, Zhang Z, Moazzami Z, Heck R, Hu P, et al. (2022) Brown adipose tissue involution associated with progressive restriction in progenitor competence. Cell Rep 39(2):110575.
-
Darcy J, Tseng YH (2019) ComBATing ageing -does increased brown adipose tissue activity confer longevity?. Gero Science 41(3): 285-296.
-
Vatner DE, Ouydanich M, Zhang J, Campbell SC, Vatner SF (2023) Exercise enhancement of RGS14 disruption is mediated by Brown adipose tissue. Ageing Cell 22(4): e13791.
-
Vatner DE, Zhang J, Ouydanich M, Gueers J, Katsuoba E, et al. (2018) Enhanced longevity and metabolism by Brown adipose tissue with disruption of Regulator of G protein signaling14. Ageing Cell 17(4): e12751.
-
Loos RJF, Yeo GSH (2022) The genetics of obesity: from discovery to biology. Nat Rev Genet 23: 120-33.
-
Thaker VV (2017) Genetic and Epigenetic Causes of Obesity. Adolesc Med State Art Rev 28(2): 379-405.
-
Lapa C, Arias-Loza P, Hayakawa N, Wakabayashi H, Werner RA, et al. (2017) Whitening and impaired glucose utilization of brown adipose tissue in a rat model of type 2 diabetes mellitus. Sci Rep 7(1): 16795.
-
Amato AA, Wheeler HB, Blumberg B (2021) Obesity and endocrine-disrupting chemicals. Endocr Connect 10(2): 87-105.
-
Zhang J, Yang Y, Liu W, Schlenk D, Liu J (2019) Glucocorticoid and mineralocorticoid receptors and corticosteroid homeostasis are potential targets for endocrine-disrupting chemicals. Environ Int 133: 105133.
-
Pestana D, Teixeira D, Meireles M, Marques C, Norberto S, et al. (2017) Adipose tissue dysfunction as a central mechanism leading to dysmetabolic obesity triggered bychronic exposure to p,p’-DDE. Sci Rep 7(1): 2738.
-
Jin H, Li D, Wang X, Jia J, Chen Y, et al. (2018) VEGF and VEGFB playbalancing roles in adipose differentiation, gene expression, and function. Endocrinology 159(5): 2036-2049.
-
Zayek M, Parker K, Rydzewska M, Rifai A, Bhat R, et al. (2021) Bevacizumab for retinopathy of prematurity: 2-year neurodevelopmental follow-up. Am J Perinatol 38(11): 1158-1166.
-
Jo DH, Park SW, Cho CS, Powner MB, Kim JH, et al. (2015) Intravitreally injected anti-VEGF antibody reduces brown fat in neonatal mice. PloS One 10(7): e0134308.
-
Mishra A, Chaturvedi P, Datta S, Sinukumar S, Joshi P, et al. (2015) Harmful effects of nicotine. Indian J Med Paediatric Oncol 36(1): 24-31.
-
Riedel C, Schönberger K, Yang S, Koshy G, Chen YC, et al. (2014) Parental smoking and childhood obesity: Higher effect estimates for maternal smoking in pregnancy compared with paternal smoking-a meta-analysis. Int J Epidemiol 43(5): 1593-1606.
-
Zhang WX, Chen HJ, Fan J, Li GL, Sun A, et al. (2019) The association between maternal nicotine exposure and adipose angiogenesis in female rat off spring: A mechanism of adipose tissue function changes. Toxicol Lett 318: 12-21.
-
Martı́nez de Morentin PB, Whittle AJ, Fernø J, Nogueiras R, Diéguez C, et al. (2012) Nicotine induces negative energy balance through hypothalamic AMP-activated protein kinase. Diabetes 61(4):807-817.
-
Seoane Collazo P, Liñares Pose L, Rial Pensado E, Romero Picó A, Moreno Navarrete JM, et al. (2019) Central nicotine induces browning through hypothalamic κ opioid receptor. Nat Commun 10(1): 4037.
