The Role of a Rich Landscape Vegetation on kob Antelope (Kobus kob) Group Feeding Activity in Bouba Ndjidda National Park, North Region, Cameroon
Thorough knowledge of the ecological requirements of ungulates provides the basis for the formulation of an appropriate programme of management actions directed at that species. More so, the identification of the minimum habitat requirements for a species is essential before specific management actions can be instituted. However, the relationship between antelopes and habitat plays a crucial role in the ecology of any species. The association between a herbivore and its habitat not only encompasses the availability of preferred food plants and their particular growth stages in its home range, it is also intimately linked to other features of the habitat. Hence, the key objective of this study was to assess the role of landscape vegetation on antelopes’ sustainability. Data collection was initiated immediately after a thorough study was done. The kob antelopes groups were visited regularly to record both feeding activity and ecological factors. The study showed a significance between kob antelope group and vegetation, X2 = 7.383 df=8, P<0.05. The antelope-group strength was dominated by 1-5 antelopes 89%, but larger group-sizes were equally recorded though not common. The three vegetation types most used by the antelopes were grassland 49%, shrub-land 36%, and forest patches 11%. Nonetheless, the antelope-group activity showed a significance on vegetation type X2 = 11.510 df=4, P<0.05. The most significant animal activities recorded were movement 49%, feeding 35%, and resting 16% respectively. Similarly, antelope-group activity revealed a significance on photo-period X2 = 5.348 df=2, P<0.05. Additionally, the antelope-group size showed a significance on landscape X2 = 7.364 df=8, P<0.05. Bouba ndjidda national park in the northern region of Cameroon has three landscape types useful to the kob antelopes, flat 73%, slope 25%, and hill 2% respectively. More so, there was a positive correlation significance between vegetation and food resources r = 0.248, P<0.05. The landscape of the national park has very little escapement that could obstruct movement of these medium- sized antelopes. Furthermore, their vigilance, speed against predators, and prolific nature might help the antelope achieve a high population in the national park in future. Most of these antelopes have received very little research attention, however, more study is needed to explore the social organization, feeding and population ecology.
Introduction
The acquisition of resources for survival, growth and reproduction of foraging animals depends on the habitat used, on the set of physical environmental conditions and resources surrounding the animal Garshelis [1]; Morrison [2] and on the breadth of habitat conditions used [3]. In addition, habitat breadth provides insights into the animal’s potential distribution range [4], such that species with broad habitat tolerance are potentially more widely distributed than species using a narrower diversity of habitats. The use of a narrow range of habitat features reflects specialization or marginality, which is one measure of niche breadth [5]. Habitat specialization may limit the potential of species to use spatially and temporally variable resources, conditions and security. Brown [6] and Rosenzweig and Lomolino [7] argued that habitat specialization leads to both local and regional rarity of species. However, Seagle and McCraken [8] and Gaston and Kunin [9] demonstrated that specialization on abundant resources or widely distributed habitats can result in high abundance of a species. du Toit and Owen- Smith [10] observed differences in habitat use and in habitat breadth among African browsers, with smaller species such as steenbok (Raphicerus campestris, 11 kg) using a narrower range of habitats than larger browsers such as kudu (Tragelaphus strepsiceros, 180 kg) and giraffe (Giraffa camelopardalis, 800 kg), which exploited a wider range of habitats, including habitats patches neglected by the smaller browser. Following Bell [11] principle, du Toit and Owen- Smith [10] proposed that the more even use of a broader range of habitats by kudu and giraffe was because larger animals tolerate a wide range of food items, including low quality and food items of large size neglected by the smaller animal.
Habitat use is a hierarchical process [12], scaling spatially from the selection of geographical range (first order habitat selection), selection of broad vegetation types across the landscape when establishing home ranges (second order habitat selection), selection of habitats within the home range (third order selection), selection of feeding areas within habitats to selection of food items within feeding areas (fourth order selection) [13]. Following the limiting similarity theory for species coexistence Rosenzweig [14] stated that species using the same landscape differ in the use of one or more habitat features, because differential habitat use reduces the potential for competition. Therefore, similarities or differences in habitat selection among co- occurring ungulates, hence potential for competition or for stable coexistence through habitat partitioning should take place at those hierarchical levels of habitat selection. The relationships and interactions between an animal and its environment play a fundamental role in the ecology of any species. To conserve an animal one must take cognizance of the need to identify its habitat requirements and preserve the various elements which constitute the habitat Riney [15] defined habitat requirements as the minimum requirements for the survival of an animal. Identification of limiting habitat elements allows for the manipulation of these to enhance the survival of threatened or endangered species Habitat studies generally involve relating the distribution of an animal to those habitat resources that are considered to be critical for its survival, for example, food, water and shelter. Various approaches have been adopted for the purpose of studying habitat use. These include descriptive surveys of the historical distribution of a particular species, qualitative assessments of habitat preferences and analytical approaches which involve detailed statistical evaluation of the relationship between site occupancy and various habitat elements [16].
