Role of α-1 Anti Trypsin, A Constitutive Form of Nitric Oxide Synthase in Cyclic AMP Mediated Inhibition of Platelet Aggregation
Introduction: As reported before, the treatment of platelets with all currently known platelet aggregating agents was resulted in the reduction of nitric oxide (NO) synthesis through the inhibition of enzyme nitric oxide synthase (NOS). This reduction of NO in consequence released arachidonic acid from the platelet membrane ultimately leading to the aggregation of platelets. Aim: Experiments were carried out to identify the platelet NOS. Results: The amino acid sequence of the protein sample thus prepared was determined by mass spectrometry in which the platelet cytosolic NOS was identified to be α1 anti trypsin (AAT) when matched with the protein databank. The incubation of NOS/AAT with l-arginine resulted in the increase of NO synthesis to ~1nmol/h (P
Introduction
α1 anti-trypsin (AAT), one of the serine proteinase inhibitors known as SERPIN, has been reported to be associated with chronic obstructive pulmonary diseases (COPD) [1] in that the deficiency of AAT is well known to be associated with the pathogenesis of the development of emphysema due to tobacco smoking [2]. In recent years, the deficiency of AAT has been implicated in various diseases ranging from rheumatoid arthritis [3, 4] coronary atherosclerosis [5] to cancer [6, 7, 8, 9]. The aggregation of platelets has a dual role; on one hand it is essential for life-saving process of blood coagulation [10]. And, on the other hand excessive platelet aggregation on the site of a atherosclerotic plaque rupture or fissuring on the coronary artery reported to cause formation of thrombus (a microaggregate of platelets embedded in fibrin mass) The formed thrombus thus cause blockade of normal circulation in the heart resulting in acute coronary syndrome (ACS) which is a major killer of human race [11]. There are several agents including insulin [12], prostacyclin [13], estriol [14] and acetyl salicylic acid (aspirin) [15] that inhibit aggregation of platelets. While insulin and estriol have been reported to inhibit platelet aggregation through nitric oxide (NO) synthesis [12, 14], prostacyclin has been reported to inhibit platelet aggregation through the cellular increase of cyclic AMP (cAMP) [16]. On the other hand, aspirin has been reported to inhibit platelet aggregation through the inhibition of platelet cycloxygenase (COX) [17, 18]. This compound has also been reported to stimulate NO synthesis in different cells including platelets and is known to inhibit platelet aggregation [19]. Although, the aggregation of platelets by different aggregating agents has been reported to be mediated through the synthesis of prostaglandins and thromboxanes due to the activation of COX [20], the mechanism for the release of arachidonic acid from the platelet membrane phospholipids by the aggregating agents remains obscure. Furthermore, the mechanism of inhibition of platelet aggregation induced by the increase of cellular cAMP level also remains obscure. We have recently found that all platelet aggregating agents both natural or synthetic inhibited NO synthesis in platelets that in consequence resulted in the liberation of arachidonic acid from the platelet membrane phospholipids due to the activation of phospholipase A2 [21]. To find out the nature and properties of the platelet cytosolic nitric oxide synthase (NOS) that was found to be inhibited by the platelet aggregating agents, the enzyme (NOS) was purified to homogeneity from the platelet cytosolic fraction. We herein report for the first time ever, that the identified NOS were actually α-1 anti trypsin which was not only inhibited by different aggregating agents but also found to be stimulated by aspirin.
Methods & Materials
Ethical Approval
The study used blood samples from both normal male and female volunteers. Appropriate approval was obtained from the Internal Review Board, Sinha Institute of Medical Science and Technology, Kolkata. All selected volunteers signed informed consent form before they were asked to donate blood.
Chemicals & Supplies
ADP, collagen, thrombin, l-epinephrine, NG-methyl-l- arginine acetate ester (l-NAME) and the goat anti-rabbit immunoglobulin G-alkaline phosphatase, cAMP and its antibody were the products of Sigma Chemical, St Louis, MO, USA. Dermcidin isoform-2 was prepared as described before [22]. Aspirin was obtained from Medica Zydus Healthcare. Maxisorp plates for the enzyme linked immunosorbent assay (ELISA) were the products of Nunc, Roskilde, Denmark. The chemicals used in the study were of analytical grade. All aggregating agents and inhibitors of aggregation were dissolved in 0.9% NaCl and neutralized to pH 7.0 at 00C and discarded after use.
Selection of normal volunteers
Both male and female volunteers (M=30, F=10, n=40) between the ages of 30 to 45 years old participated in the study. None of the selected participants had systemic hypertension or diabetes mellitus. None of the volunteers had life threatening infection, suffered cardiovascular, cerebrovascular diseases and had been hospitalized for any reason at least for 6 months before they were requested to donate blood. All participating volunteers were asked to stop taking any medication including aspirin for at least 4 weeks before donating blood. No female volunteers had ever used any contraceptive medications.
