Influence of Aspect and Elevational Gradients on The Regeneration of Festuca Macrophylla and Helicrysum Spelindidum Plant Species in GCCA, Central Highlands of Ethiopia
Natural regeneration is an important process that ensures the flow genes and thus preserves the diversity of life in ecosystems. However, natural regeneration is constrained by many factors including biotic, climatic and edaphic factors. The aim of this work was to assess the effects of aspect and elevational gradient on the natural regeneration potential of two competing species in Guassa Community Conservation Area in North Central Ethiopian highlands. We stratified our study area into eight blocks based on aspect and elevational gradient and conducted shrub removal experiment on 160 plots (5m x 5m). From these plots, we conducted vegetation survey following the modified Braun-Blanquette methods every three months. We used two elevational (<3350 m/lower and >3350 m/upper elevation) and four aspect categories (northeast, northwest, southwest and southeast) to compare the seeding density of the target species and species composition studies. On these plots, all vascular plants were recorded and the data was analyzed by two-way ANOVA using SAS statistical software version 9.0.1. In addition, we used LSD test for mean separation test. We computed the similarities in composition, density, abundance and species similarity between soil seedbank and the aboveground vegetation. Results showed that a total of 31 species, representing 27 genera and 16 families were recorded. The families with the highest number of species were Asteraceae (18 species, 22.5%), Fabaceae (3 species, 10%), and Poaceae (3 species, 10%). Of the total families, the dicots constituted 87.5% and monocots 12.5%. Species abundance was negatively correlated with elevation, with fewer species occurring at higher elevation. Meanwhile, species abundance was positively correlated with both northwest and northeast aspect. Treatments done during the study also highly influenced the recovery of vegetation species. High species similarity was also observed between below and aboveground vegetation.
Introduction
For the management and protection of biodiversity, it is essential to have knowledge of the plant diversity and regeneration status of specific vegetation type. Natural regeneration is a key element of the dynamics of the specific ecosystem and is crucial for the conservation and preservation of biodiversity [1]. The diversity of their species, variety, and intricate community organization are among the most significant traits of tropical and subtropical vegetation, along with regeneration [2]. According to Pala [3], a given species’ ability to regenerate successfully depends on the structure of its overall populations as well as the availability of sufficient numbers of seedlings, saplings, and adults.
Ecosystem’s species richness, population structure, and natural regeneration are all impacted by both natural and man-made activities. Invasive species, floods, fires, and storms are examples of natural causes, whereas illegal logging, excessive use of natural resources, encroachment, harvesting (overexploitation) and domestic grazing are examples of human-caused factors that can harm the natural ecosystems. The conservation of biodiversity and human use of resources in developing countries are at odds [4], and degradation and destruction of habitats as a result of human activities are the main causes of global biodiversity loss [5].On the other hand, understanding the distribution and patterns of vegetation growth as well as the influencing factors in those areas is crucial and has been researched by numerous researchers [6, 7].
According to Dawes and Short [8], the type of soil and the amount of rainfall only have a little impact on the size of hill slopes, where topography is the primary governing factor for vegetation growth. The three primary topographic parameters that regulate the distribution and patterns of vegetation in mountainous locations are elevation, aspect, and slope [9]. Elevation is the most crucial of these three variables [10]. The microclimate, and consequently the large-scale spatial distribution and patterns of vegetation are frequently influenced by elevation, aspect, and slope [10].
In the central highlands of Ethiopia, there are no officially protected areas, but there is a community where natural resources are protected for its sustainable use and for benefits of the entire community. The nature of community conserved areas depends on the ecological characteristics in sites, species diversity and regeneration status of species. Natural regeneration is essential for preservation and maintenance of biodiversity in community conserved areas. Depending on management objectives, it is important to maintain the process of ecosystem renewal by appropriate natural and assisted regeneration. The clear felling of grasses accelerates loss of seedlings and saplings as well as disturbs the natural condition of the ecosystem [11].
Knowledge of plant diversity and natural regeneration status of certain ecosystems provides basic information for ecosystem management and biodiversity conservation [12]. Natural regeneration is a fundamental component of ecosystem dynamics and is essential for the conservation and maintenance of biological diversity [13, 1, 14], as well as for identifying priority plant species for conservation [15]. Regeneration management is an essential component for sustainable ecosystems as it ensures future growing stocks [16]. Successful regeneration of a species can be predicted by overall population structure as well as sufficient numbers of seedlings, saplings and adults [3, 17].
For the conservation and management of plant diversity in biodiversity conservation areas, information on plant diversity is considered essential because of its potential importance and implications [18]. Many researchers [19, 20, 21, 22, 23, 24, 25] have carried out various studies on GCCA but the regeneration studies were not investigated in the biodiversity conservation areas of GCCA, Northern Ethiopia.
Thus, the aim of this study was to assess the effect of aspect and elevation on the natural regeneration status, composition and diversity of herbaceous species in conservation area namely GCCA. In the study site, the ground is mainly covered with herbs and shrubs. Therefore, assessing the regeneration status, composition and diversity of Festuca grass in the study area is highly valuable to understand the current status of the conservation area and to predict future conservation strategies and design [26].
Materials and Methods
Study Area
The study area is located in the Amhara Regional State, North Shewa Zone, in the central portion of the Ethiopian highland massif in an area locally known as Menz Gera Midir district. It is located at a latitude of 100 15’–100 27’N and longitude 390 45’–390 49’E, 295 km north-east from Addis Ababa (capital city of Ethiopia) [22]. The guassa range comprises 111 km2 and lies at an altitude range of 3200 to 3700 m above sea level [19].
The high plateaus of Ethiopia formed 45 million years ago from tertiary uplift followed by volcanism and form basalt layers [27, 28]. This long-term volcanic activity resulted in the building up of the northern and central highlands of Ethiopia which include GCCA. Ethiopia is located in the horn of Africa stretching from 3o N to 15o N latitude and from 33o E to 48o E longitude with an area coverage of 1,104,300 km2. It is a country of great geographical diversity with high altitude and physical-geographical variations. It is one of the 25 richest countries in biodiversity in the world, where two of the 36 biodiversity hotspots in the world [29, 30], namely the East Afromontane and the Horn of Africa, trespass through its mountain and lowland areas. Ethiopia consists of three major high plateau regions (the North-western Highlands, the Central Highlands and South-eastern Highlands) divided by the Great Rift Valley and by the Abay (Blue Nile) Gorge. From these regions guassa area is located in the Central Highlands.
