Ecological and Evolutionary Consequences of Gene Flow through Pollen from Transgenic Crops to Their Wild Relatives
The rapid progress in biotechnology has significantly promoted the development and production of genetically modified (GM) crops. The extensive global cultivation of GM crops has generated great benefits, which may provide opportunities for solving the problems inherent in world food security, but it has also aroused considerable biosafety concerns worldwide. Among these, the potential environmental consequences created by possible transgene escape from a GM crop to its non- GM crop counterparts and wild or weedy relatives via gene flow are the most debated biosafety issues internationally. Gene flow indicates the movement of genes or genetic materials from one plant population into another. There are a range of predicted possible environmental and evolutionary consequences such as the creation of new weeds, change of fitness-related characters, and loss of genetic diversity in crop landraces and wild relatives) caused by wild transgene flow. In addition, transgene flow also arouses biosafety concerns for food/feed and health and socio-economics and ethics. Through pollen- mediated gene flow, a transgene can move from a GM crop into populations of a wild relative, and persist or disseminate in the wild population through further hybridization and introgression between the GM crop and wild relative. If a transgene can express in wild plants as it does in the GM crop, the transgene may change a certain trait of the wild plants, possibly leading to further undesired consequences. If a transgene can alter the fitness of wild plants and the dynamics of the wild populations, the introgression of the transgene in the wild population may cause either local extinction of the population or make the wild population more invasive and competitive. Therefore, a wide variety of possible ecological and evolutionary consequences of gene flow through Pollen from transgenic plants to their wild relatives are discussed on the basis of previous work.
Introduction
Gene flow is the movement of genetic materials (genes or alleles) from one organism to another. Theoretically, there are two types of gene flow: vertical gene flow and horizontal gene flow. Horizontal gene flow occurs only among unrelated species, such as between plants and microbes, as well as between microorganisms Thomson JA [1] whereas Vertical gene flow is the movement or transfer of genes or alleles by normal reproductive processes, between separate populations of plant and animal species. Gene flow (vertical) is a term extensively used in evolutionary biology and population genetics, long before the issue of transgene escape from GM crops was raised. In population genetics, gene flow (also known as gene migration) refers to the transfer of alleles or genes from one population to another Hartl DL [2] thereby changing the composition of the gene pool of the receiving population. If the rate of gene flow is high enough, then two populations will have equivalent allele frequencies and therefore can be considered a sine effective population. It has been shown that it takes only “one migrant per generation” to prevent populations from diverging due to drift. The introduction of new alleles through gene flow increases variability within the population and makes possible new combinations of traits. Although gene flow does not change allele frequencies for a species as a whole, it can alter allele frequencies in local populations. In the case of migration, the greater the difference in allele frequencies between the resident and the migrant individuals, and the larger the number of migrants, the greater the effect the migrants have in changing the genetic constitution of the resident population [3].
The rapid development of biological science and technology has brought the world into a new era of biotechnology [4, 5]. One of the most important characteristics of the biotechnology era is the wide application of genetic engineering (also referred to as genetic modification) technologies for the improvement of plant, animal and microorganism species for human benefit. As a consequence, modern biotechnology has greatly promoted the research and development of genetically modified (GM), or transgenic crops worldwide. To date, a large number of modified genes conferring diverse traits have been successfully transferred into crop varieties through modern biotechnology with various agronomically beneficial traits have opened a new dimension for meeting the great challenge of world food security by enhancing the efficiency of crop production. But on the other hand, the extensive environmental release and cultivation of GM crop varieties have aroused tremendous biosafety concerns and debates worldwide [6].
Flows of genes from a Genetically Modified (GM) crop to its non-GM crop counterparts (particularly conventional varieties) or to a population of weedy/wild relatives have been considerably discussed as one of the central ecological or environmental risks associated with the application of transgenic biotechnology to crop production [7]. Such environmental risks include potential adverse effects on natural biodiversity and the survival of wild populations. Assessing the consequences caused by transgene flow is challenging because it is difficult to predict the ecological effects of transgenes that are integrated into different genetic backgrounds or expressed in different ecological contexts. Indeed, plants that acquire transgenes will continue to evolve, subject to natural and artificial selection pressures in the agricultural setting and beyond. Importantly, once transgenes have moved into new populations, it is impossible to remove them from the environment if the transgenes can successfully persist and spread in the population [8]. Therefore, understanding issues such as what is gene flow, what causes gene flow, and what will be the fate of a transgene that has moved into a recipient population through gene flow, is very useful for understanding their potential consequences.