Collection of Blood and the Preparation of Platelet-Rich-Plasma (PRP)
Peripheral blood (50mL) was collected from both male or female volunteers by venipuncture using 19- gauge siliconized needle and collected in plastic vials, anticoagulated by adding 1vol of sodium citrate to 10 vol of blood (final concentration of sodium citrate was 13mM), and mixed by gentle inversion. The
anticoagulated blood was centrifuged at 200g for 15min at room temperature (230C). After centrifugation PRP in the supernatant was collected [23].
Preparation of Washed Platelet Suspension
PRP in 1.0mM EDTA was centrifuged at 2000g for 20 min at 230C, and the platelet pellet thus obtained was washed 3 times by centrifugation after resuspending the pellet in Kreb’s buffer, pH 7.4, containing 121mM NaCl, 3mM KCl, 10.0mM Glucose, 0.4mM KH2PO4, 0.24mM MgSO4, 1.73mM CaCl2, 25mM NaHCO3 in 250 ml distilled water. After the final wash the pellet was resuspended and washed 3 more times in the same buffer without EDTA containing 2.0mM CaCl2.
Preparation of aspirin solution
Aspirin was dissolved in deionised water. The pH was adjusted to 7.0 by adding 0.1M NaHCO3 just before use and was discarded after use.
Aggregation of platelets
Unless otherwise stated, the aggregation of platelets in PRP was performed by incubating the PRP with different aggregating agents which includes 4.0μM ADP, 5μM epinephrine, 2μg/ml collagen, 1U/ml thrombin or 0.1μM dermcidin, 0.1mM l-NAME and was studied by using an aggregometer as described before [23]. In separate experiments, PRP was pre-treated with 80µM aspirin for 30 mins and after incubation aggregation of platelets was initiated by adding different aggregating agents as indicated above.
Purification of a Novel Nitric Oxide Synthase (NOS) From the Platelet Cytosolic Fraction
The platelets were isolated from blood sample of normal volunteers and were washed as described above. The washed platelets were then disrupted by freez-thaw in liquid N2. The cytosolic fraction was obtained by separating the supernatant from the membrane fraction by ultracentrifugation at 60,000g for 30min at 00C. When the cytosolic fraction was electrophoresed using SDS-PAGE, a novel protein band of Mr. 47 kDa was found to be present in the SDS gel. This protein band of Mr 47 kDa was next excised from the gel, triturated in 0.9% NaCl, and clarified by centrifugation at 10 000g at 00C. The clarified sample was reelectrophoresed on polyacrylamide gel in the absence of SDS. The staining of the gel demonstrated the presence of a single band.
Determination of Nitric Oxide (NO) Synthesis of the Gel Bands in SDS-PAGE
The unstained gel was cut in 1mm each from which a total of 70 gel bands were obtained. Each gel bands were pooled, triturated in 0.9% NaCl and dialyzed by Poly-ethylene glycol method. The NO synthesis of each protein bands were performed in presence of l-arginine using methhemoglobin method by determining the absorption maxima at 575nm and 600nm in a Beckman Spectrophotometer Model DU under N2 [24, 25]. The synthesis of NO was independently verified by chemiluminescence method [25]. The platelet count was determined by optical microscopy. In separate experiments, NO synthesis in the isolated 47kDa protein band in presence of different aggregating agents were performed by incubating the clarified sample prepared as described above with different aggregating agents which includes 4.0μM ADP, 5μM epinephrine, 2μg/ml collagen, 1U/ml thrombin or 0.1μM dermcidin, 0.1mM l- NAME. The NO synthesis was determined as described above by methhemoglobin method.
Amino Acid Sequencing of the Isolated NOS
The 47 kDa band in an identical but unstained gel was excised out and the final gel slices were washed twice with 50% high-performance liquid chromatography grade acetonitrile in water for 2-3min with gentle shaking and discarding the supernatant after each wash. The gel bands were then made fully dried using a lyophilizer. The amino acid sequence of the protein sample thus prepared was determined by Mass Spectrometry and Proteomic Resource Core, Harvard University using microcapillary reverse-phase HPLC nanoelectrospray tandem mass spectrometry (μLC/MS/MS) on a Thermo LTQ Orbitrap mass spectrometer. The identity of the protein of Mr 47kDa was determined by protein database matching [26, 27, 28].
Scatchard Plot of the Equilibrium Binding of Camp to AAT
The purified NOS/AAT (0.1mg) was incubated with different concentrations of cAMP for 60 min (optimal time required for the maximal binding of cAMP to purified protein determined in separate experiments). After incubation, 0.5 mL of incubation mixture was filtered over micro glass fibre membrane filter (GF/C, Sigma alrich) by using Millipore filtration unit as described before [29]. The use of GF/C membrane filtration allowed the protein to remain adhered to the filter and allowing the free cAMP passed through the filtrate under mild vacuum [30]. The membrane filter was washed with the 3 vol of the reaction buffer. After washing, the membrane filter was air dried and the bound cAMP eluted from the filter by washing the filter with 0.1% TritonX-100. The filtrate was used to determine the amount of cAMP present in the filtrate by ELISA using cAMP antibody. Scatchard plot of the equilibrium binding characteristics to the platelets was constructed and the dissociation constant (Kd), and the cAMP binding number (n) on the platelet surface was calculated.