Ben-Shahar [17] was unable to show that vegetation structure had any effect on the distribution pattern of roan in the Northwest Transvaal. Roan in his study area, however, showed an affinity for moderate slopes. Favored roan habitat in the national park was on slightly undulating flats consisting of heavy clay soils derived from basalt although hillier areas on clay-loam soils derived from dolerite were also utilized, but to a lesser extent [18]. Ruminants require higher quality forage than non-ruminants of similar body mass [19]. Smaller herbivores require higher quality forage than larger herbivores [20]. Sharing space with conspecifics results in rapid attrition of food resources within habitat patches [20], but the high mobility of animals in large herds allows animals to use spatially variable resources, whereas sedentary species use resources occurring in small areas [21]. Animals occurring in smaller groups are able to use high quality food resources occurring in small habitat patches not detected or neglected by larger herds [22]. Therefore, following allometric relations of body mass and group size as well as gut morphology and function, smaller ruminants occurring in smaller herds should select for the combination of habitat features that ensure the acquisition of the required higher quality food resources, whereas larger ruminants occurring in larger herds and hindgut fermenters should use habitat feature more evenly.
Materials and Methods
The Description of Study Area
Bouba ndjidda national park is located in the northern region of Cameroon near the Chadian border, between latitude 8°37′ and 8°37′ N and longitude 14°39′ and 14°39′ E. It was created in 1932 as a wildlife reserve and became a national park in 1968, and covers an area of 220,000 ha [23]. The national park has the Sudano-guinean climate characterized by two seasons, a six-month rainy season from late April to mid-October and a dry season from November to April during the period which no rain is observed [23]. The region receives between 1000 and 1,250 mm of precipitation per year and the rainiest months are August and September while the annual average temperature is 28°C. The region has a dense hydrographic network, unlike other rivers that are seasonal in this part of the country. Water is abundant throughout the national park and even during the dry season, thus ensuring the development of ecological interaction processes between wildlife species and habitat characteristics in the environment. The national park is also a home to 24 species of large and medium mammals such as African elephant, Lions, Giraffes, Leopards, Hyenas, Bush pig, Warthogs, Hippos, Buffaloes, Hartebeest antelopes, Eland antelopes, Kob antelopes, Reedbuck antelopes, etc. Additionally, the national park homes about 250 bird species. Considering the ornithological richness of the national park, it has been declared and area of importance for the conservation of birds Figure 1.
![Figure 1: Map of Bouba ndjidda National Park [23].](/fulltextimages/9554/fig_1.png)
Data Collection
Data on antelopes activity in the preferred habit were collected by direct observation (focal sampling method) from appropriate visible points. This includes watching an individual or group of antelopes for 10 minutes within an interval of 5 minutes. Antelopes were observed in the early morning (06:30 to 10:30) and in the late afternoon (14:00 to 18:00) when they are usually active [24, 25]. Recordings were ceased when the animal moved out of the sight. Data were collected more than ten times per month on antelopes’ habitat activity. More so, observations were recorded when the antelopes were foraging (grazing/browsing/chewing/ biting) plants (tree/shrub/herb/grass) or consumed (Leaf/ shoot/stem/fruit) [25]. Each record was counted when individual hartebeest were observed feeding continuously from a single plant species or plant group [26]. More so, Herd size and composition were recorded whenever the antelope herd was encountered during the study period. The activity that the majority of the herd members were engaged in, on each observation, specifically grazing, resting and other activities (feeding, drinking, walking and other social interactions) was noted along with the time.
Data Analysis
Before conducting the actual data analysis, data were checked for some errors (e.g. recording and consistencies). Data were analyzed using statistical package for social science (SPSS) version 20 software (α = 95% level of significance, p < 0.05), descriptive statistics and Excel spreadsheet. The variables were cross-tabulated to understand their degree of association.