The movement of genes from one plant population to another, any medium that can move genes around will lead to gene flow. Typically, there are three avenues for gene flow to be mediated; either by pollen, seed or vegetative propagule. Pollen-mediated gene flow occurs when pollen grains travel from one plant individual to another individual resulting in fertilization. It will be primarily determined by the intrinsic biological features, particularly the pollination biology of the plant species, such as breeding systems, outcrossing rates, the amount of pollen (pollen load) produced by pollen donors, and pollen competition between donors and recipients [9]. In addition, physical or environmental conditions, such as distances between pollen donors and recipients, the strength and direction of the wind, temperature, light intensity, and air humidity, will also influence pollen-mediated gene flow to a great extent [9]. It is therefore, very important to generate such baseline biological and physical data through a science- based approach for the accurate prediction of pollen- mediated gene flow. In agricultural ecosystems, humans can play an important role in seed and vegetative-organ dispersal and migration, as would be the case of seeds or vegetative organs falling on the ground during harvesting and picking, transportation to the processing manufacturers, and trading at the local, regional, and international level. The intensity and avenues of gene flow in different crop species can vary significantly, depending on annual or perennial characteristics, the capacity for seed dormancy, the longevity of seeds or vegetative propagules during storage (under natural or artificial conditions), differences in breeding (mating) systems, the importance of such crops in national and international markets, and those parts of the crop that are consumed by humans. Given the complexity of gene movement through seeds or vegetative organs, seed- mediated gene flow and vegetative-propagule-mediated gene flow will not be discussed further in this review, but it is necessary to point out that these are very important avenues for gene flow in terms of evolutionary processes or GM-related biosafety issues.
There is great concern that the extensive cultivation of GM crops will pose potential threats to the genetic diversity of traditional crops [10]. The loss of genetic diversity of crops, in general, will reduce the capacity to breed more productive and stress-resistant crop varieties. The concerns of genetic erosion are twofold. First, the extensive adoption of GM crops may lead to rapid losses of traditional crop varieties because of the continuous replacement of the traditional varieties by more commercially advantageous GM varieties. Gene flow is a natural process that contributes to species evolution. However, in the particular case of GM crops, the flow of one or more transgenes could have adverse environmental, socio- economic, or ethical impacts. Hybridization between a GM crop and a non-GM crop or a wild relative can occur in one generation, from which the escaped transgene(s) may then integrate into the genome of the non-GM crop varieties or wild relative species through further introgression, resulting in the gradual integration of the transgene into a related plant genome (wild relatives) through consecutive backcrossing. Hybridization and introgression will promote the long-term persistence and dissemination of transgenes in populations of wild or weedy populations, and may cause unwanted ecological and environmental consequences. The pollen- mediated transgene outflow may pose more significant environmental consequences than either seed-mediated or vegetative-organ-mediated gene flow because of subsequent genetic recombination and introgression of the transgenes in populations of crops and weedy/wild relative.
Methods of gene flow from the GM crops to wild relatives mediated by pollen
Gene flow is a natural part of the biology of plant species and is important in the maintenance of genetic variation in populations, as well as in the spread of new traits among populations and across species boundaries. It typically involves the movement of pollen and is dependent upon wind or animal vectors (pollinators).
The necessary initial conditions for the most common pathway for transgene introgression include; transgenic agronomic populations (typically in designated fields, but possibly also in the adjacent area, such as road sides) in close proximity with their wild or weedy relatives that exist in the non-agronomic matrix. Overlapping flowering phonologies are co-occurring periods of time when floral organs of species/populations are receptive to cross-pollination. Given the conditions outlined above, the following events are necessary for introgression to take place (Figure 1). This is the most commonly envisioned model of introgression, which involves an initial pollen movement step of transgenic pollen flow to the wild or weedy relative. Fertilization (i.e. hybridization) Seed! Seedling! Adult transitions of the F1 hemizygous transgenic plant within the non-agronomic matrix where the wild or weedy relative population exists. Backcrossing of the F1 hemizygous transgenic plants with existing non-transgenic plants or transgenic agronomic plants, if present Charles Kwit [11].