Removal of Bound cAMP by Dialysis
The bound cAMP from the NOS/AAT was removed by continued dialysis using Himedia dialysis membrane-60 at 20C kept in 0.9% NaCl buffer for 24h. Following dialysis the reaction mixture was treated with different aggregating agents and NO synthesis was measured as described above.
Statistical Analysis
Results shown are mean ± standard deviation (S.D). The significance of the results was determined by Student’s t-test. The significance “P”<0.01 was considered to be significant.
Results
Identification of Novel NOS from the Platelet Cytosol
When the platelet cytosol prepared by disrupting the whole washed platelets in liquid nitrogen as described in Materials and Methods was electrophoresed on SDS- polyacrylamide gel and when the gel was stained using Coomassie blue, the presence of a protein band of Mr 47 kDa was found (Figure 1, Panel-A, Lane 2). When the Mr 47 kDa protein band was excised from the gel, triturated in 0.9% NaCl, clarified, dialyzed and reelectrophoresed on polyacrylamide gel in the absence of SDS, a single protein band stained by coomassie blue was found in the gel (Figure 1, Panel-A, Lane 3). In an identical gel (Panel-A, Lane 2) which was not stained with the dye to avoid denaturation was cut into slices (Each 1mm). Each slice was separately triturated in 0.9% NaCl and centrifuged as described in Methods and Materials and the NO synthesis was measured in the clarified supernatant. The result demonstrated the maximum synthesis of NO from l-arginine (the substrate of NOS) from the 47kDa band of interest indicating that the isolated protein was a constitutive form of NOS (Figure 1, Panel-B). Additionally, incubation of the isolated gel with aspirin also stimulated the NO synthesis activity of the protein.
Effect of Different Aggregating Agents on the Enzymic Activity of NOS Isolated from the Platelet Cytosol
When the isolated NOS from the platelet cytosol was treated with different platelet aggregating agents and the synthesis of NO from l-arginine was studied, it was found that the presence of 4µM ADP, 2µg/ml collagen, 1.0U/ml thrombin, 5µM l-epinephrine, 0.1µM dermcidin and 0.1mM l-NAME in the reaction mixture resulted in the inhibition of NOS activity by 84.5%, 75.8%, 90.3%, 71%, 69.1% and 80.6% respectively (p<0.01) (Figure 2, Panel-A). The decrease in the NOS activity resulted in the increase of aggregation of platelets in PRP as shown in Figure 3. Corollary to the results, the inference drawn indicate that the increase of NO would result in the inhibition of aggregation of platelets. This possibility was tested by pre-incubating the isolated NOS with pure NO solution (0.8nM) in 0.9% NaCl and subsequently treated with different aggregating agents (Figure 2, Panel-B). In control experiments, only equal volume of 0.9% NaCl was used. The results indicated the failure of aggregating agents to inhibit NO synthesis in presence of pure NO. Additionally, in all cases, the treatment of PRP with NO solution in 0.9% NaCl and initiating aggregation with aggregating agents resulted in the inhibition of platelet aggregation (Figure 3). Similarly, NO synthesis was also measured in platelets treated with different aggregating agents and pure 0.8nM NO solution as described in Materials and Methods.
The Amino Acid Sequence of the Electrophoretically Purified NOS from the Platelet Cytosol
The amino acid sequence of isolated NOS was determined as described in Materials and Methods. The protein database matching demonstrated that the isolated cytosolic protein was α-1 anti trypsin that showed NOS activity, Uniprot ID: P01009 comprising of 418 amino acids (Figure 4).
Determination of Binding Capacity of Electrophorectically Purified NOS/AAT With Camp
The increase of cAMP in platelets has been reported to result in the inhibition of platelet aggregation induced by different aggregating agents [31, 32]. As the binding of cAMP to the target protein (e.g. protein kinase) has been reported to be essential for the expression of the nucleotide effect, the possibility of cAMP binding to the isolated NOS/AAT was carried out. The incubation of cAMP to the isolated protein resulted in the saturable binding profile of the nucleotide to the protein was obtained (Figure 5). Scatchard plot of the equilibrium binding of cAMP to the NOS produced a linear plot (Figure 6) indicating homogenous binding sites population of the cyclic nucleotide on the NOS/AAT protein molecule. The analysis of binding characteristics of the plot demonstrated the dissociation constant, (Kd) was 15.54nM with 1.56 X 103 molecules of cAMP bound to per molecule of the protein.