GCCA is characterized by its plateau which cross-cut by various gorges and river valleys that flow west and eastwards. It straddles the catchment basins of two major rivers in East Africa – the Abay (the Blue Nile) and the Awash, and thus its conservation is critical to the health and functioning of those headwaters [31]. The swamps of the area increase water holding capacity and also limit runoff in the rainy seasons, thus providing flow all year round. Generally the soil of the guassa area is deep and humic. However, on higher ground, the soil is shallow, and highly mineralized [20]. The climate in the guassa area varies widely due to the differences in elevation and the size of the mountain block. The annual mean rain fall is 828mm. Temperatures are characterized by mild days and cold nights. In the drier months (December to February), daytime temperatures can rise to 25 0C, while night time temperatures can drop to -7 0C [32, 22]. The area gets its name from the “guassa grass” Festuca macrophylla, which is highly valued by the local community. The guassa area is a common property resource area managed by the community for various uses such as grazing land, essential source of income for the local community, firewood collection, and the cutting of guassa grass for various purposes such as thatching, making household equipment, baskets, painting brushes, mattresses and shepherds’ raincoats and farm implements like ropes and whips.
GCCA is a biodiversity hotspot, containing several endemic and threatened species [21]. The area is rich in biodiversity and contains several endemic and threatened species of flora and fauna species [20]. The vegetation of the guassa area is characterized by high afro-alpine vegetation with different types of habitats, namely: Euryops-Alchemilla shrubland, Festuca grassland, Euryops-Festuca grassland, Helichrysum - Festuca grassland, Swamp Grassland and Erica moorland [33, 32]. GCCA is a place for important and endemic plant species including guassa grass (F. macrophylla), giant Lobelia (Lobelia rhynchopetalum), Kniphofia foliosa and Alchemilla species [20, 21]. There are about nine (which accounts to 23% mammal fauna of the country) endemic fauna including the Ethiopian wolf (Canis simensis), gelada baboon (Theropethicus gelada) and Ethiopian Highland hare or Abyssinian hare (Lepus starcki) and about 114 (12% of 861 bird species) have been recorded in the GCCA (Ashenafi and Leader-Williams, 2005). The guassa area is also home to the world’s largest concentration of the globally endangered Ankober serin (Serinus ankoberensis) and the spot-breasted plover (Vannellus melocephalus) [19].
The human population is predominantly of Amhara ethnicity and Orthodox Christian religion. The main economic activity of the people is farming which involves mainly crop production and livestock husbandry. The community can generate income from off-farm activities such as petty trading, selling local beer (‘Tella’), liquor (‘Arekie’) and local green tea. Wool processing (known in sheep and cattle rearing) is the most common off-farm activity of the Guassa community. The area is regarded as food insecure, and in many years, parts of the population depend on the national food aid program (safety-net) [19]. The guassa area is currently managed by nine farmers’ associations, local institutions that were created throughout Ethiopia in 1975 for rural administration. The community protects the area by promulgating various regulations that restrict the use of natural resources by the communities [20].
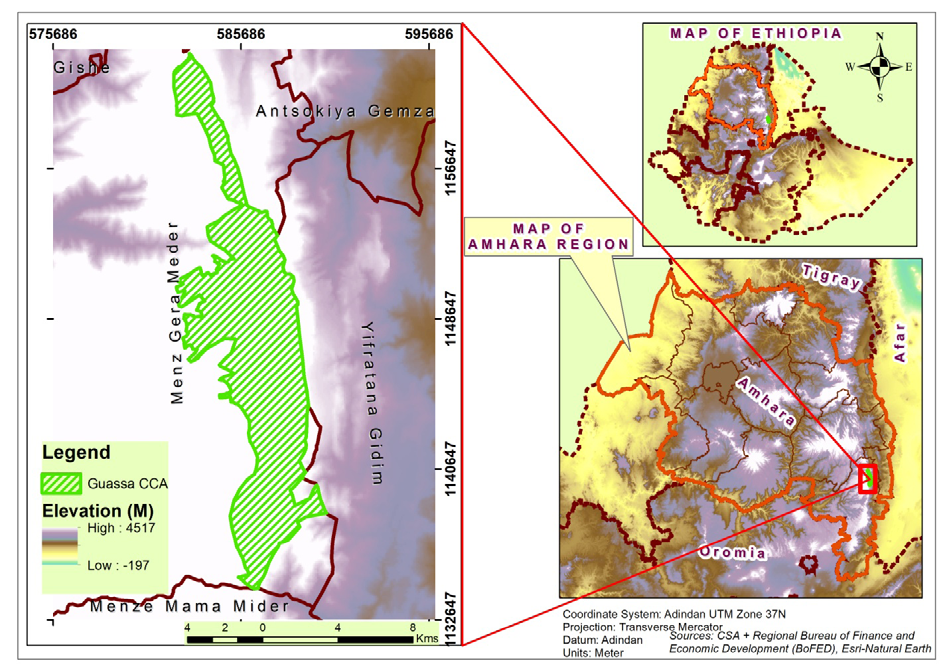
Figure1: Map of Ethiopia showing the GCCA.
Data Collection Procedure and Experimental Design
First, the vegetation in the study area was stratified and purposefully divided into eight blocks based on elevation and aspect. Each block has twenty 5 m x 5 m established plots, thus we have a total of 160 plots. Vegetation data were collected from each (160) sampling plots. We used a block design that allowed us to explore removal and regrowth of shrubs across a range of environmental variability. We organized our removal according to aspect and altitudinal gradients. We also categorized aspects in to four as NE, NW, SE and SW to compare the seedling density for the target species along elevation gradient. Elevation in GCCA ranges from about 3,000 m – 3550 m.a.s.l with the bulk of the area falling between 3250 – 3550 m. Therefore, we further stratified these plots according to two elevational categories (<3350 m/lower and >3350 m/upper elevation) to test whether removal and subsequent regrowth will respond differently at low and high elevations.
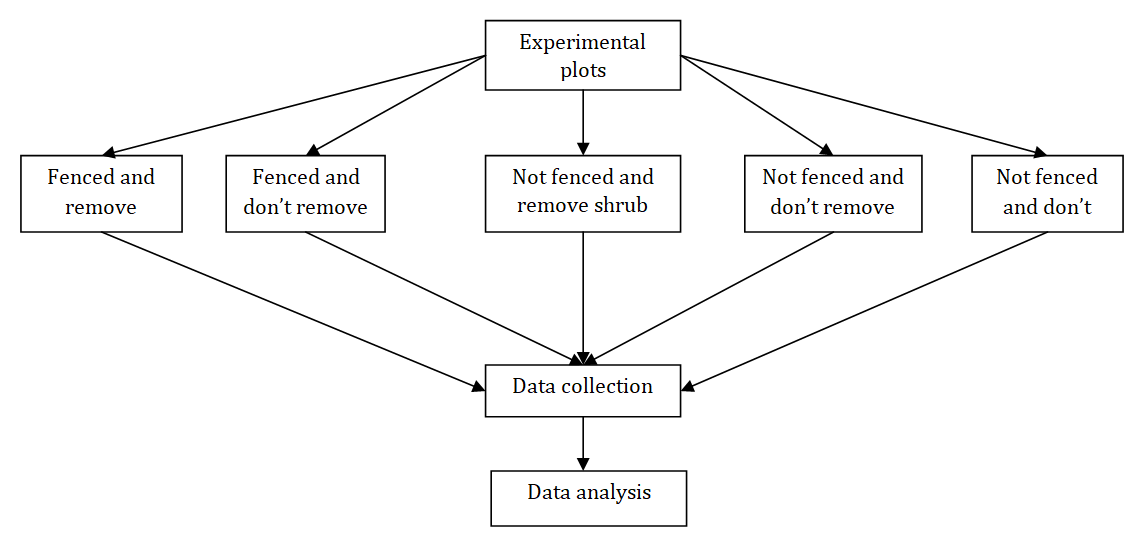
Plate 1: A schematic diagram of the experimental treatments showing data collection and analysis procedure.