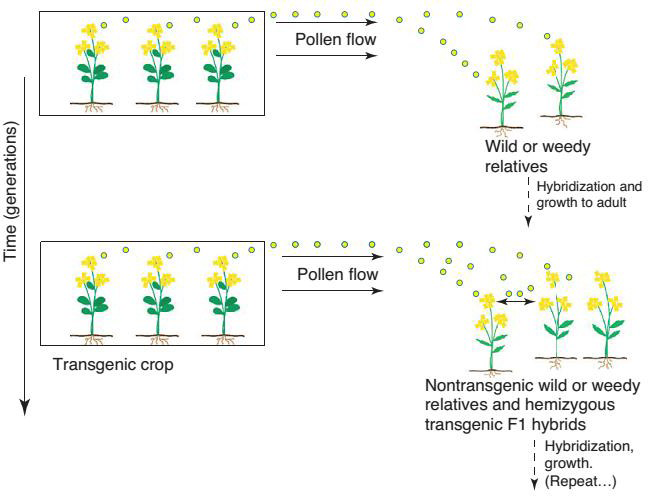
There are concerns shown by the ecologists that transgenic crops themselves may turn into weeds in subsequent generations. The main biological concern is introgression that is hybridization among distinct plant species. Evidence indicates that such genetic exchanges among wild, weed and crop plants are already occurring in the nature. The incidence of shatter cane (Sorghum bicolor), a weedy relative sorghum and the gene flow between maize and teosinte demonstrate the potential for crop relatives to become serious weeds. Two potential risks following transgene introgression from crops to their wild or weedy relatives. (i) Invasive hybrid population from introgressed (a) weedy or (b) wild population (dashed arrow representing seed dispersal) brought about by positive selection and/ or evolution; (ii) extinction of wild relative population brought about by demographic swamping (e.g. copious pollen from transgenic crops) and negative selection. We note that an introgressed coexisting weedy population might present problems for weed management when introgressed transgenes confer a selective advantage in the managed agronomic system, and that an introgressed wild population might also be of concern to managers of crop-wild relative genetic conservation [12].
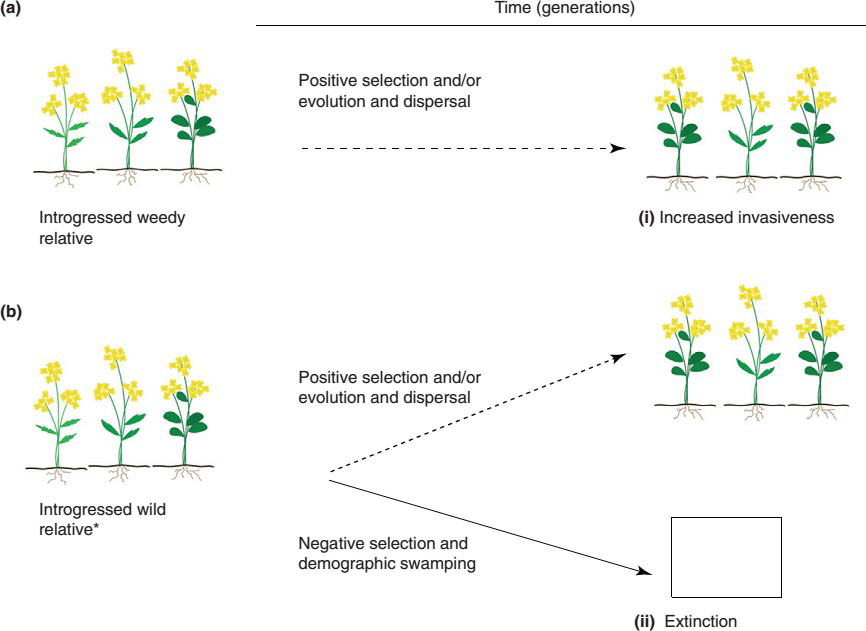
Ecological consequences of gene flow through pollen from transgenic crops to their wild relatives.
