HPSC-Based 2.5D In Vitro Model for Embryonic Morphogenesis Study
Introduction
Embryonic developmental processes see the formation of multiple cell types in predictable, polarised manners. One such developmental process is gastrulation, which firstly breaks the symmetry and establishes future body axes while defining the three germ layers-ectoderm, mesoderm, and endoderm- and followed by the later organogenesis that forms all the different cell types in our body [1]. As we cannot study human gastrulation in vivo, scientists have turned to other model organisms like the mouse and zebrafish. Through these studies, researchers have found that the processes during gastrulation are relatively well-conserved [2]. However, such model systems introduce significant inter-species variation [3]. The subsequent discovery of embryoid bodies (EBs) provided an alternative human cell- based substitute that cells could migrate and rearrange in a 3D fashion [4]. However, in vitro 3D models have their disadvantages, especially concerning imaging their deeper layers. Most recently, it was observed that hESCs on confined 2D substrates micropatterns could differentiate and self- organize to the three germ layers in a spatially defined manner that recapitulate the human gastrulation [5]. It is thus poised to be an attractive method of studying human embryonic gastrulation in vitro.
Several approaches have been addressing how the biochemical cues contribute to the germ layers differentiation on micropatterned hESCs. These approaches used different growth factor recipes to trigger the differentiation and all of which appear to produce similar results. These growth factors include Activin A, bone morphogenetic protein 4 (BMP4), fibroblast growth factor 2 (FGF2), or a combination thereof, with BMP4 being the constant and the crucial [5, 6, 7, 8, 9]. It is because BMP4 is known to work with FGF2 to induce mesoderm formation, while Activin A and FGF2 signaling initiate endoderm formation [10]. Moreover, BMP4 activates the Wnt-pathway and subsequently activates the Activin- Nodal singling in the mouse embryo, which makes Activin A probably dispensable [7, 11]. Yet, it is still unclear why micro patterned hESCs can self-organize into different germ layers despite being exposed to the same biochemical signals though some have proposed that differential diffusion rates of growth factors or biased distribution of receptors could play crucial roles [6, 7]. Nevertheless, how the signaling transduces from growth factors to the cell-fate determinants that ultimately instruct germ layers differentiation with a spatiotemporal precision is vastly elusive and can be further investigated.
Furthermore, in addition to the biochemical stimuli, the mechanical forces have been shown to play a significant role in polarity establishment during embryogenesis in worms [12]. As a result of the activation of the RHO-mediated actomyosin contraction, locally elevated cortical tension benefits the segregation of the polarity protein PAR3 by maintaining its clustering degree, whereas PKC3 shuffles between PAR3 and CDC42 pools and thus exhibits differential kinase activities that later modulate downstream cell-fate determinants. Hence, it’s interesting to further understand how the biochemical- and mechanical- cues cooperate to regulate gene expression and collective cell migration by which proper germ layers differentiation can be achieved during gastrulation using the in vitro 2.5D gastruloid model. Interestingly, there’s an E- to N-Cadherin transition from the center to the edge of the micropattern, suggesting an EMT-like property of the trophectoderm and endoderm populations within which the RHO is prominently enriched. Collectively, these results indicate the robustness of this in vitro 2.5D model that ideally recapitulates in vivo morphogenesis and polarity establishment in early embryogenesis.
In addition to serving as a tool to understand underlying mechanisms driving embryo development, the 2.5D gastruloid model is also suitable for teratogenicity testing for any given industrial, environmental, food-relevant, or pharmaceutical compounds due to its spatial consistency of the differentiated germ layers and easiness of preparation. Moreover, the small size (1mm diameter) and fast development time (3 days) required for the micropattern make it possible for a high-throughput and large-scale teratogen screening. As we come to better harness the hPSC-based in vitro embryo models in the future, wider applications that not only allow us to have a glimpse on how the amazing human embryos get mature in the womb but also aid to reduce the occurrence of developmental failures by understanding the underlying etiologies of the congenital disease progression by using the 2.5D gastruloids derived from disease-specific iPSC will be possible.