Effect of Binding of NOS/AAT to Camp on the NOS Activity of the Purified Protein
The treatment of cAMP bound NOS with different platelet aggregating agents showed that the nucleotide conferred resistance to the inhibitory effect of different aggregating agents on the inhibition of the NOS activity compared to appropriate controls (Figure 7). In contrast, the binding of cAMP to NOS/AAT did not result in the inhibition of NO synthesis ability of the enzyme. The removal of the bound cAMP by continued dialysis fully restored the ability of different aggregating agents to inhibit the NOS activity of the isolated protein (Figure 7).
Discussion
These results demonstrated that the aspirin induced inhibition of platelet aggregation is not necessarily mediated only through the inhibition of COX due to the inhibition of prostaglandin synthesis [20]. These results suggests that aspirin, the well known inhibitor of platelet aggregation may also inhibit platelet aggregation independent of COX through the synthesis of NO catalyzed by NOS/AAT instead of the inhibition of prostaglandin synthesis. The aggregation of platelets by different aggregating agents through the involvement of COX pathway is reported to be a central event both in the life-saving process of normal blood coagulation and in the extremes of life threatening thrombus formation in the acute coronary artery syndromes [10]. Currently, the regulation of platelet aggregation itself with the involvement of the COX pathway, particularly in the context of inhibition of aggregation induced by platelet aggregating agent like nitric oxide or prostacyclin through the increase of cellular cAMP level remains an unsettled issue. We have reported before that, all aggregating agents, man-made or natural, decreased basal NO level in platelets [21], which suggested that the inhibition of nitric oxide synthase (NOS) by different aggregating agents was critically involved in the aggregation of platelets by the aggregating agents through COX pathway [21]. As COX itself has no NOS activity, in an effort to identify the NOS, the enzyme from the cytosolic fraction that could be inhibited by the aggregating agents was purified to homogeneity. It was unexpectedly found that the NOS in question, in reality, were actually α-1 antitrypsin as identified by the amino acid sequence analysis of the protein (Figure 4). It should be mentioned here that, α-1 antitrypsin , a member of the SERPIN superfamily of serine protease inhibitor [1] is well known for its role in the pathogenesis of COPD, particularly in emphysema which is reported to be pathologically related to the tobacco smoke induced condition [2]. That the α-1 antitrypsin in platelets is a form of constitutive NOS was evidenced by the stimulation of the synthesis of NO (Figure 1, Panel-B) catalyzed by the electrophoretically purified α-1 antitrypsin (Figure 1, Panel-A) that used l- arginine as its substrate [33].
Furthermore, as in the case of inhibition of NOS activity by different aggregating agents in intact platelets [21], the NOS activity of the purified α-1 antitrypsin was also inhibited by all known aggregating agents (Figure 2, Panel-A). Corollary to the inference drawn from the results implied that in the aggregation of platelets by different aggregating agents, the inhibition of NO synthesis in platelets was necessary. The increase of NO synthesis by pure NO solution in 0.9% NaCl were capable of inhibiting platelet aggregation induced by the aggregating agent (Figure 2, Panel-B). Although the increase of cAMP in the platelets are known to inhibit platelet aggregation [31, 32], the mechanism of cAMP induced inhibition of platelet aggregation currently remains obscure. No evidence is currently available to suggest that the binding of cAMP to any particular platelet protein results in the inhibition of aggregation induced by platelet aggregating agents. In-silico analyses of binding of cAMP to the purified protein NOS/AAT demonstrated hydrogen bonding interaction with Serine 56 residue according to docking results obtained in Hex software. In contrast, when the serine 56 residue was mutated and the docking was re-performed, no interactions took place. As the difference in minimum free energy (∆G) between the wild and mutated type was ˃2kcal/mol, the serine residue was considered to be a ‘hot spot’ for interactions [34]. Based on these in-silico analyses, in-vitro analyses of binding of cAMP to NOS/AAT demonstrated that the purified α-1 anti-trypsin was capable of binding cAMP with a saturable profile (Figure 5). Scatchard plot of the equilibrium binding of cAMP to α-1 anti-trypsin showed that the dissociation constant Kd= 15.54nM with 1.56 X 103 molecules of cAMP bound to per molecule of α-1 anti-trypsin (Figure 6). Conversely, when the bound cAMP in the α-1 anti- trypsin was removed by dialysis, the α-1 anti-trypsin was found to be inhibited by the aggregating agents (Figure 7). Interestingly, the binding of cAMP to α-1 anti-trypsin did not prevent the activation of the NOS activity of α-1 anti-trypsin induced by aspirin. In this context, it could be suggested that if the binding of cAMP to NOS/AAT would have impaired the systemic NO synthesis, it could be a contradictory/counter- productive event in many physiologic/metabolic processes. As NO not only inhibits platelet aggregation, this ‘biologic messenger’ molecule is also reported to be involved in thrombolysis [19], in the control of arterial hypertension [35] and also in the synthesis of insulin [36]. The cyclic nucleotide on the other hand is also known to have numerous role as a ‘messenger molecule’ in many metabolic and physiologic process [37] including that in the β-adrenergic stimulation of the cardiac cells [37]. As suggested by the results presented above, the role of cAMP on the enzymic activity of NOS/AAT was a finely tuned physiologic process in the aggregation of blood platelets by aggregating agents and in the cAMP induced inhibition of platelet aggregation without interference to each other’s biologic functions.
