Modulating Lipid Composition in Cancer Cell - An Alternative Way to Augment Cancer Treatment
Many cancer drugs are mostly designed with the intent of targeting proteins deregulated in cancer. Seldom are drugs designed to target lipid composition changes in cancer. Whilst targeting proteins seems rational (because they provide diverse function to the cell and are the 'workhorse' of the cell), considering lipids in regards to cancer treatment could provide treatment alternatives. The purpose of this review is to shed light on the role of lipids in cancer and to provide insight into how lipids could be targeted to treat cancer.
Introduction
Lipids are one of the major components of the cell and if we think of cancer as of a 'cellular origin', then it is important to consider not only proteins but also lipids. The functions of lipids in the cell are numerous - Lipids form the plasma membrane of the cells which anchors many proteins (both peripheral and transmembrane proteins). The aggregation of proteins, lipids and other molecules into domains has importance in cell signaling. Lipids also influence cell shape by regulating the cells fluidity through changes of its constituent. Polyunsaturated fatty acid and cholesterol levels in cell membrane affects the cells rigidity and fluidity which also Modulating Lipid Composition in Cancer Cell - An Alternative Way to Augment Cancer Treatment could affect cell signaling, adhesion and migration [1, 2]. Lipids also play a role in cell growth and survival. The exposure of certain lipid molecules on the cell functions as a trigger of apoptosis. For instance, phosphoinositol which is mainly located in the inner leaflet of the cell membrane, when exposed on the cell becomes recognizable by immune cells which results in cell death [3]. Lastly, lipids also form a barrier which compartmentalizes cells (and organelles within it) and regulate the permeation of molecules (like drugs) in and out of a cell [4].
The above roles played by lipids (including others not listed) seem to be affected in cancer cells. Unregulated cell proliferation, drug resistance, metastasis and invasion amongst others are some of the characteristics observed in cancer cells and when these characteristics are observed closely, the roles that lipids might have played within the cells to achieve that characteristic cannot be missed. For instance, in metastasis (Figure 1), cells break Int J Lipid Res Adv
International Journal of Lipid Research & Advances
as the cell migrates to secondary site and then establishes ‘new colony’ of cells at secondary site.
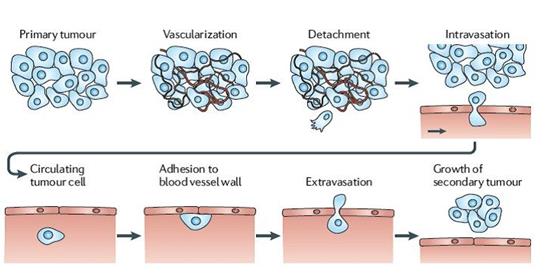
Such dynamic cell structure changes (exhibited by metastatic cells in 'changing' shapes as it moves through tight junction of blood vessel) could have been achieved not only by proteins alterations in a cell but also by lipid changes as well. Lipid changes could affect metastatic cell in ways like cell fluidity changes, curvature and deregulation in cell signaling. In fact, cell fluidity and curvature changes have been observed in some cancers and have been shown to correlate with malignancy and metastasis [5, 6]. Whilst some cancers like lymphomas and some carcinomas have been shown to exhibit high membrane fluidity, some cancers like hepatoma shows decreased membrane fluidity [7, 8, 9] (Figure 2).
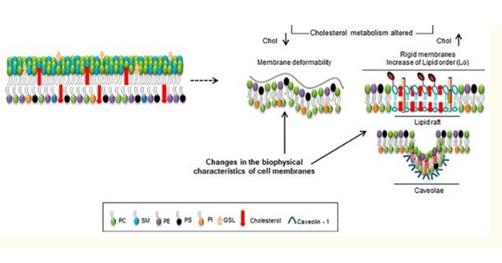
Deregulation in cell proliferation as seen in cancers would imply that, cancer cells consume high amounts of energy and increase fatty acid synthesis to achieve this process [10]. One of the main sources of energy that fuel this process is through aerobic glycolysis in what is termed as the Warburg effect [11]. During this process, cancer cells direct excess pyruvate to lactic acid production (lactic acid fermentation) regardless of the presence of oxygen. Despite cancer cells utilizing the ‘aerobic glycolysis’ pathway to meet its energy demands, fatty acid oxidation could also be seen as another pathway that cancers could use as well. Fatty acid oxidation International Journal of Lipid Research & Advances
releases more ATP per mole than the oxidation of any other nutrients. Prostate cancers and B cell lymphomas have been shown to utilize the fatty acid oxidation pathway for its energy demand even in nutrient rich environment [12].