The most relevant questions relating to transgene flow and its potential biodiversity and environmental impacts should be scientifically addressed and analyzed. This will not only facilitate our objective understanding of the potential biosafety problems caused by transgene flow at various situations but also for the effective assessment and management of transgene flow and its impacts. Such knowledge will guarantee the further development of transgenic biotechnology and promote the safe and sustainable utilization of its products. The environmental impacts created by transgene escape into different recipients can vary significantly in terms of categories and magnitudes. Transgene escape from GM crops to their non-GM counterparts will have completely different consequences compared with the escape to weedy and wild relative species. Even in the latter case, different types of transgenic traits will have different effects on wild populations under different environmental conditions and human influences. Therefore, the case-by-case principle should be applied rigidly to assess the environmental impacts from transgene flow, which should be dependent upon the types of recipients (crops or wild species) that may have acquired the transgenes.
Ecological studies have shown that some GM crops are viable in natural ecosystems and can interbreed with their wild relatives. The most publicized environmental consequence is that invasive weeds may be created if GM crops modified to tolerate herbicides or to resist diseases and pests transfer their transgenes to wild or weedy relatives via gene flow. In principle, the potential environmental consequences caused by crop-to-wild or crop-to-weedy transgene flow can be effectively determined by the amount of transgenes that have outflowed to the wild and weedy populations, and by the characteristics of the GM traits that have or do not have evolutionary advantages under natural selection. When populations incorporate a GM trait likely to confer a selective advantage and are then exposed to a relevant selective pressure (e.g. pest attacks or drought/ salinity stresses), the populations will most likely exhibit an enhanced performance Ellstrand NC, Lu BR [6, 13] leading to unwanted environmental consequences. Even though genetically engineered crops could have a number of agronomical or environmental benefits, such as an increase in yield or a decrease of the use of pesticides or fertilizers, there are serious concerns about the possible consequences of the escape of transgenes into the environment. Examples of the risks mentioned in the context of gene flow from GM plants are:
Creates new weeds
The release of organisms with novel phenotypes bears similarities to the introduction of non-native species. Many well-documented examples reveal non-native plants, including kudzu and purple loosestrife, becoming aggressive weeds with devastating environmental and economic consequences. Sometimes, introduced plants invade successfully because no insect herbivores attack them. Consequently, insect-resistant transgenic plants might be more likely to become invasive weeds than would the parental variety [12].
Moreover, hybridization between a transgenic crop and a related non-crop plant can spread novel traits to additional species, which further complicates the analysis of the risk of creating new weeds. For example, Norman Ellstrand and his colleagues at the University of California at Riverside found that 12 of the world’s 13 most important food crops hybridize with wild relatives in some part of their distributions. If a transgenic crop can hybridize with nearby wild relatives, the transfer of genes will be virtually inevitable once farmers plant the crop on a commercial scale. As a result, insect- resistance traits could create aggressive weeds from either the crop or, more likely, from related non-species. Still, some scientists argue that the escape of insect-resistance genes into wild populations would not substantially increase the population growth rate of wild plants [14, 15].
Without direct field results indicating how herbivore- resistant crops could generate weeds, some insight can be gained from ecological experiments on herbivory in natural plant populations. To quantify the impact of herbivores, scientists often manipulate the number of herbivores attacking plants and then measure changes in plant growth or reproduction. To speculate on the future impacts of transgenic crops, an investigator must summarize a large collection of such studies across many species and situations, which can be done with a statistical approach called a meta-analysis. Peter Kareiva of the Marine Fisheries Service applied this method to results reported in different publications that involved 52 different plant-herbivore combinations. He found an extraordinarily large average effect of herbivory: Plants protected against invertebrates’ primarily insect herbivores produced more reproductive structures than did 81 percent of unprotected plants. Thus any herbivore-resistance trait is likely to confer a substantial advantage, which could easily increase the occurrence of weeds. Although increases in seed production do not always translate into enhanced weediness, limitations on seed production do constrain many natural plant populations, at least in some years. Consequently, insect-resistance genes could cause non-domesticated relatives of transgenic crops to become weeds [16].