Results
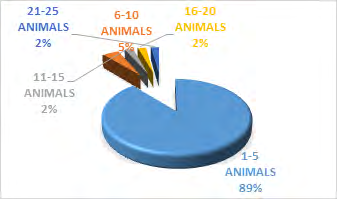
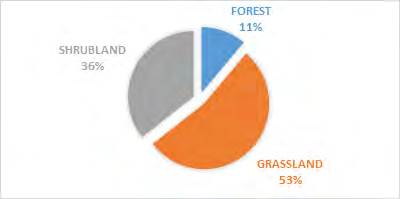
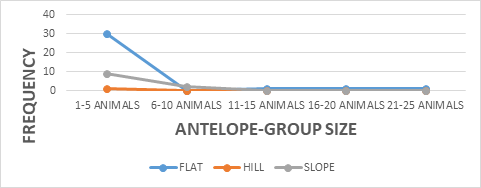
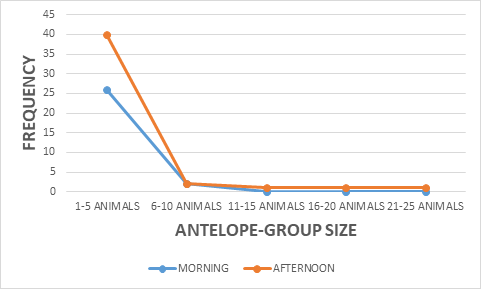
The study showed a significance between kob antelope group and vegetation, X2 = 7.383 df=8, P<0.05 (Figure 2). The kob antelope population in Bouba ndjidda is seemly high, for this reason they form larger group congregations compared to other antelope species in the national park. The animal- group strength was dominated by 1-5 animals 89%, but larger group sizes were equally recorded though not common (Figure 3). Generally, kob antelopes are characterized by large groups in most national parks in Cameroon. Their population in the savanna national parks has placed this animal species non-threatened on conservation management strategy. The floodplain has been the most favored habitat for the kob antelopes in the national park. Bouba ndjidda is rich in drainage system compared to other national parks in the savanna ecosystem of Cameroon. The three vegetation types used most by the antelopes were grassland 49%, shrub-land 36%, and forest patches 11% (Figure 4). The study observed the dwelling of kob antelopes on grassland more than any other habitat because they are grazing herbivores.
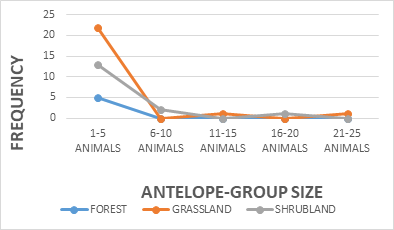
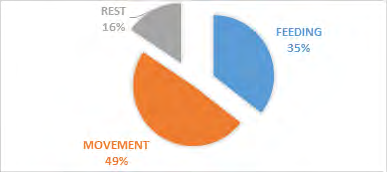
Nonetheless, the antelope-group activity showed a significance on vegetation type X2 = 11.510 df=4, P<0.05 (Figure 5). The most significant animal activities recorded were movement 49%, feeding 35%, and resting 16% respectively (Figure 6). The movement activity dominance in antelopes could be caused by food scarcity in most savanna national parks. The reason was that wildlife such as antelopes seems to move more in search of food resources during the dry season period when grasses are dried up in many areas. The fact that the survey was done during the dry season period could contribute to the high animal movement recorded. Most of the studies carried out on wildlife in the savanna ecosystem areas in sub Saharan Africa has favored animal movement activity during the dry season compared to the rainy season. Similarly, antelope-group activity revealed a significance on photo-period X2 = 5.348 df=2, P<0.05 (Figure 7). Both the morning and afternoon periods witnessed an increase in antelope-group activity, feeding, movement and rest.