Exploiting Lipid Changes for Cancer Research
The enormous contribution of lipids towards cell structure and function suggests that they can be exploited in a way to contribute to cancer treatment. It is important to state that tumors are composed of heterogeneous cells including cancer cells, normal cells and other stromal cells. One of the challenge to cancer therapy lies in the ability to selectively target (and kill) cancer cells whilst limiting damage to normal cells. The first step towards this approach has to do with successful identification and separation of cancer cells from normal cells and a follow up to this step will be to identify differentially expressed molecules which could be targeted in cancer.
Differentially expressed proteins (antigens) have mainly been used to characterize cancer cells. Howbeit, observed differences in lipid composition between cancer and normal cells (Figure 3) could be used as a ‘marker’ to characterize cancer cells as well.
![Figure 3: Differential distribution of lipids across membranes of Normal and Cancer Cells The outer leaflet of the plasma membrane of normal cells are predominantly composed of phosphatidylcholine (PC) and sphingomyelin (SM) (with phosphatidylserine (PS) and phosphatidylethanolamine(PE) in the inner leaflet of the plasma membrane of normal cells), Cancer cells exhibit the opposite – PE, PS on the outer leaflet of the plasma membrane and PC and SM in the inner membrane [13-15]. This ‘flip’ of leaflets as seen in cancer influences the charge distribution on cancer cell surface membrane and affects its extracellular pH (pHe) as well. It was observed by Chen B, et al. (2016) that, all twenty (20) cancer cell lines (but not normal cells) used in the experiment bound selectively to positively charged nanoprobes [16]. Manjarika De, et al. (2018) exploited this differential distribution of phosphatidylserine and developed a targeted therapy using phosphatidylcholine- stearylamine (PC-SA). They observed that, PC-SA when used either alone or in complex with doxorubicin and camptothecin had profound tumour killing with no signs of toxicity [17].](/fulltextimages/3951/fig_3.jpeg)
Figure 3: Differential distribution of lipids across membranes of Normal and Cancer Cells The outer leaflet of the plasma membrane of normal cells are predominantly composed of phosphatidylcholine (PC) and sphingomyelin (SM) (with phosphatidylserine (PS) and phosphatidylethanolamine(PE) in the inner leaflet of the plasma membrane of normal cells), Cancer cells exhibit the opposite – PE, PS on the outer leaflet of the plasma membrane and PC and SM in the inner membrane [13, 14, 15]. This ‘flip’ of leaflets as seen in cancer influences the charge distribution on cancer cell surface membrane and affects its extracellular pH (pHe) as well. It was observed by Chen B, et al. (2016) that, all twenty (20) cancer cell lines (but not normal cells) used in the experiment bound selectively to positively charged nanoprobes [16]. Manjarika De, et al. (2018) exploited this differential distribution of phosphatidylserine and developed a targeted therapy using phosphatidylcholine- stearylamine (PC-SA). They observed that, PC-SA when used either alone or in complex with doxorubicin and camptothecin had profound tumour killing with no signs of toxicity [17].
Lipid displacement represents another method in which lipid alteration could contribute to cancer treatment2. Research has shown that, consumption of polyunsaturated fatty acids and mono (oleic acid) contributes to preventing colon cancer. The mechanism by which these fatty acids contribute to colon cancer prevention is through changing the lipid composition of the plasma membrane and affecting the cells fluidity as the fatty acids incorporates into the cell membrane. Synthetic analog of oleic acid, 2 hydroxyoleic acid and also propofol-DHA have shown to exhibit antitumour activity [18, 19].
Again, lowering membranous cholesterol levels have been shown to be relevant in cancer treatment. It was shown that Emodin (the biologically active components found in roots and rhizome of Rheum palmatum) inhibits tumour cell migration, invasion and adhesion by lowering cholesterol levels and disrupting membrane lipid raft- associated integrin signaling [20, 21].
Conclusion
Lipids are important class of molecules pertinent to maintaining cell structure and function. The general cell characteristics displayed by cancer cells seems to reflect what could 'go wrong' in lipid alteration in cells. It is therefore important that lipids studies be given much attention to explore avenues of drugs that could target them in cancer.