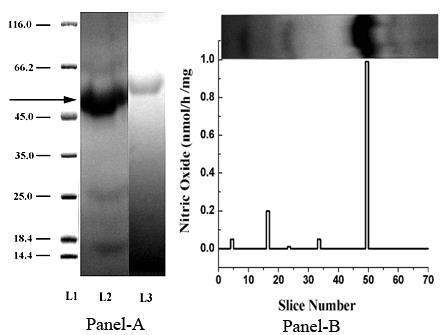
Figure 1: Presence of a novel protein from platelet cytosolic fraction and its effect on NO synthesis. Panel A- Platelet cytosolic fraction was isolated from washed platelets by freez-thaw method as described in Materials and Methods. SDS-polyacrylamide gel electrophoresis of proteins present in platelet cytosolic fraction as shown. L (Lane) 1 represents the standard base pair protein markers of known molecular weight (kDa), L2 shows the presence of proteins present in the platelet cytosolic fraction, L3 shows PAGE of the protein band in the L2 (black arrow) that was excised out and reelectrophoresed in the absence of SDS. The arrow indicates the band of interest whose NOS activity was measured as shown in Panel B. Panel B- The NO synthesis of each protein bands obtained as described in Materials and Methods were performed in presence of l-arginine using methhemoglobin method. Control was a portion of the gel of similar area without any protein band that was cut out from the gel and similarly treated with 0.9% NaCl as in the case of the identified 47 kDa protein from the gel.
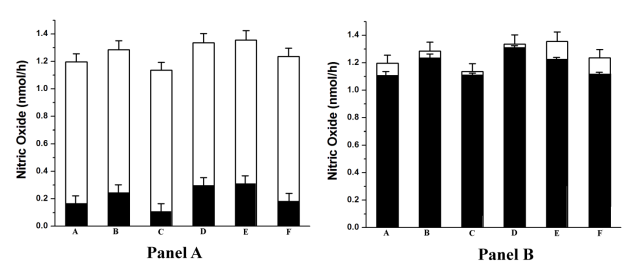
The protein band was excised out of the gel and triturated and clarified by centrifugation as described in Materials and Methods. The clarified supernatant was dialyzed and used to study the NOS activity in presence of different aggregating agents (Panel-A) and in the presence of pure NO (Panel-B). □ portion indicates NO synthesis in control. ■ portion indicates NO synthesis in the purified band treated with different aggregating agents (Panel-A) and pure NO (Panel-B). A= 4µM ADP; B= 2µg/ml collagen; C= 5µM thrombin; D= 5µM l-epinephrine; E= 0.1mM l-NAME; F= 0.1µM dermcidin isoform 2, pure NO= 0.8nM.
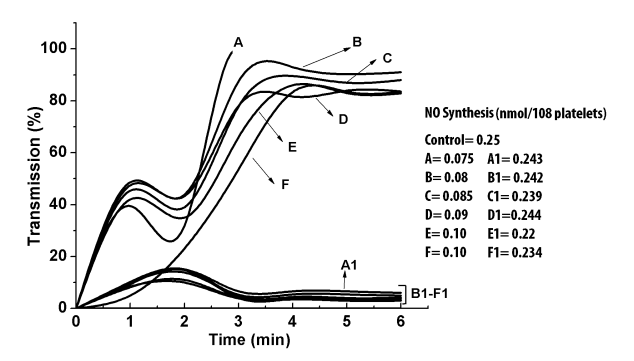
Figure 3: Aggregation profile and synthesis of NO in presence of aggregating agents and pure NO. Different aggregating agents including ADP, epinephrine, collagen, thrombin and dermcidin and l- NAME was added to the washed PRP and the aggregation of platelets at different time intervals was studied as shown: Curve A= aggregation of PRP in case of 5µM thrombin , Curve A1= aggregation of PRP in case of 5µM thrombin and 80µM aspirin, Curve B= aggregation of PRP in case of 2µM ADP , Curve B1= aggregation of PRP in case of 2µM ADP and 80µM aspirin, Curve C= aggregation of PRP in case of0.1µM dermcidin , Curve C1= aggregation of PRP in case of 0.1µM dermcidin and 80µM aspirin, Curve D= aggregation of PRP in case of 5µM epinephrine, Curve D1= aggregation of PRP in case of 5µM epinephrine and 80µM aspirin, Curve E= aggregation of PRP in case of 0.1mM l-NAME, Curve E1= aggregation of PRP in case of 0.1mM l-NAME and 80µM aspirin, Curve F= aggregation of PRP in case of 2µg/ml collagen, Curve F1= aggregation of PRP in case of 2µg/ml collagen and 80µM aspirin. The upward increase of the transmission in curve A was due to the clotting of the washed PRP induced by thrombin. The figure is a typical representative of at least 10 different experiments using blood from both male and female volunteers (M=5, F=5).