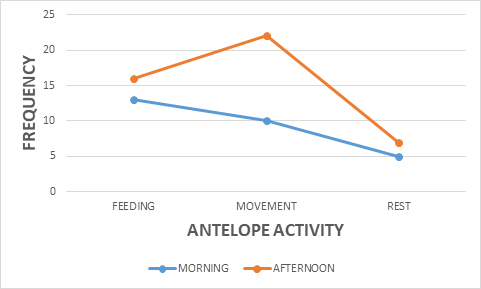
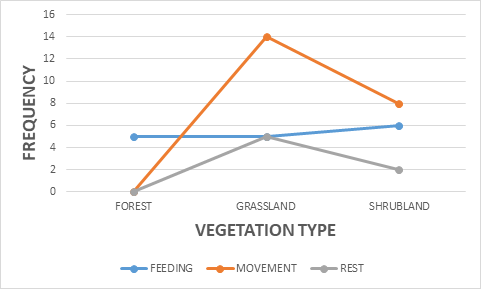
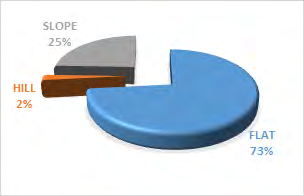
Additionally, the antelope-group size showed a significance on landscape X2 = 7.364 df=8, P<0.05 (Figure 8). Bouba ndjidda national park in the northern region of Cameroon has three landscape types useful to the kob antelopes, flat 73%, slope 25%, and hill 2% respectively (Figure 9). Most of the kob antelope feeding is carried out in the flat areas dominated by grassland vegetation. The flat landscape is rich in grass vegetation, especially in the flood plains, might have contributed to the high population of this antelope species in the national park. Kob antelopes have a smaller home range and territory compared to other antelope species which help them to cope with overlapping home ranges with other wildlife species.
Furthermore, the antelope-group size recorded a positive correlation significance on photo-period r = 0.280, P<0.05 (Figure 10). Wildlife-species grouping has social benefits such as inter species defense, location of food resources in other areas of the national park, and control of sub adults and juveniles. This social organization also helps to keep the antelopes closed to each other for life sustenance.
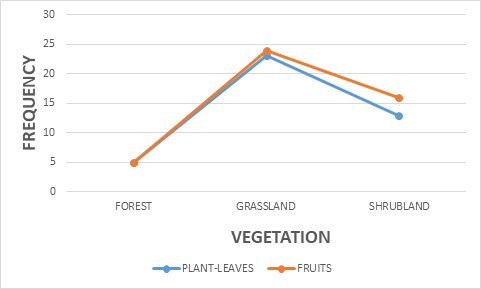
More so, there was a positive correlation significance between vegetation and food resources r = 0.248, P<0.05 (Figure 11). The three vegetation types have been observed to contribute towards the feeding welfare of the kob antelopes in the national park. Bouba ndjidda national park is in the woodland savanna ecosystem region of north Cameroon, poor in forest vegetation compared to grassland and shrub- land. This antelope species makes more use of the grassland vegetation because of its herbivorous grazing characteristics.
Discussion
The relationship between animal and habitat plays a crucial role in the ecology of any species. The association between a herbivore and its habitat not only encompasses the availability of preferred food plants and their particular growth stages in its home range, it is also intimately linked to other features of the habitat [27]. These include inter alia, (i) the availability of suitable cover, (ii) reproduction, (iii) the absence of adequate nutrition excessive intra for and interspecific competition, (iv) the frequency of fires, (v) the influence of predation and parasitism and (vi) the extent to which the habitat provides minimum resources to sustain the species. Effective management of multi-species ungulate populations requires knowledge of the habitat requirements of the individual species and should take cognizance of the existence of interspecific competition. The extent of resources available must also be quantified to prevent environmental degradation. In the last three decades, numerous studies have been conducted of mammalian herbivores in relation to their environment. These range from qualitative observations of the types of habitats used by particular ungulate species or assemblages to more complex quantitative investigations that attempt to explain why such habitats are used [28, 29]. Studies aimed at establishing the habitat preferences and other ecological requirements of herbivorous animals are particularly important for the conservation of vulnerable species [17].
Thorough knowledge of the ecological requirements of an endangered ungulate provides the basis for the formulation of an appropriate programmer of management actions directed at that species. The identification of the minimum habitat requirements for a species is essential before specific management actions can be instituted. When sufficient knowledge. is available, measures that can be implemented include, (i) manipulation of the habitat to make it as productive for the species as possible, (ii) the installation of dry season water points, (iii) the reduction of competitors for food and (iv) the control of predators [15]. In the absence of sufficient grazing pressure and as a result of high rainfall, certain grass species tend to alter the structure of the grass layer resulting in a loss of palatability particularly for species that are sensitive to the amount of stem in the grass sward [30]. Burning is used as a method to defoliate grassland periodically and maintain it in an optimum condition for antelopes.