Our experimental treatments were fenced and open plots. In each blocks, we have eight fenced and twelve unfenced plots. Therefore, we have a total of 160 (8 fenced plots * 8 blocks + 12 unfenced plots * 8 blocks) permanent plots from both fenced and unfenced treatments. We used meshed wire that can preclude herbivores except insects. We assumed insect herbivory is the same for both fenced and unfenced treatments.
Once we established the plots, we then classified the collected data in the experimental treatments in to two main rainy seasons (Meher) between June to September and minor rainy season (Belg) in February, March and April. Then the measurements taken during this time were categorized into five groups; 1 (Fenced remove shrub/FR), 2 (Fenced don’t remove shrub/FN), 3 (Not fenced remove shrub/ NR), 4 (Not fenced don’t remove shrub/NN) and 5 (Not fenced don’t remove shrub harvest grass/NNH). After that, we completely removed all the above ground Helichrysum biomass by cutting for the plots that require removal of the shrub and guassa grass. Members of the local community were trained and monitored the sites every month for eighteen consecutive months. During the monitoring abundance data (count) were collected for analysis. Altitude, longitude, latitude and aspect were measured and recorded for each plots using GPS.
Data Analysis Techniques
Before conducting the data analysis, all of the gathered data were organized in Excel and checked for errors, normal distribution, and homogeneity of variance. The abundance of Festuca macrophylla and Helichrysum splendidum along an altitudinal gradient was determined using SAS statistical software version 9.0.1. All the collected data was fitted into a general linear model (GLM) and analyzed using two-way ANOVA (Analysis of variance). Fisher’s least significant difference (LSD) test was employed to separate the means when significant F values were found at a significance level of p≤ 0.05.
We calculated species abundance by totaling the number of seedlings per sample area. The composition, density and frequency of seeds were determined from the germination data recorded in the sample plots. The density of seeds was derived from the total number of seeds recovered after cutting [34]. Density is a count of the numbers of individuals of a species within the plots [35]. It refers to the total number of stems of a species ha -1 which was calculated by summing up all the stems across all sample plots (abundance) and translated to hectare base for all the species encountered in the study plots. Afterwards, the sum of individuals per species is calculated in terms of species density per convenient area unit, such as hectare [34]. It was calculated using the following formula.
The species richness of above ground vegetation and soil seed bank in each soil profile was counted. Sorensen coefficient of similarity Sorensen [36] was used to compare the similarity between soil seed bank compositions with the aboveground vegetation and it is given as:
Where, a = number of species common to both soil seed bank and aboveground vegetation; b = number of species recorded only in aboveground vegetation; and c = number of species recorded only in soil seed bank. The closer Sorensen’s coefficient of similarity to 1 the more similar the sites are. This similarity coefficient was selected because it gives weight to species that exist in both sites.
Results
Floristic Composition
Results indicated that, a total of 31 species in 27 genera and 16 families were recorded from the study plots (Table 1). Of the 16 plant families, Asteraceae has the highest number of species (7 species, 22.5 %) followed by Fabaceae and Poaceae represented with three species each (Table 1). These three families contained 42.5% of the total species recorded in the study area and the remaining 13 families were represented only by less than two species each (Table 2). Of the total families, the dicots constituted 87.5% and monocots 12.5%.
Herbs were found to occur more abundantly (24 species, 77.42%) than shrubs (7 species, 22.58%).
The result of species composition showed that the area contains all the five “typical” afroalpine life-forms; namely Giant Rosette Plants (e.g. Lobelia rhynchopetalum), Tussock Grasses and sedges (e.g. Agrocharis melanantha, Carex monostachya, Festuca macrophylla), Acaulescent Rosette Plants (e.g. Haplocarpha rueppellii), Sclerophyllus and Dwarf-Shrubs (e.g. Erica arborea), and Cushion Plants (e.g. Helichrysum splendidum, Helichrysum formosissimum).
| No | Name of Species | Family | Abundance | Habit |
|---|---|---|---|---|
| 1 | Agrocharis melanantha Hochst. | Apiaceae (Umbelliferae) | 156 | H |
| 2 | Alchemilla abyssinica Fresen. | Rosaceae | 903 | H |
| 3 | Anchusa affinis R.Br. ex DC. | Boraginaceae | 124 | H |
| 4 | Andropogon lima (Hack.) Stapf | Poaceae | 2 | H |
| 5 | Anthemis tigreensis J. Gay ex A. Rich. | Asteraceae (Compositae) | 3 | H |
| 6 | Argyrolobium ramosissimum Bak. | Fabaceae (Leguminosae) | 3 | H |
| 7 | Artemisia abyssinica Sch. Bip. ex A. Rich. | Asteraceae (Compositae) | 140 | H |
| 8 | Bartsia longiflora Hochst.ex Benth. | Scrophulariaceae | 4 | SH |
| 9 | Carduus schimperi Sch. Bip. ex A. Rich. | Asteraceae (Compositae) | 114 | H |
| 10 | Carex monostachya A. Rich. | Cyperaceae | 3 | H |
| 11 | Cynoglosum coeruleum Hochst. ex A. DC. | Boraginaceae | 3 | H |
| 12 | Cyperus elegantulus Steud. | Cyperaceae | 61 | H |
| 13 | Erica arborea L. | Ericaceae | 9 | SH |
| 14 | Euryops pinifolius A. Rich. | Asteraceae (Compositae) | 1619 | SH |
| 15 | Festuca abyssinica, Hochst. ex A. Rich. | Poaceae (Gramineae) | 28 | H |
| 16 | Festuca macrophylla Hochst. ex A. Rich. | Poaceae (Gramineae) | 4988 | H |
| 17 | Haplocarpha rueppellii (Sci.Bip.) | Asteraceae (Compositae) | 52 | H |
| 18 | Helichrysum formosissimum Sch. Bip. ex A. Rich. | Asteraceae (Compositae) | 47 | H |
| 19 | Helichrysum splendidum (Thumb.) Less | Asteraceae (Compositae) | 1863 | SH |
| 20 | Hypericum revolutum Vahl | Hypericaceae | 3 | SH |
| 21 | Kalanchoe petitiana A.Rich. | Crassulaceae | 7 | SH |
| 22 | Lobelia rhynchopetalum Hemsl. | Lobeliaceae | 1 | H |
| 23 | Ranunculus multifidus Forssk. | Ranunculaceae | 4 | H |
| 24 | Rubus volkensii Engl. | Rosaceae | 19 | SH |
| 25 | Rumex abyssinicus Jacq. | Polygonaceae | 13 | H |
| 26 | Rumex nepalensis Spreng. | Polygonaceae | 32 | H |
| 27 | Thymus schimperi Ronniger | Lamiaceae | 867 | H |
| 28 | Trifolium polystachyum Fresen. | Fabaceae (Leguminosae) | 223 | H |
| 29 | Trifolium usambarense Taub. | Fabaceae (Leguminosae) | 114 | H |
| 30 | Urtica simensis Steudel | Urticaceae | 73 | H |
| 31 | Verbascum sinaiticum Benth. | Scrophulariaceae | 19 | H |
| Family | Genera | Species | Percentage | Classification |
| Apiaceae | 1 | 1 | 3.3 | Dicot |
| Asteraceae | 6 | 7 | 22.5 | Dicot |
| Boraginaceae | 2 | 2 | 6.6 | Docot |
| Crassulaceae | 1 | 1 | 3.3 | Docot |
| Cyperaceae | 2 | 2 | 6.6 | Monocot |
| Ericaceae | 1 | 1 | 3.3 | Dicot |
| Fabaceae | 2 | 3 | 10 | Dicot |
| Hypericaceae | 1 | 1 | 3.3 | Docot |
| Lamiaceae | 1 | 1 | 3.3 | Dicot |
| Lobeliaceae | 1 | 1 | 3.3 | Dicot |
| Poaceae | 2 | 3 | 10 | Monocot |
| Polygonaceae | 1 | 2 | 6.6 | Dicot |
| Ranunculaceae | 1 | 1 | 3.3 | Dicot |
| Rosaceae | 2 | 2 | 6.6 | Dicot |
| Scrophulariaceae | 2 | 2 | 6.6 | Dicot |
| Urticaceae | 1 | 1 | 3.3 | Dicot |
Table 1: List of species with their family and habit recorded from the study plots.