The analogy between the commercial-scale planting of transgenic crops and the introduction of non-native plant species suggests several additional factors to consider when assessing the risks associated with transgenic crops. First, the vast majority of introduced plant species cause no serious environmental problems. Accordingly, most transgenic crops will probably pose little threat to the environment. As in plant introductions, though, a small percentage of transgenic varieties might become serious pests that cause vast economic and environmental damage. Second, even introduced plants that become aggressive weeds often remain relatively uncommon for long periods before becoming problematic. For example, Mimosa pigra, or catclaw mimosa, was a minor weed in Australia for about a century before it expanded dramatically and excluded other plants over large areas. In addition, Michelle and colleagues searched historical reconstructions of plant invasions and found many examples of remarkably long lags between the time of a plant’s introduction and the detection of weed spread. Although there would be enormous pressure to discontinue monitoring efforts of transgenic crops if weeds have not been detected after many years, we must remember that detection lags can be quite long and that effective monitoring might require 30 or more years of sampling [16, 17].
The second major ecological concern surrounding insect-resistant transgenic crops from pollen is that they might harm non-target animals. For example, a plant that is toxic to Colorado potato beetles could conceivably also be toxic to non pest beetles or to beetles that actually benefit farmers, including ladybird beetles. Losey and his colleagues studied the effects of pollen produced by transgenic corn that resists lepidopteran pests, including the European corn borer. Plant breeders can transfer a gene from a bacterium called Bacillus thuringiensis into corn, which causes the corn plant to produce an insecticidal compound, commonly called Bt toxin. In fact, there are several distinct Bt toxins, each capable of binding to receptors in the midgut of particular groups of insects, but humans and other vertebrates do not express these receptors. Wind pollinates corn, and its copious pollen can move up to 60 meters, coating the surfaces of neighboring non-crop plants. Thus, non-target insects, including monarch larvae that feed on milkweed plants, consume some of the windblown corn pollen [16, 18].
Losey and his colleagues coated milkweed leaves with transgenic corn pollen in quantities similar to those observed in nature. Only 56 percent of the monarch larvae survived when fed milkweed plants coated with transgenic corn pollen, whereas 100 percent of them survived when the plants were coated with non-transgenic corn pollen. Given that roughly half of the monarchs in the United States spend their summer months feeding on milkweeds in corn- producing regions, the effects on monarch populations could be substantial. Nevertheless, critics of this study point out that the toxins in transgenic corn’s pollen might become inactive more quickly in the field than in a laboratory setting [17, 18].
Loss of genetic diversity in crop landraces and wild relatives
There is great concern that the extensive cultivation of GM crops will pose potential threats to the genetic diversity of traditional crops [10]. The loss of genetic diversity of crops, in general, will reduce the capacity to breed more productive and stress-resistant crop varieties. The concerns of genetic erosion are twofold first, the extensive adoption of GM crops may lead to rapid losses of traditional crop varieties because of the continuous replacement of the traditional varieties by more commercially advantageous GM varieties. For example, after only a decade of adopting GM cotton, the current cultivation area of insect-resistant GM cotton (Bt) comprises more than 70 % of the total cotton cultivation area in China, and more than 40 % of the total cotton cultivation area in India [30, 31]. A similar situation is found with GM soya beans and GM oilseed rape in northern America. However, the counterargument is that losses to genetic diversity in traditional crop varieties have already taken place prior to the introduction of GM crops into modern agriculture, with its landscape fragmentation, adoption of improved crop varieties, and degradation of the agricultural environment. Second, the spread of transgenes from a GM crop variety to non-GM traditional varieties through gene flow may change the integrity of the traditional varieties if the transgenes have a selective advantage. During the process of cultivation and seed production, hybrids containing any beneficial transgenes may gradually accumulate unintentionally during selection to ultimately replace the original genotypes of the traditional varieties Wild relatives of crop species are widely viewed as valuable resources of genetic diversity for future breeding [6]. Even if these wild relatives are somewhat weedy, germplasm experts believe that these reservoirs of diversity should be protected from population extinction and genetic “swamping,” which results from a heavy influx of crop genes. To some people, the more presence of transgenes in the wild germplasm of crop relatives represents a form of “contamination” or “genetic pollution”. There are two scenarios for the unwanted effects of transgenes on genetic diversity. First, it is theoretically possible that strong selection for fitness- enhancing transgenes could generate selective sweeps in which portions of the crop genome that are linked to these transgenes displace corresponding portions of wild genomes [6, 32]. This process is expected to be more common in self-pollinating species than in out-breeders, which have a greater potential for the mixing and dilution of crop alleles during sexual reproduction.