Habitat use by large herbivores is partly influenced by the spatial and temporal variation in the quantity and quality of forage and by water availability [31, 32]. However, apart from differences in food preference among grazers and differences in the availability of preferred food among habitats, differences in predation risk among habitats as well as differences in anti-predator behavior Sinclair and Arcese [33]; Bowyer [31] and social system Duncan [34] might contribute in explaining differential habitat use among ungulate species. Aggregation also reduces the risk of predation through simple dilution or by increased probability of detecting predators [35]. Accordingly, species that occur in large herds and that are able to fight against predators through group attack such as buffalo Prins and Iason [36]; Prins [37] may use a wide range of habitats, including habitats with relatively higher predation risk such as thick bush [38]. Ungulates that run away to escape predators such as plains zebra (Equus burchelli) Estes [39] should select for open habitats with good visibility for rapid detection of predators and should avoid thick bush providing cover for stalking predators [40], whereas species such as sable, not documented to successfully fight against large predators, should avoid being detected by predators by using habitats providing adequate woody cover for concealment.
Differences between grass species in greenness retention, leaf production and quality lead grazing animals to prefer some grass species and neglect others [41]. Selective feeding results in a progressive reduction in the standing biomass of preferred grass species through the dry season until the first wet season rains promote the regrowth of these depleted grass species [40]. Foraging theory predicts that foraging animals widen their diets to incorporate less favored food types, as the availability of preferred food resources declines towards the end of the dry season [42]. The widening of the diet by grazers could be through widening the range of grass species grazed [41, 43], but also through increasing tolerance to brown grass, stems or less favored grass height as the dry season progresses. Owen-Smith [40] argued that hungry herbivores face the dilemma of how to be narrowly selective or broadly tolerant when food resources are abundant but most of which is of low nutritional quality such as during the late dry season.
Grassland height, catena position, woody canopy cover, grassland species composition Ben-Shahar [17]; Owen-Smith [40] and grass greenness [44] influence forage resources and habitat conditions for large grazers. Therefore, distinct habitat preference Johnson [40]; Owen-Smith [45] among grazers could be through differences among species in preference for these habitat features. Green grass leaves have higher concentration of protein, minerals and soluble carbohydrates than brown leaves [3]. Accordingly, during the dry season grazers should select foraging areas where the highly nutritious green grass prevails [46]. Food intake rate increases with grass height because with increasing grass height, bite size increases as animals apprehend a bigger volume of herbage through increased bite depth [46, 47]. Thus, during the dry season when forage quantity declines due to the absence of regrowth, large grazers should concentrate foraging on patches of tall grass offering high intake rates [48, 49].
Conclusion
Kob antelopes are grazers, a characteristic that fits their existence and population increase in this national park. Additionally, their group aggregation behavior in social organization order has been one of the major survival strategies in woodland savanna ecosystems like bouba ndjidda national park. The fact that this national park in particular is far from cities, wildlife faces insignificant poaching effect. More so, the surveillance done by the patrol teams made up of the military and eco-guards has further strengthen the security of wildlife in the national park. However, the grass-vegetation dominance has contributed to the increase of herbivorous population. Even in the heart of dry season so many areas in this national park still maintain grassland vegetation. The landscape of the national park has very little escapement that could obstruct movement of these medium-sized antelopes. Furthermore, their vigilance, speed against predators, and prolific nature might help the antelope achieve a high population in the national park in future. Most of these antelopes have received very little research attention; however, more study is needed to explore the social organization, feeding and population ecology.
References
-
Garshelis DL (2000) Delusions in Habitat Evaluation: Measuring Use, Selection, and Importance, In: Research Techniques in Animal Ecology (eds: L. Boitani and T.K. Fuller). Columbia University Press, pp: 111-164.
-
Morrison ML (2001) A proposed research emphasis to overcome the limits of wildlife-habitat relationship studies. Journal of Wildlife Management 65(4): 613-623.
-
Owen-Smith N (1982) Factors influencing the consumption of plant products by large herbivores. Ecology of Tropical Savannas, In: Huntley BJ and Walker BH (Eds.), Springer-Verlag, Berlin pp: 359-404.
-
Morrison ML, Marcot BG, Mannan RW (2006) Wildlife- Habitat Relationships - Concepts and Applications. Island Press, Washington.
-
Calenge C, Dufour AB, Maillard D (2005) K-select analysis: a new method to analyse habitat selection in radio-tracking studies. Ecological Modelling 186: 143- 153.