Density of above ground vegetation
The above ground plant species density per hectare (11495 ha-1) was recorded in the study plots. The species with the highest densities in descending order include Festuca macrophylla, Helichrysum splendidum, Euryops pinifolius, Alchemilla abyssinica and Thymus schimperi (Table 3).
The above ground density of Helichrysum spelindidum was found to be lower than that of Festuca macrophylla which was opposite result with soil seed bank density of the two species (Helichrysum spelindidum is more abundance than that of Festuca macrophylla), (the soil seed bank article is apparently under preparation for publication). Densities of the species per hectare are shown in (Table 3).
| No | Name of species | Count | Density (%) |
|---|---|---|---|
| 1 | Agrocharis melanantha Hochst. | 156 | 1.41 |
| 2 | Alchemilla abyssinica Fresen. | 903 | 8.14 |
| 3 | Anchusa affinis R.Br. ex DC. | 124 | 1.12 |
| 4 | Andropogon lima (Hack.) Stapf | 2 | 0.02 |
| 5 | Anthemis tigreensis J. Gay ex A. Rich. | 3 | 0.03 |
| 6 | Argyrolobium ramosissimum Bak. | 3 | 0.03 |
| 7 | Artemisia abyssinica Sch. Bip. ex A. Rich. | 140 | 1.26 |
| 8 | Bartsia longiflora Hochst.ex Benth. | 4 | 0.04 |
| 9 | Carduus schimperi Sch. Bip. ex A. Rich. | 114 | 1.03 |
| 10 | Carex monostachya A. Rich. | 3 | 0.03 |
| 12 | Cynoglosum coeruleum Hochst. ex A. DC. | 3 | 0.03 |
| 13 | Cyperus elegantulus Steud. | 61 | 0.55 |
| 14 | Erica arborea L. | 9 | 0.08 |
| 15 | Euryops pinifolius A. Rich. | 1619 | 14.59 |
| 16 | Festuca abyssinica, Hochst. ex A. Rich. | 28 | 0.25 |
| 17 | Festuca macrophylla Hochst. ex A. Rich. | 4988 | 44.94 |
| 18 | Haplocarpha rueppellii (Sci.Bip.) | 52 | 0.47 |
| 19 | Helichrysum formosissimum Sch. Bip. ex A. Rich. | 47 | 0.42 |
| 20 | Helichrysum splendidum (Thumb.) Less | 1863 | 16.77 |
| 21 | Hypericum revolutum Vahl | 3 | 0.03 |
| 22 | kalanchoe petitiana A.Rich. | 7 | 0.06 |
| 23 | Lobelia rhynchopetalum Hemsl. | 1 | 0.01 |
| 24 | Ranunculus multifidus Forssk. | 4 | 0.04 |
| 25 | Rubus volkensii Engl. | 19 | 0.17 |
| 26 | Rumex abyssinicus Jacq. | 13 | 0.12 |
| 27 | Rumex nepalensis Spreng. | 32 | 0.29 |
| 28 | Thymus schimperi Ronniger | 867 | 7.81 |
| 29 | Trifolium polystachyum Fresen. | 223 | 2.01 |
| 30 | Trifolium usambarense Taub. | 114 | 1.03 |
| 31 | Urtica simensis Steudel | 73 | 0.66 |
| 32 | Verbascum sinaiticum Benth. | 19 | 0.17 |
Table 2: Density of above ground plants in the study plots.
Impact of elevation difference on F. macrophylla and H. splendidum species abundance
The distribution of the plant species growing at the study sites varied with altitudinal range (figure 2).The five most abundant species in the study area are given in Table 1.
The abundance of the two target species (F. macrophylla and H. splendidum) in the study area was statistically significant. The pooled abundance and altitude were negatively correlated (Two-way ANOVA: P<0.0009), where abundance of these two species showed decreasing pattern as altitude increase in the sample blocks (Figure.2).
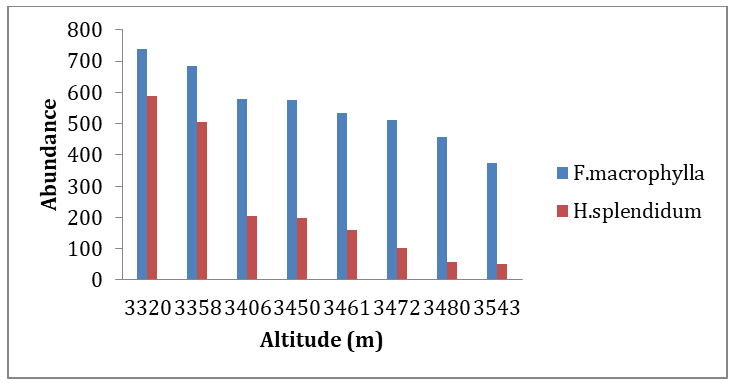
The analysis done also showed that topographical difference had a significant (P<0.05) impact on the abundance of H. spelindidum for all the three seasons in the GCCA (Table 4).
| Elevation | Meher1 | Belg | Meher2 |
|---|---|---|---|
| F. macrophylla | |||
| Lower elevation | 11.96a | 5.10a | 19.20a |
| Upper elevation | 9.78b | 4.10b | 12.34b |
| P- value | ** | ** | ** |
| H. spelindidum | |||
| Lower elevation | 8.03a | 0.60a | 10.0a |
| Upper elevation | 1.61b | 0.01b | 3.01b |
| P- value | *** | ** | *** |
Table 3: Effect of aspect on F. macrophylla and H. splendidum abundance.