-
Fan J, Ping J, Zhang WX, Rao YS, Liu HX, et al. (2016) Prenatal and lactation nicotine exposure affects morphology and function of brown adipose tissue in male rat off spring. Ultrastruct Pathol 40(5): 288-295.
-
Chen HJ, Li GL, Zhang WX, Fan J, Hu L, et al. (2020) Maternal nicotine exposure during pregnancy and lactation induces brown adipose tissue whitening in female offspring. Toxicol Appl Pharmacol 409: 115298.
-
Mohajer N, Du CY, Checkcinco C, Blumberg B (2021) Obesogens: How they are identified and molecular mechanisms underlying their action. Front Endocrinol (Lausanne) 12: 780888.
-
Peshdary V, Calzadilla G, Landry A, Sorisky A, Atlas E (2019) Dechlorane plus increases adipogenesis in 3T3- L1 and human primary preadipocytes independent of peroxisome proliferator-activated receptor g transcriptional activity. Int J Obes 43: 545-555.
-
Winn NC, Vieira Potter VJ, Gastecki ML, Welly RJ, Scroggins RJ, et al. (2017) Loss of UCP1 exacerbates Western diet- induced glycemic dysregulation independent of changes in body weight in female mice downloaded from. Am J Physiol Regul Integr Comp Physiol 312(1): 74-84.
-
Winn NC, Grunewald ZI, Gastecki ML, Woodford ML, Welly RJ, et al. (2017) Deletion of UCP1 enhances ex vivo aortic vasomotor function in female but not male mice despite similar susceptibility to metabolic dysfunction. Am J Physiol Endocrinol Metab 313(4): 402-412.
-
Oh CM, Namkung J, Go Y, Shong KE, Kim K, et al. (2015) Regulation of systemic energy homeostasis by serotonin in adipose tissues. Nat Commun 6: 6794.
-
Rozenblit Susan S, Chapnik N, Froy O (2018) Serotonin prevents differentiation into brown adipocytes and induces trans differentiation into white adipocytes. Int J Obes 42: 704-710.
-
Luijten IHN, Cannon B, Nedergaard J (2019) Glucocorticoids and brown adipose tissue: Do glucocorticoids really inhibit thermogenesis?. Mol Aspects Med 68: 42-59.
-
Gasparini SJ, Swarbrick MM, Kim S, Thai LJ, Henneicke H, et al. (2019) Androgens sensitise mice to glucocorticoid- induced insulin resistance and fat accumulation. Diabetologia 62(8): 1463-1477.
-
Zilberfarb V, Siquier K, Strosberg AD, Issad T (2001) Effect of dexamethasone on adipocyte differentiation markers and tumour necrosis factor-alpha expression in human PAZ6 cells. Diabetologia 44(3): 377-386.
-
Deng J, Guo Y, Yuan F, Chen S, Yin H, et al. (2010) Autophagy inhibition prevents glucocorticoid-increased adiposity via suppressing BAT whitening. Autophagy 16(3): 451-465.
-
Bel JS, Tai TC, Khaper N, Lees SJ (2022) Chronic glucocorticoid exposure causes brown adipose tissue whitening, alters whole-body glucose metabolism and increases tissue uncoupling protein-1. Physiol Rep 10(9): e15292.
-
Zilberfarb V, Siquier K, Strosberg AD, Issad T (2001) Effect of dexamethasone on adipocyte differentiation markers and tumour necrosis factor-alpha expression in human PAZ6 cells. Diabetologia 44: 377-386.
-
Della Guardia L, Shin AC (2022) White and brown adipose tissue functionality isimpaired by fine particulate matter (PM 2.5) exposure. J Mol Med (Berl) 100: 665-676.
-
Wibmer AG, Becher T, Eljalby M, Crane A, Andrieu PC, et al. (2021) Brownadipose tissue is associated with healthier body fat distribution and metabolic benefits independent of regional adiposity. Cell Rep Med 2(7):100332.
-
AlcaláM, Calderon Dominguez M, Serra D, Herrero L, Viana M (2019) Mechanisms of impaired brown adipose tissue recruitment in obesity. Front Physiol 10: 94.