-
Brown JH (1984) On the relationship between abundance and distribution of species. The American Naturalist 124(2): 255-277.
-
Rosenzweig, ML, Lomolino MV (1997) Who gets the short bits of the broken stick? In: Kunin, W.E. & Gaston (eds) The Biology of Rarity: causes and consequences of rare-common differences. Chapman & Hall, London pp: 63-90.
-
Seagle SW, McCraken GF (1986) Species abundance, niche position and niche breadth for five terrestrial animal assemblages. Ecology 67(3): 816-818.
-
Gaston KJ, Kunin WE (1997) Rare-common differences: an overview In: The Biology of Rarity: causes and consequences of rare-common differences, In: Kunin WE and Gaston (Eds.), Chapman & Hall, London pp: 12-30.
-
Du Toit JT, Owen-Smith N (1989) Body size, population metabolism and habitat specialization among large African herbivores. American Naturalist 133(5): 736- 740.
-
Bell RH (1970) The use of the herb layer by grazing ungulates in the Serengeti. Animal Populations in Relation to Their Food Resources In: Watson A (Eds.), Blackwell Scientific Publications, Oxford pp: 112-124.
-
Senft RL, Coughnour MB, Bailey DW, Rittenhouse LR, Sala OE, et al. (1987) Large herbivore foraging and ecological hierarchies-landscape ecology can enhance traditional foraging theory. Bio Science 37(11): 789-799.
-
Johnson DH (1980) The comparison of usage and availability measurements for evaluating resource preference. Ecology 61(1): 65-71.
-
Rosenzweig ML (1981) A theory of habitat selection. Ecology 62(2): 327-335.
-
Riney T (1982) Study and management of large mammals. John Wiley and Sons.
-
Beardall GM, Joubert SCJ, Retief PF (1984) An evaluation of the use of correspondence analysis for the analysis for the analysis of herbivore-habitat selection. S Afr J Wildl Res 14: 79-88.
-
Ben-Shahar R (1995) Habitat classification in relation to movements and densities of ungulates in a semi-arid savanna. African Journal of Ecology 33(1): 50-63.
-
Joubert SCJ (1976) The population ecology of the roan Hippotragus equinus (Desmarest 1804) in the Kruger Park. DSC. (Wildlife Management) dissertation, University of Pretori.
-
Illius AW, Gordon IJ (1992) Modelling the nutritional ecology of ungulate herbivores: evolution of body size and competitive interactions. Oecologia 89: 428-434.
-
Owen Smith N (1988) Megaherbivores-The influence of a very large body size on ecology. Cambridge University Press. Cambridge.
-
Ritchie M (2002) Competition and Coexistence of Mobile Animals. Competition and Coexistence, In: Sommer V and Worm B (Eds.), Springer-Verlag, Berlin pp: 109-131.
-
Fryxell JM (1991) Forage quality and aggregation by large herbivores. American Naturalist 138(2): 478-498.
-
Klop E, van Goethem J (2008) Savanna fires govern community structure of ungulates in Bénoué National Park, Cameroon. J Trop Ecol 24(1): 39-47.
-
Tahani AH, Ibrahim MH (2014) Diurnal Activity of Waterbuck (_Kobus ellipsiprymnus defassa_) in Dinder National Park, Sudan. Journal of Natural Resource & Environmental Study 2(2): 15-19.
-
Wubie Bayie and Mesele Yihune (2018) Population status, feeding ecology and activity pattern of common bushbuck (_Tragelaphus Scriptus decula_) in Sekele Mariam Forest, West Gojjam, Ethiopia. Journal of Ecology and the Natural Environment 10(5): 69-79.
-
Brnesh H, Tsegaye G, Tadese H, Gelaye GM (2015) The Status and Feeding Ecology of Menelik’s Bushbuck (_Tragelaphus scriptus meneliki)_ in Wof-Washa Forest, North Shoa, Ethiopia. International Journal of Current Research 75: 16366-16370.
-
Pienaar UDEV (1974) Habitat preference in South African antelope species and its significance in natural and artificial distribution patterns. Koedoe 17(1): 185- 189.
-
Williamson DT (1990) An outline of the ecology and behaviour of red lechwe (Kobus leche leche Gray 1850). Afr J Ecol 28: 89-101.