Figure 4: Amino acid sequence of α1 anti-trypsin. Standard one letter abbreviation of the amino acids are: G glycine, P proline, A alanine, V valine, L leucine, I isoleucine, M methionine, C cysteine, F phenylalanine, Y tyrosine, W tryptophan, H histidine, K lysine, R arginine, Q glutamine, N asparagine, E glutamic acid, D aspartic acid, S serine, T threonine
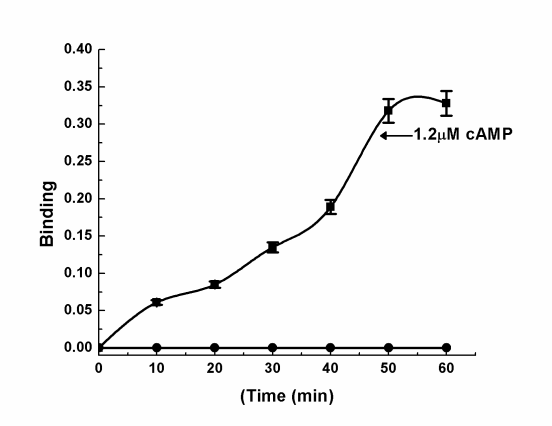
Figure 5: Equilibrium binding of cAMP to the purified NOS/AAT. The 47 kDa purified protein (0.1mg) in 0.9% NaCl was incubated with 1.2µM cAMP for different times as indicated. After incubation, amount of cAMP present in the supernatant was determined by spectrophotometer at 405nm absorbance spectra. Each point is mean±SD of atleast 5 individual experiments using purified NOS/AAT.
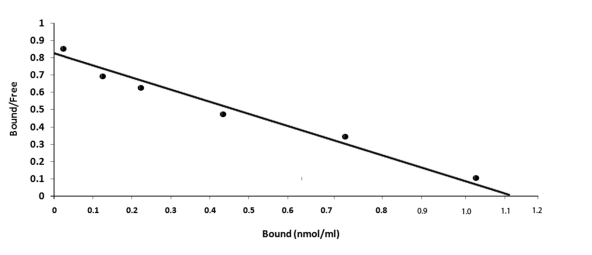
Figure 6: Scatchard plot analysis of the equilibrium binding of cAMP to the electrophoretically purified NOS/AAT. The purified protein NOS/AAT (0.1 mg) was incubated with different concentrations of cAMP for 60 min (optimal time required for the maximal binding of cAMP to NOS/AAT as shown in Figure 5). The unbound cAMP was separated from the protein bound cAMP by Millipore filtration as described in Methods and Materials.
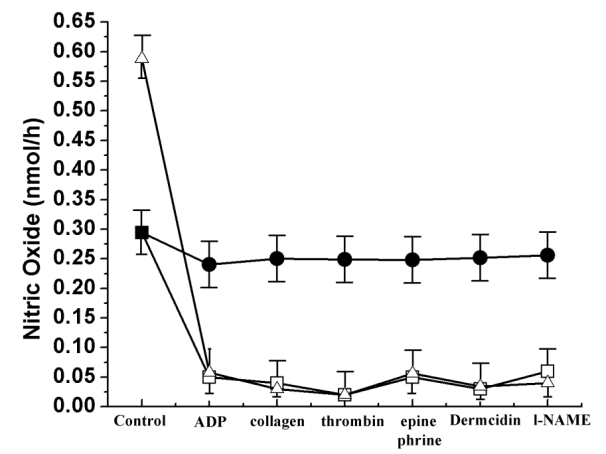
Figure 7: NO synthesis before and after removal of cAMP from the purified protein in presence of different aggregating agents. The purified protein NOS/AAT (0.1mg) was incubated with 1.2µM cAMP for 60min and the bound cAMP was removed by continued dialysis in 0.9% NaCl buffer for 24h. Subsequently NO synthesis was measured before and after the removal of the bound cAMP from the purified protein. Closed circles (•) indicate treatment of purified NOS/AAT with cAMP and aggregating agents. Open squares (□) indicate purified NOS/AAT with different aggregating agents. Open triangle (∆) indicate treatment of dialysed NOS/AAT with different aggregating agents. Each point is a mean±SD of different aggregating agents carried out atleast for 5 individual experiments. Ethics: The study used blood samples from both normal male and female volunteers. Appropriate approval was obtained from the Internal Review Board, Sinha Institute of Medical Science and Technology, Kolkata. All selected volunteers signed informed consent form before they were asked to donate blood. Funding: The study was funded by departmental resources. Disclosures: The author declares that no conflict of interests exists. Acknowledgement: The authors acknowledge the immense contribution of Late Prof. Asru Kumar Sinha throughout the study.