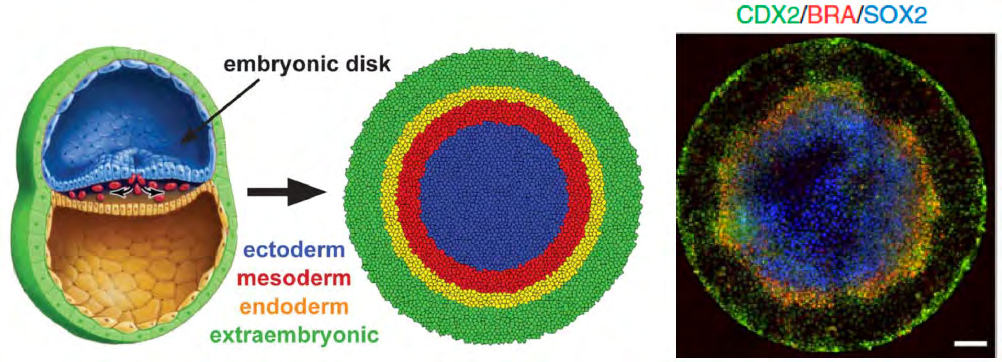
Figure 1: The 2.5D gastruloid model perfectly recapitulates the in vivo gastrulation by giving rise to the three fundamental germ layers from a homogeneous hESC micropattern. CDX2, BRA and SOX2 are specific markers for trophectoderm (extraembryonic), mesoderm and ectoderm, respectively. Endoderm marker was not included in the staining, so it’s shown as a blank. Figures are adapted from the works published by Heemskerk, et al. and Warmflash, et al.
References
-
Arnold SJ, Robertson EJ (2009) Making a commitment: cell lineage allocation and axis patterning in the early mouse embryo. Nat Rev Mol Cell Biol 10(2): 91-103.
-
Solnica Krezel L (2005) Conserved Patterns of Cell Movements during Vertebrate Gastrulation. Curr Biol 15(6): R213-R228.
-
Tesar PJ, Chenoweth JG, Brook FA, Davies TJ, Evans EP, et al. (2007) New cell lines from mouse epiblast share defining features with human embryonic stem cells. Nature 448(7150): 196-199.
-
Simunovic M, Metzger JJ, Etoc F, Yoney A, Ruzo A, et al. (2019) A 3D model of a human epiblast reveals BMP4- driven symmetry breaking. Nat Cell Biol 21(7): 900-910.
-
Deglincerti A, Etoc F, Guerra MC, Martyn I, Metzger J, et al. (2016) Self-organization of human embryonic stem cells on micropatterns. Nat Protoc 11(11): 2223-2232.
-
Etoc F, Metzger J, Ruzo A, Kirst C, Yoney A, et al. (2016) A Balance between Secreted Inhibitors and Edge Sensing Controls Gastruloid Self-Organization. Dev Cell 39(3): 302-315.
-
Martyn I, Brivanlou AH, Siggia ED (2019) A wave of WNT signaling balanced by secreted inhibitors controls primitive streak formation in micropattern colonies of human embryonic stem cells. Development 146(6): 172791.
-
Warmflash A, Sorre B, Etoc F, Siggia ED, Brivanlou AH, et al. (2014) A method to recapitulate early embryonic spatial patterning in human embryonic stem cells. Nat Methods 11(8): 847-854.
-
Xing J, Toh YC, Xu S, Yu H (2015) A method for human teratogen detection by geometrically confined cell differentiation and migration. Sci Rep 5(1): 10038.
-
Bernardo AS, Faial T, Gardner L, Niakan KK, Ortmann D, et al. (2011) BRACHYURY and CDX2 Mediate BMP-Induced Differentiation of Human and Mouse Pluripotent Stem Cells into Embryonic and Extraembryonic Lineages. Cell Stem Cell 9(2): 144-155.
-
Ben Haim N, Lu C, Guzman Ayala M, Pescatore L, Mesnard D, et al. (2006) The Nodal Precursor Acting via Activin Receptors Induces Mesoderm by Maintaining a Source of Its Convertases and BMP4. Dev Cell 11(3): 313-323.
-
Wang SC, Low TYF, Nishimura Y, Gole L, Yu W, et al. (2017) Cortical forces and CDC-42 control clustering of PAR proteins for Caenorhabditis elegans embryonic polarization. Nat Cell Biol 19(8): 988-995.
- Research Progress of Induced Pluripotent Stem Cells and Their Clinical Application Prospects
- Nishan Al-Kamal is the Starting Point of A Feminist Scientist
- Current Concepts and Future Perspectives of Stem Cell Therapy in Peripheral Arterial Disease
- Stem Cell and Oxidative Stress-Inflammation Cycle
- Adipose Derived Mesenchymal Stem Cells Origin, Characteristics and Promises
- Mitochondria Targeted Antioxidants can Improve In Vitro Embryo Production in Buffalo