-
Lopez Vicchi F, de Winne C, Ornstein AM, Sorianello E, Toneatto J, et al. (2022) Severe hyperprolactinemia promotes brown adipose tissue whitening and aggravates high fat diet induced metabolic imbalance. Front Endocrinol (Lausanne) 13: 883092.
-
Lu X, Altshuler Keylin S, Wang Q, Chen Y, Sponton CH, et al. (2018) Mitophagy controls beige adipocyte maintenance through a parkin-dependent andUCP1- independent mechanism. Sci Signal 11(527): eaap8526.
-
Altshuler Keylin S, Shinoda K, Hasegawa Y, Ikeda K, Hong H, et al. (2016) Beige adipocyte maintenance is regulated by autophagy-induced mitochondrial clearance. Cell Metab 24: 402-419.
-
Blázquez-Medela AM, Jumabay M, Rajbhandari P, Sallam T, Guo Y, et al. (2019) Noggin depletion in adipocytes promotes obesity in mice. Mol Metab 25: 50- 63.
-
Vernochet C, Damilano F, Mourier A, Bezy O, Mori MA, et al. (2014) Adipose tissue mitochondrial dysfunction triggers a lipodystrophic syndrome with insulin resistance, hepatosteatosis, and cardiovascular complications. FASEB J 28(10): 4408-19.
-
Cannavino J, Shao M, An YA, Bezprozvannaya S, Kim J, et al. (2021) Regulation of cold-induced thermogenesis by the RNA binding protein FAM195A. Proc Natl Acad Sci USA 118(23): e2104650118.
-
Ahmadian M, Liu S, Reilly SM, Hah N, Fan W, et al. (2018) ERRg preserves brown fat innate thermogenic activity. Cell Rep 22(11): 2849-2859.
-
Cui J, Pang J, Lin YJ, Gong H, Wang ZH, et al. (2017) Adipose-specific deletionof Kif5b exacerbates obesity and insulin resistance in a mouse model of diet- inducedobesity. FASEB J 31(6): 2533-2547.
-
Silvester AJ, Aseer KR, Yun JW (2018) Ablation of DJ-1 impairs brown fat function indiet-induced obese mice. Biochimie 154:107-118.
-
Ahmadian M, Abbott MJ, Tang T, Hudak CSS, Kim Y, et al. (2011) Desnutrin/ATGL is regulated by AMPK and is required for a brown adiposephenotype. Cell Metab 13(6): 739-748.
-
Mori MA, Thomou T, Boucher J, Lee KY, Lallukka S, et al. (2014) Altered miRNA processing disrupts brown/ white adipocyte determination and associates with lipodystrophy. J Clin Invest 124(8): 3339-3351.
-
Lou P, Bi X, Tian Y, Li G, Kang Q, et al. (2021) MiR- 22 modulates brown adipocyte thermogenesis by synergistically activating the glycolytic and mTORC1 signaling pathways. Theranostics 11(8) :3607-3623.
-
Ziqubu K, Mazibuko-Mbeje SE, Mthembu SXH, Mabhida S, Jack BU, Nyambuya TM, et al. (2023) Anti- obesity effects of metformin: A scoping review evaluating the feasibility of brown adipose tissue as a therapeutic target. Int J Mol Sci 24(3): 2227.
- The Muculent Bleb-Mucinous Cystic Neoplasm-Hepatobiliary Region
- Insulin Sensitizers as Anti-Aging Agents: Unveiling Synergies with Albumin, GLP-1RA, Klotho Protein, and Metformin in the Quest to Combat Aging
- Reprogramming of GLP-1 Response at Prediabetes for the Prevention of Type 2 Diabetes: The Role of Albumin and GLP-1 Receptor Agonists
- The Mingled Allies-Combined Hepatocellular Carcinoma and Cholangiocarcinoma
- Compilation and Embodiment-Leydig Cell Tumour Testis
- Glucolipotoxicity: A Novel Different Perspective on the Causes of Cancer