References
-
Janciauskiene SM, Bals R, Koczulla R, Vogelmeier C, Kohnlein T, et al. (2011) The discovery of a1- antitrypsin and its role in health and disease. Respir Med 105(8): 1129-1139.
-
Morse D, Rosas IO (2014) Tobacco smoke-induced lung fibrosis and emphysema. Annu Rev Physiol 76: 493-513.
-
Teh LG, Steven MM, Capell HA (1985) Alpha-1- antitrypsin associated liver disease in rheumatoid arthritis. Postgrad Med J 61(712): 171-172.
-
Breit SN, Wakefield D, Robinson JP, Luckhurst E, Clark P, et al. (1985) The role of alpha 1-antitrypsin deficiency in the pathogenesis of immune disorders. Clin Immunol Immunopathol 35(3): 363-380.
-
Talmud PJ, Martin S, Steiner G, Flavell DM, Whitehouse DB, et al. (2003) Diabetes atherosclerosis intervention study investigators. Progression of atherosclerosis is associated with variation in the alpha1-antitrypsin gene. Arterioscler Thromb Vasc Biol 23(4): 644-649.
-
Yang P, Sun Z, Krowka MJ, Aubry MC, Bamlet WR, et al. (2008) Alpha1-antitrypsin deficiency carriers, tobacco smoke, chronic obstructive pulmonary disease, and lung cancer risk. Arch Intern Med 168(10): 1097-1103.
-
Li Y, Krowka MJ, Qi Y, Katzmann JA, Song Y, et al. (2010) Alpha1- antitrypsin deficiency carriers, serum alpha 1-antitrypsin concentration, and non- small cell lung cancer survival. J Thorac Oncol 6(2): 291-295.
-
Topic A, Ljujic M, Nikolic A, Petrovic-Stanojevic N, Dopudja-Pantic V, et al. (2010) Alpha-1-antitrypsin phenotypes and neutrophil elastase gene promoter polymorphisms in lung cancer. Pathol Oncol Res 17(1): 75-80.
-
Lindor NM, Yang P, Evans I, Schowalter K, De Andrade M, et al. (2010) Alpha-1-antitrypsin deficiency and smoking as risk factors for mismatch repair deficient colorectal cancer: a study from the colon cancer family registry. Mol Genet Metab 99(2): 157-159.
-
Furman MI, Benoit SE, Barnard MR, Valeri CR, Borbone ML, et al. (1998) Increased platelet reactivity and circulating monocyte-platelet aggregates in patients with stable coronary artery disease. J Am Coll Cardiol 31(2): 352-358.
-
Fuster V, Badimon J, Chesebro JH, Fallon JT (1996) Plaque rupture, thrombosis, and therapeutic implications. Haemostasis 4: 269-284.
-
Trovati M, Anfossi G, Massucco P, Mattiello L, Costamagna C, et al. (1997) Insulin stimulates nitric oxide synthesis in human platelets and through nitric oxide increases platelet concentrations of both guanosine 3, 5- monophosphate and adenosine 3, 5-cyclic monophosphate. Diabetes 46(5): 742-749.
-
Whittle BJ, Moncada S, Vane JR (1978) Comparison of the effects of prostacyclin (PGI2), prostaglandin E1 and D2 on platelet aggregation in different species. Prostaglandins 16(3): 373-388.
-
Jana P, Maiti S, Ghosh R, Ghosh TK, Sinha AK (2013) Estriol, a stimulator of nitric oxide synthesis in platelets, and its role as the powerful inhibitor of platelet aggregation. Cardiovasc Endocrinol 2(3): 50-54.
-
Bhattacharyya M, Karmohapatra SK, Bhattacharya G, Bhattacharya R, Sinha AK (2009) The role of leucocytes in the acetyl salicylic acid (aspirin) induced nitric oxide synthesis in the production of interferon-alpha, a potent inhibitor of platelet aggregation and a thrombolytic agent. J Thromb Thrombolysis 28(2): 173-184.
-
Acharya K, Acharya R, Sinha AK (2001) Inhibition of low km phosphodiesterase of human blood platelets by nitric oxide. Biosci Res Bull 17: 47-53.
-
Hack N, Carey F, Crawford N (1984) The inhibition of platelet cycloxygenase by aspirin is associated with the acetylation of a 72kDa polypeptide in the intracellular membranes. Biochem J 223(1): 105- 111.
-
Kalgutkar AS, Crews BC, Rowlison SW, Garner C, Seibert K, et al. (1998) Aspirin- like molecules that covalently inactivates cycloxygenase-2. Science 280: 1268-1270.