Effect of aspect on the abundance of F. macrophylla and H. splendidum plant species.
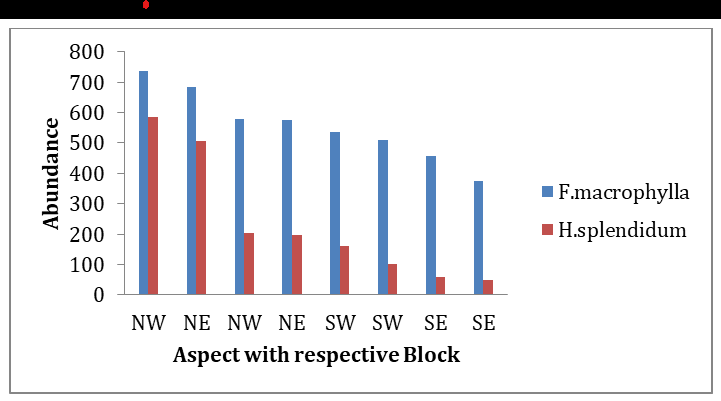
The result of the analysis done within each blocks shows that, aspect significantly affected abundance of the two species (Two-way ANOVA: P<0.0009), where both species with high abundance recorded highest at North-West and North-East direction followed by South-West and South-East directions (Figure 4).
| Aspect | Meher1 | Belg | Meher2 |
|---|---|---|---|
| F. macrophylla | |||
| North-West | 14.30a | 4.76a | 17.00ab |
| North-East | 9.62a | 5.63a | 21.40a |
| South-West | 7.65b | 3.53b | 11.68b |
| South-East | 5.90b | 3.28b | 13.00b |
| P- value | ** | * | * |
| H. spelindidum | |||
| North-West | 7.85a | 0.75a | 8.83a |
| North-East | 8.20a | 0.45ab | 11.18a |
| South-West | 0.73b | 0.00b | 1.98b |
| South-East | 2.50b | 0.03b | 4.05b |
| P- value | *** | * | *** |
Table 4: Effect of aspect on F. macrophylla and H. splendidum abundance.
Mean values followed by the same letters in each column and treatment showed no significant difference by LTD (p = 0.05). * donates = significant at P ≤ 0.05; = significant at P ≤ 0.01; * = significant at P ≤ 0.001. Table 5: Effect of aspect on F. macrophylla and H. splendidum abundance.
Regeneration status of F. macrophylla and H. splendidum species
From the findings of this work, measurements taken during data collection were also highly impacts the recovery of vegetation species and determines the abundance of mature vegetation and seeding stage of the existing shrub and grass species in GCCA. As a result the most seedling counts was observed for treatment 5 (plot not fenced don’t remove shrub harvest grass/NNH) followed by treatment 1 (Fenced remove shrub/FR), treatment 3 (Not fenced remove shrub/ NR), treatment 2 (Fenced don’t remove shrub/FN) and treatment 4 (Not fenced don’t remove shrub/NN) for both target species (Table 6). Thus the five treatments had a significant (P< 0.05) impact on regeneration status of F. macrophylla and H. splendidum species in the study area.
| Meher1 | Belg | Meher2 | |
|---|---|---|---|
| Measurement | |||
| F. macrophylla | |||
| Fenced remove shrub/FR | 15.44b | 4.91b | 19.16b |
| Fenced don’t remove shrub/FN | 1.16d | 2.79bc | 5.28c |
| Not fenced remove shrub/ NR | 8.63c | 2.81bc | 14.19b |
| Not fenced don’t remove shrub/NN | 0.88d | 1.00c | 3.78c |
| Not fenced don’t remove shrub harvest grass/NNH | 28.25a | 11.50a | 36.50a |
| P- value | *** | *** | *** |
| H. splendidum | |||
| Fenced remove shrub/FR | 7.00ab | 0.41ab | 8.06b |
| Fenced don’t remove shrub/FN | 3.00cd | 0.06b | 3.81c |
| Not fenced remove shrub/ NR | 4.41bc | 0.09b | 5.67bc |
| Not fenced don’t remove shrub/NN | 1.22d | 0.00b | 2.94c |
| Not fenced don’t remove shrub harvest grass/NNH | 8.47a | 0.97a | 12.03a |
| P- value | *** | ** | *** |
Table 5: Effect of different treatments on F. macrophylla and H. splendidum regeneration.
Mean values followed by the same letters in each column and treatment showed no significant difference by LTD (p = 0.05). * donates = significant at P ≤ 0.05; = significant at P ≤ 0.01; * = significant at P ≤ 0.001. Table 6: Effect of different treatments on F. macrophylla and H. splendidum regeneration.
Similarity between soil seed bank and aboveground vegetation
The total number of species recorded in the aboveground vegetation was 31 and the total number of species recorded in the soil seed bank was 74. Species recorded only in soil seed bank were 44, species recorded only in the aboveground vegetation were 1, and species recorded both in soil seed bank and standing vegetation were 30 (table 7). The Sorensen Similarity index was 0.6 (60% Sorenson’s coefficient).
| Aboveground vegetation (Abgv) (A) | |||
|---|---|---|---|
| No. of species present | No. of species absent | ||
| Soil seed bank SSB (B) | No. of species present | 30 | 1 |
| No. of species absent | 44 | 0 |
Table 6: Similarity between soil seed bank and aboveground vegetation.
Discussion
The total number of naturally regenerating species recorded in our experimental plots was 31 species in 27 genera and 16 families. From these families Asteraceae was the family with high species representation (7 species, 22.5%). Tadesse [37] stated that Asteraceae was among the richest family in the flora area (Flora of Ethiopia and Eritrea (FEE). According to Hedberg [38], wind dispersal is one of the main mechanisms affecting long-distance dispersal for Afroalpine landscapes. Moreover, many species of Asteraceae have umbrella shape capitula that are adapted for wind dispersal [38] and increase their opportunity for the successful establishment of their populations. The study was in accordance with results of other studies, for instance, Boditi (39 species, 17%; [39], Komto (17 species, 9.44%; [40], Dense (20 species, 27%; [39], Hugumburda-Gratkahsu (38 species, 11%; [41]).
The result of this study indicated that densities of plant species collected from the eight blocks varied greatly. The total density of naturally regenerating individuals at the study area was 11,495 per hectare. The density of Helichrysum spelindidum was found to be lower than that of Festuca macrophylla which was opposite result with soil seed bank density of the two species. Variations in density distributions can be attributed to variations in habitat preferences of species and topographic gradients forming the degree of anthropogenic influences [42].