Also, selective sweeps could be favoured by clonal reproduction, which might allow more vigorous transgenic crop-wild hybrids to out-compete the non-GM plants at the local level. The potential for rapid selective sweeps in most self-pollinating plants seems remote because 1) few transgenes seem likely to confer fitness benefits that are strong enough to lead to selective sweeps in wild populations, and 2) the extent of pollen-mediated gene flow is typically very low. However, massive transgene flow to wild relatives through recurrent pollination may pose threats to wild germplasm, particularly for out-breeders. Second, in some situations, a large influx of fitness-reducing transgenes could contribute to population declines or even local extinction of small, isolated populations of wild plants that occur near the crop [33]. In populations of 100 individuals or more, frequencies of fitness-reducing transgenes would diminish due to the purifying force of natural selection. Therefore, current information suggests that gene flow from self- pollinating GM crops may not threaten the genetic diversity of wild and weedy relatives to a greater extent than the current gene flow from conventional varieties [6, 32].
Evolutionary Consequences of Gene Flow through Pollen from Transgenic Crops to their Wild Relatives
The effect of gene flow is to reduce genetic differences between populations, thereby preventing or delaying the evolution of the populations in different geographical areas into separate species. As Ellstrand, et al. [34] already pointed out, gene flow between a crop and its wild relatives can have two potentially harmful ecological/evolutionary consequences: enhanced weediness in the wild/weedy relatives by introgression of certain crop traits and extinction of wild relatives through genetic swamping or outbreeding depression. For example, the evolution of enhanced weediness in one of the world’s worst weeds, Sorghum halepense, is assumed to be a result of introgression from the crop Sorghum bicolor. Transgenes may enhance these chances, depending on the specific traits encoded.
The evolution of novel weeds and invasives need not involve domesticates at all. Natural gene flow and subsequent admixture involving only non‐domesticated taxa are known to have preceded the evolution of dozens of new weedy and invasive lineages [35]. Examples include plants, animals and microorganisms. Recently, an increasing number of studies have revealed that even within‐species admixture appears to have played a role in the evolution of many invasive lineages [36]. In that case, admixture depends on multiple introductions [37]. Likewise, a small, but growing, number of examples of invasives that evolved from intertaxon hybridization have been shown to be the result of multiple, independent hybridization events. One such example is the tumbleweed, Salsola ryanii, evolved in California as recently as a few decades ago and is certainly no more than a century old. An allohexaploid derivative of hybridization between Salsola australis and Salsola tragus, its known range expanded from a few isolated populations in California’s Central Valley to a broad distribution in that Valley as well as California’s coastal valleys in roughly a decade. Despite its recent origin and rapid spread, molecular analysis by Welles and Ellstrand [38] demonstrates that this neo‐invasive is the result of three independent admixture events and that gene flow among the individual lineages is just beginning, leading to within‐species admixture.
Conclusion
Gene flow is not a risk because it is a natural process and a part of evolution that happens incessantly and permanently. Transgene escape from a GM crop to its non- GM crop counterparts and populations of wild or weedy relatives through gene flow may pose potential biosafety problems for food and health, environment, and socio- economics and ethics. Potential environmental consequences from transgene escape essentially depends on whether or not the transgenes will express normally in wild relatives, and whether or not the transgenes will change the fitness of introgressed plants, which will determine the dynamics of populations that have maintained the transgene under selection pressure. There is general agreement that gene flow from pollen of GM crops to sexually compatible wild relatives can occur. Experimental studies have shown that GM crops are capable of spontaneously mating with wild relatives, however at rates on the order of what would be expected for non-transgenic crops. Much empirical information about crop-wild relative hybridization is now available indicating that such hybridization occurs when sexually compatible wild relatives are present in close proximity to the crop, albeit at low (and variable) rates. Hybridization between conventional (non-GM) crops and their wild relatives has occasionally caused problems in ecological and evolutionary times. There is no evidence yet, that GM crops pose any greater risk than do non-GM crops, but our knowledge of the fitness consequences of transgenes in wild populations is incomplete. It is difficult to judge a priori whether a transgenic phenotype will have a special fitness advantage relative to a non-transgenic counterpart and if an advantage exists, whether this will result in increased weediness. The data published so far indicate that serious ecological consequences have not been observed. Based on biological knowledge, the possibility of transgene escape can be assessed effectively, as well as the potential environmental consequences created by any transgene outflow.