References
-
Sunshine H, Iruela-Arispe ML (2017) Membrane Lipids and Cell Signaling. Curr Opin Lipidol 28(5): 408-413. International Journal of Lipid Research & Advances
-
Bernades N, Fialho AM (2018) Perturbing the Dynamics and Organization of Cell Membrane Components: A New Paradigm for Cancer-Targeted Therapies. Int J Mol Sci 19(12): 3871.
-
Schlegel RA, Williamson P (2001) Phosphatidylserine, a death knell. Cell Death Differ 8(6): 551-563
-
Gerrit VM, Dennis RV, Gerald WF (2008) Membrane lipids: Where they are and how they behave. Nat Rev Mol Cell Biol 9(2): 112-124.
-
Shinitzky M (1984) Membrane fluidity in malignancy adversative and recuperative. Biochim Biophys Acta Rev Cancer 738(4): 251-261.
-
Nakazawa I, Iwaizumi M (1989) A role of the cancer cell membrane fluidity in the cancer metastases: an ESR study. Tohoku J Exp Med 157(3): 193-198.
-
Galeotti T, Borrello S, Minotti G, Masotti L (1986) Membrane alterations in cancer cells: the role of oxy radicals. Ann N Y Acad Sci 488: 468-448.
-
Sherbet GV (1989) Membrane fluidity and cancer metastasis. Exp Cell Biol 57(4): 198-205.
-
Sok M, Sentjurc M, Schara M, Stare J, Rott T (2002) Cell membrane fluidity and prognosis of lung cancer. Ann Thorac Surg 73(5): 1567-1571.
-
Fadaka A, Ajiboye B, Ojo O, Adewale O, Olayide I, et al. (2017) Biology of glucose metabolization in cancer cells. Journal of Oncological Sciences 3(2): 45-51.
-
Melone MAB, Valentino A, Margarucci S, Galderisi U, Giordano A, et al. (2018) The carnitine system and cancer metabolic plasticity. Cell Death Dis 9(2): 228.
-
Wu X, Daniels G, Lee P, Monaco ME (2014) Lipid metabolism in prostate cancer. Am J Clin Exp Urol 2(2): 111-120.
-
Caro P, Kishan AU, Norberg E, Stanley IA, Chapuy B, et al. (2012) Metabolic signatures uncover distinct targets in molecular subsets of diffuse large B cell lymphoma. Cancer Cell 22(4): 547-560.
-
Ran S, Thorpe PE (2002) Phosphatidylserine is a marker of tumor vasculature and a potential target for cancer imaging and therapy. Int J Radiat Oncol Biol Phys 54(5): 1479-1484.
-
Ran S, Downes A, Thorpe PE (2002) Increased exposure of anionic phospholipids on the surface of tumor blood vessels. Cancer Res 62(21): 6132-6140.
-
Chen B, Le W, Wang Y, Li Z, Wang D, et al. (2016) Targeting Negative Surface Charges of Cancer Cells by Multifunctional Nanoprobes. Theranostics 6(11): 1887-1898.
-
Manjarika De, Ghosh S, Sen T, Shadab M, Banerjee I, et al. (2018) A Novel Therapeutic Strategy for Cancer Using Phosphatidylserine Targeting Stearylamine- Bearing Cationic Liposomes. Mol Ther Nucleic Acids 10: 9-27.
-
Lladó V, López DJ, Ibarguren M, Alonso M, Soriano JB, et al. (2014) Regulation of the cancer cell membrane lipid composition by NaCHOleate: effects on cell signaling and therapeutical relevance in glioma. Biochim Biophys Acta 1838(6): 1619-1627.
-
Escribá PV, Busquets X, Inokuchi J, Balogh G, Török Z, et al. (2015) Membrane lipid therapy: Modulation of the cell membrane composition and structure as a molecular base for drug discovery and new disease treatment. Prog Lipid Res 59: 38-53.
-
Huang Q, Shen HM, Shui G, Wenk MR, Ong CN (2006) Emodin inhibits tumor cell adhesion through disruption of the membrane lipid Raft-associated integrin signaling pathway. Cancer Res 66(11): 5807- 5815.
-
Ok S, Kim SM, Kim C, Nam D, Shim BS, et al. (2012) Emodin inhibits invasion and migration of prostate and lung cancer cells by down regulating the expression of chemokine receptor CXCR4. Immunopharmacol Immunotoxicol 34(5): 768-778.