Patterns of abundance of F. macrophylla and H. splendidum were determined against altitude. The result indicated that abundance of F. macrophylla and H. splendidum negatively correlated with altitude (P<0.0009). Altitude is the most significant factor that affects species abundance and distribution in Afroalpine region [43]. The lower elevation has high abundance compared to higher elevation gradient for both F. macrophylla and H. splendidum species. This could be due to top-climatic factors like season, seed dispersal, slope and elevation. A reduction in species abundance and richness of vegetation decline at higher elevation might also be due to harsh climatic condition, less competition, environment, a regional lack of capable species, or a multitude of disturbances caused by restriction in species’ expansion [44, 45, 43, 46].
Altitude is the independent factor that affects both the environmental and conservation area sustainability. This is the major cause of the general vegetation pattern linked with other living and non-living threaten factors [47, 48] and results of diversity and richness of grass and shrub species along with altitudinal difference. Altitude was found to be very important in many studies in influencing the plant species distribution [49, 50]. Our findings are similar to findings from mountain forests of northern China Zhang [51] and sub- alpine mountainous zone in central Japan [51]. As described by Hedberg [38] the Afoalpine grassland is “summer every day and winter every night.” The herbaceous plants in the Afroalpine grassland have almost completely different life forms from the lower elevation plants. This includes giant rosette, tussock grasses, acaulescent, Sclerophyllus and cushion plants.
Altitude was the main variable that determines the vertical distribution of vegetation whereas the horizontal distribution was affected by aspect [48]. Moreover, variations of species richness occurred among the vegetation could be attributed to the area sampled Hill and Curan [53] and large range of altitude [54], climate variability [55] and disturbance [56]. The Ethiopian vegetation was highly influenced by climate, which was associated with elevation [57]. Pollen studies in Kenya [58] and northern Ethiopia [59] indicated that the flora of East Africa has experienced several changes due to climate change and anthropogenic disturbances.
Climate, topography, aspect, inclination of slope, wind direction, soil type, and land use pattern are recognized as the major contributor to vegetation type [60]. From our findings, aspect also significantly affects species abundance. High number of F. macrophylla and H. splendidum recorded along North-West and North-East direction followed by South-West and South-East directions. North-East and North- West faces are observed as the paramount habitat for the growth of plants in the study area. Furthermore, insolation and light intensity also put down a range of microclimates at the multifaceted landscape. These factors are responsible for the diverse type of vegetation at different mountain aspects [61]. The variation in vegetation between North and South aspects could be attributed to difference in reception of solar radiation [62, 63]. South-facing slopes receive more sunlight and become warmer. Therefore, such slopes are less favorable for plant growth. However, north-facing slopes hold moisture and remain cold as well as humid, thus supporting moisture loving plants Nevo [64]; Maren [60] which is a case in the Afroalpine vegetation of the study area. On the other hand, the high number recorded in north directions may be attributed to the fact that seed dispersed by wind and also facilitated by human activity during Guassa harvest giving an opportunity to widely disperse in the soil [65].
Different measurements taken during data collection were also highly impacts the recovery of vegetation species and determines the abundance of mature vegetation and seeding stage of the existing shrub and grass species in the study area. Disturbances such as erosion, seed dispersal agents, harvesting time, encroachment and climate change had great impact on the recovery of grass species. Also the anthropogenic activates during harvesting had a great impact on the recovery of the spices in the study area. This result is in agreement with the report done by Kidane [41]. All the regeneration of vegetation in the study site is highly impacted by both climatic and human factors [66]. According to the data reported by Girma [67], the recovery and rehabilitation rate of vegetation is primarily influenced by topographical variation, site location, rate of grazing intensity, human interference and combination of all the determinant factors.
The Sorenson’s similarity between the soil seed bank and above ground vegetation was relatively high (0.6). Thirty plant species (23.3 %) were represented both in the above ground vegetation and soil seed banks. From these plant species, Haplocarpha rueppellii was found in the above ground flora were not represented in the seed bank. The regional climate has a paramount role in similarities for species distribution patterns in afroalpine ecosystems, especially in those with highly diverse topography [55]. The higher similarity observed were because both share similar climate and altitude interval. The ecological distance between any two areas has also been noted to play an important part for differences in species composition [68, 69, 70, 71, 72].
Conclusion
In this study composition, density, abundance and similarity were determined for the vegetation in GCCA. The result shows that Asteraceae was found to be the most dominant family followed by Fabaceae and Poaceae. The results of this study is important in understanding the regeneration status of plant species in GCCA, which is generously significant for the sustainable utilization, management and decision making for the future conservation of the Guassa conservation area. The findings of the study indicated that abundance of the two target species (H. splendidum and F. macrophylla) was significantly affected by altitude and aspect. Species abundance decreases as altitude increase. Differences in these elevational patterns of plant abundance may be due to management practices, spatial heterogeneity (topographic diversity) and/or microclimate. The high elevation, Afroalpine grassland, was found to host higher endemic and rare plants while harboring lower species richness. Aspect also significantly affects species abundance. High number of F. macrophylla and H. splendidum recorded along North-West and North-East direction followed by South-West and South-East directions. The availability of solar light increases air and soil temperature on the north facing slope leads to high growth of herbaceous plants. Measurements/treatments done during the study also highly impacts the recovery of vegetation species and determines the abundance of mature vegetation and seeding stage of the existing plant species in the study area. The high similarity for species distribution pattern between seed bank and aboveground vegetation was due to similar climate and altitude interval. Finally, we recommend that the management intensity of the GCCA should receive increased attention from the responsible body. Especially the local government (Guassa Administration) and the community should collaborate with NGO and other organizations working on Guassa area for the better management and sustainable use of this conservation area.
Supplementary materials
All data generated and analyzed during this study are included in this work.
Acknowledgements
We are sincerely thankful to the Department of Plant Biology and Biodiversity Management, Addis Ababa University, Ethiopia, for supporting this work. We thank the communities of Guassa Community Conservation Area for their assistance and permission during field data collection.
References
-
Rahman M, Khan M, Arfin AS, Roy B, Fardusi M (2011) Assessment of natural regeneration status and diversity of tree species in the biodiversity conservation areas of Northeastern Bangladesh. Journal of Forestry Research 22(4): 551-559.
-
Mishra Bp, Tripathi OP, Laloo RC (2005) Community characteristics of a climax subtropical humid forest of Meghalaya and population structure of ten important tree species. Tropical Ecology 46(2): 241-252.
-
Pala NA, Negi AK, Gokhale Y, Bhat JA, Todaria NP (2012) Diversity and regeneration status of Sarkot Van Panchyat in Garhwal Himalaya, India. Journal of Forestry Research 23(3): 399-404.
-
Singh S, Malik ZA, Sharma CM (2016) Tree species richness, diversity, and regeneration status in different oak (Quercus spp.) dominated forests of Garhwal Himalaya, India. Journal of Asia-Pacific Biodiversity 9(3): 293-300.
-
Mishra BP, Tripathi OP, Tripathi RS, Pandey HN (2004) Effects of anthropogenic disturbance on plant diversity and community structure of a sacred grove in Meghalaya, northeast India. Biodiversity & Conservation 13(2): 421- 436.