-
Karmohapatra SK, Chakraborty K, Kahn NN, Sinha AK (2007) The role of nitric oxide in aspirin induced thrombolysis in vitro and the purification of aspirin activated nitric oxide synthase from human blood platelets. Am J Hematol 82(11): 986- 995.
-
Hamberg M, Svensson J, Wakabayashi T, Sammuelsson B (1974) Isolation and structure of two prostaglandin endoperoxides that cause platelet aggregation. Proc Natl Acad Sci USA 71(2): 345-349.
-
Banerjee D, Mazumder S, Sinha AK (2014) The role of inhibition of nitric oxide synthesis in the aggregation of platelets due to the stimulated production of thromboxane A2. Blood Coagul Fibrinol 25(6): 585-591.
-
Ghosh R, Maji UK, Bhattachraya R, Sinha AK (2012) The role of dermcidin isoform 2: A two-faceted atherosclerotic risk factor for coronary artery disease and the effect of acetyl salicylic acid on it. Thrombosis 2012: 987932.
-
Chakraborty K, Khan GA, Banerjee PA, Ray U, Sinha AK (2003) Inhibition of human blood platelet aggregation and the stimulation of nitric oxide synthesis by aspirin. Platelets 14(7-8): 421-427.
-
Jia L, Bonaventura C, Bonaventura J, Stamler SJ (1996) S-nitrosohaemoglobin: a dynamic activity of blood involved in vascular control. Nature 380(6571): 221-226.
-
Cox RD, Frank CW (1982) Determination of nitrate and nitrite in blood and urine by chemiluminiscence. J Anal Toxicol 6(3): 148-152.
-
Taniguchi T, Garcia-Higuera I, Xu B, Andreassen PR, Gregory RC, et al. (2002) Convergence of the fanconi anemia and ataxia telangiectasia signaling pathways. Cell 109(4): 459-472.
-
Chittum HS, Lane WS, Carlson BA, Roller PP, Lung FD, et al. (1998) Rabbit β-Globin Is Extended Beyond Its UGA Stop Codon by Multiple Suppressions and Translational Reading Gaps. Biochemistry 37(31): 10866-10870.
-
Eng JK, McCormick AL, Yates JR III (1994) An approach to correlate tandem mass spectral data of peptides with amino acid sequences in a protein database. J Am Soc Mass Spectrom 5(11): 976-989.
-
Kahn NN, Sinha AK (1990) Stimulation of prostaglandin E1 binding to human blood platelet membrane by insulin and the activation of adenylate cyclase. J Biol Chem 265(9): 4976-4981.
-
Bhattacharjee KG, Bhattacharyya M, Halder UC, Jana P, Sinha AK (2012) The role of Neutrophil estrogen receptor status on maspin synthesis via nitric oxide production in human breast cancer. J Breast Cancer 15(2): 181-188.
-
Salzman EW, Levin L (1971) Cyclic 3’, 5’-adenosine monophosphate in human blood platelets. II. Effect of N6-2’-O-dibutyryl cyclic 3’,5’-adenosine monophosphate on platelet function. J Clin Invest 50(1): 131-141.
-
Haslam RJ (1971) CIBA Foundation Symposia. J Clin Invest 50: 131-141.
-
Palmer RMJ, Rees DD, Ashton DS, Moncada S (1988) L-arginine is the physiological precursor for the formation of nitric oxide in endothelium-dependent relaxation. Biochem Biophy Res Commun 153(3): 1251-1256.
-
Ofran Y, Rost B (2007) Protein–Protein Interaction Hotspots Carved into Sequences. PLoS Comput Biol 3(7): 119.
-
Ghosh R, Bank S, Maji UK, Bhattacharya R, Guha S, et al. (2014) The Effect of Acetyl Salicylic Acid Induced Nitric Oxide Synthesis in the Normalization of Hypertension through the Stimulation of Renal Cortexin Synthesis and by the Inhibition of Dermcidin Isoform 2, A Hypertensive Protein Production. Int J Biomed Sci 10(3): 158-166.
-
Bhattacharya S, Chakraborty Patra S, Basu Roy S, Kahn NN, Sinha AK (2001) Purification and properties of insulin-activated nitric oxide synthase from human erythrocyte membranes. Arch Physiol Biochem 109(5): 441-449.
-
Weishaar RE, Kobylarz-Singer DC, Quade MM, Kaplan HR (1988) Role of cyclic AMP in regulating cardiac muscle contractility: Novel pharmacological approaches to modulating cyclic AMP degradation by phosphodiesterase. Drug Dev Res 12(2): 119- 129.
- How to Identify and Overcome Barriers in Developing Blood Systems?
- Why Was Transfusion Medicine Not Recognized as a Clinical Discipline?
- Outcomes of Lenalidomide Relapsed/Refractory Patients
- Is Transfusion Always Necessary?
- The Logistics of Production and Use of Blood and Blood Components
- The Challenge for Component Therapies