References
-
Thomson JA (2001) Horizontal transfer of DNA from GM crops to bacteria and to mammalian cells. Journal of Food Science 66(2): 188-193.
-
Hartl DL, Clark AG (1989) Principle of Population Genetics. 2nd Edition. Sinauer Associates Inc., Sunderland MA, USA
-
Frankham R, Briscoe DA, Ballou JD (2002) Introduction to Conservation Genetics. Cambridge University Press.
-
Huang JS, Rozell (2003) Plant biotechnology in China. Science 295(5555): 674-676.
-
Hatti-Kaul R, Tornvall U, Gustafsson L, Borjesson P (2007) Industrial biotechnology for the production of bio-based chemicals: A cradle-to- grave perspective. Trends in Biotechnology 25(3): 119-124.
-
Ellstrand NC (2003) Dangerous liaisons? When cultivated plants mate with their wild relatives, the John Hopkins University Press, Baltimore and London.
-
Committee on Scientific Evaluation of the Introduction of Genetically Modified Microorganisms and Plants into the Environment (1989) Field Testing Genetically Modified Organisms: Framework for Decision. National Research Council (NRC), National Academy Press, Washington DC, USA.
-
Johnston WJ, Vu DQ, Giband M, Lu BR, Andow DA, et al. (2008) Environmental risks associated with gene flow from transgenic cotton in Viet Nam. Environmental Risk Assessment of Genetically Modified Organisms: Challenges and Opportunities with Bt Cotton in Viet Nam. In: Andow DA, et al. (Eds.), CABI Publishing, Wallingford, UK, pp: 10.
-
Rong J, Song Z, De Jong TJ, Zhang X, Sun S, et al. (2010) Modelling pollen‐mediated gene flow in rice: Risk assessment and management of transgene escape. Plant Biotechnology Journal 8(4): 452-464.
-
Engels JMM, Ebert AW, Thormann I, De Vicente MC (2006) Centres of crop diversity and/or origin, genetically modified crops and implications for plant genetic resources conservation. Genetic Resources and Crop Evolution 53: 1675-1688.
-
Charles Kwit, Hong S, Moon Suzanne I, Warwick C, Neal Stewart Jr (2011) Transgene introgression in crop relatives: molecular evidence and mitigation strategies. Trends in Biotechnology 29(6): 284-293.
-
Lu BR (2006) Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering, Institute of Biodiversity Science, Fudan University, Shanghai, China. Genetically Modified Organisms in Crop Production and Their Effects on the Environment: Methodologies for Monitoring and the Way Ahead: Expert Consultation: Report and Selected Papers pp: 85.
-
Lu BR, Snow AA (2005) Gene flow from genetically modified rice and its environmental consequences. Biosciences 55(8): 669-678.
-
Vacher C, Kossler TM, Hochberg ME, Weis AE (2011) Impact of Interspecific Hybridization between Crops and Weedy Relatives on the Evolution of Flowering Time in Weedy Phenotypes. PLoS ONE 6(2): e14649.
-
Ellstrand NC, Meirmans P, Rong J, Bartsch D, Ghosh A, et al. (2013) Introgression of crop alleles into wild or weedy populations. Annual Review of Ecology, Evolution and Systematics 44: 325-345.
-
Marvier M (2001) Ecology of Transgenic Crops: Genetically engineered plants might generate weed problems and affect non-target organisms, but measuring the risk is difficult. American Scientist 89(2): 160-167.
-
Bauer Panskus A, Miyazaki J, Kawall K (2020) Risk assessment of genetically engineered plants that can persist and propagate in the environment. Environ Sci Eur 32: 32.
-
Yu HL, Li YH, Wu KM (2011) Risk Assessment and Ecological Effects of Transgenic Bacillus thuringiensis Crops on Non‐Target Organisms F. Journal of Integrative Plant Biology 53(7): 520-538.
-
Gressel J (2000) Molecular biology of weed control. Transgenic Research 9(4-5): 355-382
-
Ye X, Al Babili S, Klöti A, Zhang J, Lucca P, et al. (2000) Engineering the provitamin A (beta-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 287(5451): 303-305.
-
Chen G, Wang Z, Liu QQ, Xiong F, Gu YJ, et al. (2006) Development and substance accumulation of caryopsis in transgenic rice with antisense Wx gene. Rice Science 13(2): 106-112.