-
Endress BA, Chinea JD (2001) Landscape Patterns of Tropical Forest Recovery in the Republic of Palau 1. Biotropica 33(4): 555-565.
-
Bai Y, Broersma K, Thompson D, Ross TJ (2004) Landscape-level dynamics of grassland-forest transitions in British Columbia. Journal of Range Management 57(1): 66-75.
-
Dawes WR, Short D (1994) The significance of topology for modeling the surface hydrology of fluvial landscapes. Water Resources Research 30(4): 1045-1055.
-
Titshall LW, O’Connor TG, Morris CD (2000) Effect of long-term exclusion of fire and herbivory on the soils and vegetation of sour grassland. African Journal of Range and Forage Science 17(1-3): 70-80.
-
Busing RT, White PS, MacKenzie MD (1993) Gradient analysis of old spruce–fir forests of the Great Smoky Mountains circa 1935. Canadian Journal of Botany 71(7): 951-958.
-
Haque SMS, Alam MS (1988) Some aspects of practicing the clear felling followed by artificial regeneration system in the Cox’s Bazar Forest Division. Chittagong University Studies, Part II: Science 12(2): 87-95.
-
Hossain MK, Azad AK, Alam MK (1999) Assesment of Natural Regeneration Status in a Mixed Tropical Forest at Kaptai of Chittagong Hill Tracts (South) Forest Division. Chittagong University Journal Of Science 23(1): 73-80.
-
Dutta G, Devi A (2013) Plant diversity, population structure, and regeneration status in disturbed tropical forests in Assam, northeast India. Journal of Forestry Research 24(4): 715-720.
-
Tesfaye G, Teketay D, Fetene M, Beck E (2010) Regeneration of seven indigenous tree species in a dry Afromontane forest, southern Ethiopia. Flora- Morphology, Distribution, Functional Ecology of Plants 205(2): 135-143.
-
Tyagi JV, Qazi N, Rai SP, Singh MP (2013) Analysis of soil moisture variation by forest cover structure in lower western Himalayas, India. Journal of forestry research 24(2): 317-324.
-
Level FWPA (2010) Tree growth relationships and silvicultural tools to assist stand management in private native spotted gum dominant forests in Queensland and northern New South Wales.
-
Saxena AK, Singh SP, Singh JS (1984) Population structure of forests of Kumaun Himalaya: implications for management. Journal of Environmental Management 19(4): 307-324.
-
Kadavul K, Parthasarathy N (1999) Plant biodiversity and conservation of tropical semi-evergreen forest in the Shervarayan hills of Eastern Ghats, India. Biodiversity & Conservation 8(3): 419-437.
-
Ashenafi ZT (2001) Common property resource management of an Afro-alpine habitat supporting a population of the critically endangered Ethiopian Wolf (Canis simensis). _PhD thesis, University of Kent_.
-
Ashenafi ZT, Leader-Williams N (2005) Indigenous common property resource management in the Central Highlands of Ethiopia. Human Ecology, 33(4): 539-563.
-
Ashenafi ZT, Leader-Williams N, Coulson T (2012) Consequences of human land use for an Afro-alpine ecological community in Ethiopia. Conservation and Society 10(3): 209-216.
-
Amessie G (2017) Opportunities and Challenges of Community Based Natural Resource Management in Menz-Guassa Community Conservation Area, Ethiopia.
-
Steger C, Boone RB, Dullo BW, Evangelista P, Alemu S, Gebrehiwot K, Klein JA (2022) Collaborative agent- based modeling for managing shrub encroachment in an Afroalpine grassland. Journal of Environmental Management 316: 115040.
-
Steger C, Gebrehiwot K, Chengere SA, Marinkovich J, Dullo BW, Zewde SW, Klein JA (2021) Mental models of a social-ecological system facilitate social learning among a diverse management team. Environmental Science & Policy 122: 127-138.
-
Steger C, Nigussie G, Alonzo M, Warkineh B, Van Den Hoek J, Fekadu M, Klein JA (2020) Knowledge coproduction improves understanding of environmental change in the Ethiopian highlands. Ecology and Society 25(2): 2.
-
Senbeta F, Schmitt C, Woldemariam T, Boehmer Hj, Denich M (2014) Plant diversity, vegetation structure and relationship between plant communities and environmental variables in the Afromontane Forests of Ethiopia. SINET: Ethiopian Journal of Science 37(2): 113-130.
-
Hendickx H, Jacob M, Frankl A, Guyassa E, Nyssen J, et al. (2014) Quaternary glacial and periglacial processes in the Ethiopian Highlands in relation to the current afro- alpine vegetation. Zeitschrift für Geomorphol 59(1):37- 57.
-
Rosqvist G (1990) Quaternary glaciations in Africa. Quaternary Science Reviews 9(2-3): 281-297.
-
Brooks TM, Mittermeier RA and Mittermeier CG (2002) Habitat Loss and Extinction in the Hotspots of Biodiversity. Conservation Biology 16(4): 909-923.
-
Hoffman M, Koenig K, Bunting G, Costanza J, Williams KJ (2016) Biodiversity hotspots (version 2016.1). Zenodo.
-
Moges E (2015) Population Structure, Behavioural Ecology and Habitat Vulnerability of Gelada (Theropithecus gelada) in Guassa Community Protected Area, Ethiopia.
-
Simeneh G (2010) Habitat use and diet of golden jackal (Canis aureus) and human-carnivore conflict in Guassa community conservation area, Menz. Addis Ababa, Ethiopia: Addis Ababa University.
-
Beyene H (2010) Population Estimate and Structure of the Gelada Baboon, Theropithecus Gelada, in the Guassa Community Conservation Area, Central Ethiopia.
-
Mueller-Dombois D, Ellenberg H (1974) Aims and methods of vegetation ecology. John Wiley, New York.
-
Sorensen T (1948) A method of establishing groups of equal amplitude in plant sociology based on similarity of species and its application to analyses of the vegetation on Danish commons. Kongelige Danske Videnskabernes Selskab 5 (4): 1-34.
-
Kent M, Coker P(1992) Vegetation Description and Analysis. A practical approach. John Wiley and Sons, New York 363-448.
-
Tadesse, M (2004). Asteraceae (Compositae) In: Hedberg I, Friis I, Edwards S, (eds). Flora of Ethiopia and Eritrea. (vol. 4:2). Department of Systematic Botany, Uppsala, Sweden & The National Herbarium, Addis Ababa, Ethiopia. pp. 408.
-
Hedberg O (1964) Features of Afroalpine plant ecology_._ Acta Phytogeographica Suecica 49: 1-144.
-
Yineger H, Kelbessa E, Bekele T, Lulekal E (2008) Floristic composition and structure of the dry afromontane forest at Bale Mountains National Park, Ethiopia. SINET: Ethiopian Journal of Science 31(2): 103-120.