-
Funston PJ, Skinner JD, Dott HM (1994) Seasonal variation in movement patterns, home range and habitat selection of buffaloes in a semi-arid habitat. African Journal of Ecology 32(2): 100-114.
-
Heitkonig IMA (1993) Feeding strategy of roan antelope (_Hippotragus equinus_) in a low nutrient savanna. PhD Thesis. University of Witswatersrand.
-
Bowyer RT, Kie JG, van Ballenberghe V (1998) Habitat selection by neonatal black-tailed deer: climate, forage, or risk of predation. Journal of Mammalogy 79(2): 415- 425.
-
Winnie J, Cross P, Getz W (2008) Habitat quality and heterogeneity influence distribution and behavior in African buffalo (_Syncerus caffer_). Ecology 89(5): 1457- 1468.
-
Sinclair ARE, Arcese P (1995) Population consequences of predation-sensitive foraging: the Serengeti wildebeest. Ecology 76(3): 882-891.
-
Duncan P (1983) Determinants of the use of habitat by horses in a Mediterranean wetland. Journal of Animal Ecology 52(1): 93-109.
-
Fryxell JM (1995) Aggregation and Migration by Grazing Ungulates in Relation to Resources and Predators. Serengeti II – Dynamics, Management and Conservation of an Ecosystem, In: Sinclair ARE and Arcese P (Eds.), The University of Chicago Press, Chicago, pp: 257-273.
-
Prins HHT, Iason GR (1989) Dangerous lions and nonchalant buffalo. Behaviour 108(4): 262-296.
-
Prins HHT (1996) Ecology and Behaviour of the African Buffalo-Social Inequity and Decision Making. Chapman and Hall, London.
-
van Orsdol KG (1984) Foraging behavior and hunting success of lions in Queen Elizabeth National Park, Uganda. African Journal of Ecology 22(2): 79-99.
-
Estes RD (1991) Behavior Guide to African Mammals: including hoofedmammals, carnivores, and primates. The University of California Press, California.
-
Owen-Smith N (2002) Adaptive Herbivore Ecology-from resources to populations in variable environments. Cambridge University Press. Cambridge.
-
O’Reagain P, Mentis MT (1989) Sequence and process of species selection by cattle in relation to optimal foraging theory on an old land in the Natal sour sandveld. Journal of the Grassland Society of Southern Africa 6(2): 71-76.
-
Stephens DW, Krebs JR (1986) Foraging theory. Princeton University Press, Princeton.
-
Macandza VA, Owen-Smith N, Cross PC (2004) Forage selection by African buffalo in the late dry season in two landscapes. South African Journal of Wildlife Research 34: 113-121.
-
Sinclair ARE (1985) Does interspecific competition or predation shape the African ungulate community? Journal of Animal Ecology 54(3): 899-918.
-
Johnson BK, Kern JW, Wisdom MJ, Findholt SL, Kie JG (2000) Resource selection and spatial separation of mule deer and elk during spring. Journal of Wildlife Management 64(3): 685-697.
-
O’Reagain P (2001) Foraging strategies on rangeland: effects on intake and animal performance. Proceedings of the XIX International Grassland Congress, Sao Paolo pp: 277-284.
-
Laca EA, Ungar ED, Seligman N, Demment MW (1992) Effects of sward height and bulk density on bite dimensions of cattle grazing homogeneous swards. Grass Forage Sci 47(1): 91-102.
-
Wilmshurst JF, Fryxell JM, Farm BP, Sinclair ARE, Henschel CP (1999) Spatial distribution of Serengeti wildebeest in relation to resources. Canadian Journal of Zoology 77(8): 1223-1232.
-
Ben sharar R (1990) Resource availability and habitat preferences of three African ungulates. Bio I Conserv 54(4): 357-365.
- Lessons to Learn: Trees are More than the Lungs of the World
- Community Forestry Enterprises as a Model for Sustainable Forest Development: The Case Of The "Baja Tarahumara" in Chihuahua, Mexico
- Ecological and Socio-Economic Impacts of Chromolaena odorata and Mesosphaerum suaveolens, Two Invasive Alien Species in Central and Southern Benin, West Africa
- Epigenetic Sustainability: Modeling the Human Factor as a Natural Resource through Science 4.0 and the NR3C1 Biological Pilot
- Growth-at-Risk: A Framework for Assessing Economic Vulnerability
- The Rural Territory as a Socioecological System for the Management of Public Policy for Sustainable Rural Development