-
Snow AA, Pilson D, Rieseberg LH, Paulsen MJ, Pleskac N, et al. (2003) A Bt transgene reduces herbivory and enhances fecundity in wild sunflowers. Ecology Applications 13: 279-286.
-
Committee on Environmental Impacts Associated with Commercialization of Transgenic Plants (2002) Environmental Effects of Transgenic Plants: The Scope and Adequacy of Regulation. National Research Council (NRC), National Academies Press, Washington DC, USA.
-
Snow AA, Andow DA, Gepts P, Hallerman EM, Power A, et al. (2005) Genetically modified organisms and the environment: Current status and recommendations. Ecology Applications 15(2): 377-404.
-
Jenczewski EJ, Ronfort (2003) Crop-to-wild gene flow, introgression and possible fitness effects of transgenes. Environmental Biosafety Research 2(1): 9-24.
-
Song X, Yan J, Zhang Y, Li H, Zheng A, et al. (2021) Gene Flow Risks From Transgenic Herbicide-Tolerant Crops to Their Wild Relatives Can Be Mitigated by Utilizing Alien Chromosomes. Front Plant Sci 12: 670209.
-
Xia HH, Zhang W, Wang X, Yang F, Wang J, et al. (2016) Ambient insect pressure and recipient genotypes determine fecundity of transgenic crop-weed rice hybrid progeny: implications for environmental biosafety assessment. Evolutionary Applications 9(7): 847-856.
-
Lu BR, X Yang, NC Ellstrand (2016) Fitness correlates of crop transgene flow into weedy populations: a case study of weedy rice in China and other examples. Evolutionary Applications 9(7): 857-870.
-
Ellstrand NC (2012) Over a decade of crop transgenes out-of-place. In: CA Wozniak, and A McHughen, eds. Regulation of Agricultural Biotechnology: The United States and Canada, pp: 123-135.
-
James C (2006) Global Status of Commercialized Biotech/GM Crops: ISAAA Brief No. 35. International Service for the Acquisition of Agri-Biotech Applications (ISAAA), Ithaca NY, USA.
-
Wu KM (2007) Monitoring and management strategy for Helicoverpa armigera resistance to Bt cotton in China. Journal of Invertebrate Pathology 95(3): 220-223.
-
Gepts P, Papa R (2003) Possible effects of (Trans) gene flow from crops on the genetic diversity from landraces and wild relatives. Environmental Biosafety Research 2(2): 89-103.
-
Haygood R, Ives RA, Andow DA (2003) Consequences of recurrent gene flow from crops to wild relatives. Proceedings of the Royal Society: Biological Sciences 270(1527): 1879-1886.
-
Ellstrand NC, Prentice HC, Hancock JF (1999) Gene flow and introgression from domesticated plants into their wild relatives. Annual Review of Ecology and Systematics 30: 539-563.
-
Schierenbeck KA, Ellstrand NC (2009) Hybridization and the evolution of invasiveness in plants and other organisms. Biological Invasions 11: 1093-1105.
-
Rius M, Darling JA (2014) How important is intraspecific genetic admixture to the success of colonising populations? Trends in Ecology and Evolution 29(4): 233-242.
-
Bock DG, C Caseys RD, Cousens MA, Hahn SM, Heredia S, et al. (2015) What we still don’t know about € invasion genetics. Molecular Ecology 24(9): 2277-2297.
-
Welles SR, Ellstrand NC (2016) Genetic structure reveals a history of multiple independent origins followed by admixture in the allopolyploid weed Salsola ryanii. Evolutionary Applications 9(7): 871-878.
- Lessons to Learn: Trees are More than the Lungs of the World
- Community Forestry Enterprises as a Model for Sustainable Forest Development: The Case Of The "Baja Tarahumara" in Chihuahua, Mexico
- Ecological and Socio-Economic Impacts of Chromolaena odorata and Mesosphaerum suaveolens, Two Invasive Alien Species in Central and Southern Benin, West Africa
- Epigenetic Sustainability: Modeling the Human Factor as a Natural Resource through Science 4.0 and the NR3C1 Biological Pilot
- Growth-at-Risk: A Framework for Assessing Economic Vulnerability
- The Rural Territory as a Socioecological System for the Management of Public Policy for Sustainable Rural Development