-
Gurmessa F, Soromessa T, Kelbessa E (2013) Floristic Composition and Community Analysis of Komto Afromontane Moist Forest, East Wollega Zone, West Ethiopia. Science, Technology and Arts Research Journal 2(2): 58-69.
-
Kidane L, Nemomissa S, Woldu Z (2016) The effects of disturbance on the population structure and regeneration potential of five dominant woody species– in Hugumburda‐Gratkhassu National Forest Priority Area, North‐eastern Ethiopia. African journal of ecology 54(1): 20-28.
-
Whittaker RJ, Willis KJ, Field R (2003) Climatic-energetic explanations of diversity. In Macroecology: Concepts and Consequences: 43rd Symposium of the British Ecological Society, Cambridge University Press. 43: 107.
-
Melese GT, Tsegay BA, Kassa GM (2017). Effects of environmental variables on the patterns of plant community distribution in the afro-alpine vegetation of simien mountains national park, ethiopia. Journal of Biotechnology International 10(1): 8-21.
-
Bruun HH, Moen J, Virtanen R, Grytnes JA, Oksanen L, et al. (2006) Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities. Journal of Vegetation Science, 17(1): 37-46.
-
Korner C (2012) Alpine treelines: functional ecology of the global high elevation tree limits. Springer Science & Business Media.
-
Gairola S, Ghildiyal SK, Sharma CM, Suyal S (2009) Species richness and diversity along an altitudinal gradient in moist temperate forest of Garhwal Himalaya. Am J Sci 5 119-128.
-
Otypkova Z, Chytry M, Tichy L, Pechanec V, Jongepier JW, Hajek O (2011) Floristic diversity patterns in the White Carpathians biosphere reserve, Czech Republic. Biologia 66(2): 266-274.
-
Simegn TY, Soromessa T (2015) Carbon stock variations along altitudinal and slope gradient in the forest belt of Simen Mountains National Park, Ethiopia. American Journal of Environmental Protection 4(4) 199-201.
-
Jin XM, Zhang YK, Schaepman M, Clevers JGPW, Su Z (2008) Impact of elevation and aspect on the spatial distribution of vegetation in the Qilian Mountain area with remote sensing data. International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences-ISPRS Archives, Part B7 37, pp: 1385-1390.
-
Khalik KA, El-Sheikh M, El-Aidarous A (2013) Floristic diversity and vegetation analysis of wadi Al-Noman, Mecca, Saudi Arabia. Turkish Journal of Botany 37(5): 894-907.
-
Zhang W, Huang D, Wang R, Liu J, Du N (2016) Altitudinal patterns of species diversity and phylogenetic diversity across temperate mountain forests of northern China. PloS one 11(7): e0159995.
-
Tsujino R, Yumoto T (2013) Vascular plant species richness along environmental gradients in a cool temperate to sub-alpine mountainous zone in central Japan. Journal of plant research 126(2): 203-214.
-
Hill JL, Curran PJ (2003) Area, shape and isolation of tropical forest fragments: effects on tree species diversity and implications for conservation. Journal of biogeography 30(9): 1391-1403.
-
Bekele T (1993) Vegetation ecology of remnant Afromontane forests on the central plateau of Shewa, Ethiopia. Sv vaxtgeografiska sallsk.
-
Schmitt CB, Senbeta F, Woldemariam T, Rudner M, Denich M (2013) Importance of regional climates for plant species distribution patterns in moist Afromontane forest. Journal of Vegetation Science 24(3): 553-568.
-
Økland RH (1990) Vegetation ecology: theory, methods and applications with reference to Fennoscandia. Sommerfeltia, 1(s1): 1-238.
-
Dugdale JS (1964) Ethiopian climates and vegetation: the state of our present knowledge. Journal of Semitic Studies 9(1): 250-256.
-
Lamb H, Darbyshire I, Verschuren D (2003) Vegetation response to rainfall variation and human impact in central Kenya during the past 1100 years. The Holocene 13(2): 285-292.
-
Darbyshire I, Lamb H, Umer M (2003) Forest clearance and regrowth in northern Ethiopia during the last 3000 years. The Holocene 13(4): 537-546.
-
Maren IE, Karki S, Prajapati C, Yadav RK, Shrestha BB (2015) Facing north or south: Does slope aspect impact forest stand characteristics and soil properties in a semiarid trans-Himalayan valley?. Journal of arid environments 121: 112-123.
-
Gillingham P (2010) The relative importance of microclimate and land use to biodiversity (Doctoral dissertation, University of York).
-
Ghimire B, Mainali KP, Lekhak HD, Chaudhary RP, Ghimeray AK (2010) Regeneration of Pinus wallichiana AB Jackson in a trans-Himalayan dry valley of north- central Nepal. Himalayan Journal of Sciences 6(8): 19- 26.
-
Paudel S, Vetaas OR (2014) Effects of topography and land use on woody plant species composition and beta diversity in an arid Trans-Himalayan landscape, Nepal. Journal of Mountain Science 11(5): 1112-1122.
-
Nevo E (2001) Evolution of genome-phenome diversity under environmental stress. Proceedings of the National Academy of Sciences 98(11): 6233-6240.
-
Schimmel J, Granstrom A (1996) Fire severity and vegetation response in the boreal Swedish forest. Ecology 77(5): 1436-1450.
-
Kuma M, Shibru S (2015) Floristic Composition, Vegetation Structure, and Regeneration Status of Woody Plant Species of Oda Forest of Humbo Carbon Project, Wolaita, Ethiopia. Journal of Botany 2015.
-
Girma A (2012) Plant communities, species diversity, seedling bank and resprouting in Nandi forests, Kenya.
-
Mueller-Dombois D, Ellenberg H (1974) Aims and methods of vegetation ecology. John Wiley, New York.
-
Betemariam E A (2011) Forest diversity in fragmented landscapes of northern Ethiopia and implications for conservation.
-
Molla EL (2014) Plant Diversity and Ethnobotanical Study of Medicinal Plants in Ankober District, North Shewa Zone of Amhara Region, Ethiopia.
-
Woldemichael LK, Bekele T, Nemomissa S (2010) Vegetation composition in Hugumbirda-Gratkhassu national forest priority area, South Tigray. Momona Ethiopian Journal of Science 2(2): 27-48.
-
World Conservation Monitoring Centre (1994) Biodiversity Data Sourcebook. World Conservation Press, Cambridge, UK.
- Lessons to Learn: Trees are More than the Lungs of the World
- Community Forestry Enterprises as a Model for Sustainable Forest Development: The Case Of The "Baja Tarahumara" in Chihuahua, Mexico
- Ecological and Socio-Economic Impacts of Chromolaena odorata and Mesosphaerum suaveolens, Two Invasive Alien Species in Central and Southern Benin, West Africa
- Epigenetic Sustainability: Modeling the Human Factor as a Natural Resource through Science 4.0 and the NR3C1 Biological Pilot
- Growth-at-Risk: A Framework for Assessing Economic Vulnerability
- The Rural Territory as a Socioecological System for the Management of Public Policy for Sustainable Rural Development