Polycomb Group Proteins and Lineage Specification
Lineage specification is a crucial step during development of multicellular organism that requires differential gene expression. Epigenetic modifiers such as chromatin remodelers, histone modifiers, non-coding RNAs, and DNA methylases enable differential gene expression. Polycomb group (PcG) proteins are one of the important histone modifier which are involved in development as well as disease. PcG proteins assemble into large protein complexes termed Polycomb Repressive Complexes (PRCs), which introduce histone modifications that leads to gene repression. The PcG proteins and the Polycomb repressive complexes have been extensively studied in fruit flies and mice, and these studies show that PcG protein are crucial determinants of lineage specification. However, for obvious ethical reasons, the activity of early lineage specification in early human development has not been fully explored. This review attempts to update the readers about the role of Polycomb Group (PcG) proteins during lineage specification from studies on human pluripotent and human adult stem cells.
Introduction
Multicellular organisms arise from a single totipotent cell, which undergoes multiple rounds of cell division. Large group of these new cells specialize for specific functions, these later form tissues. Epigenetics permits differential gene expression that enables cells with same genetic material to form specialized cells with distinct gene expression profile. DNA methyltransferases, chromatin remodellers and histone modifiers are some of the key epigenetic mechanisms used during lineage specification. The histone modifications such as H3K27me3/2, H3K9me3, H2AK119ub1 lead to gene repression while H3K4me3, H3K36me3 activate gene expression [1, 2, 3, 4, 5, 6]. Polycomb group (PcG) proteins include large number of proteins that assemble into large complexes – Polycomb Repressive Complexes (PRCs) which catalyze histone modifications [7, 8, 9]. It has been shown in mice that loss of PcG proteins such as Ezh2, Suz12, Eed, Pcgf6 and Ring1b causes death of the developing mice embryo; while loss of others such as Bmi1, Cbx2, cbx7 and Rybp lead to developmental abnormalities [10, 11, 12, 13, 14, 15]. Surprisingly, when inner cell mass (ICM) was taken from mice embryos lacking key PcG proteins they showed the characteristics pluripotency features but were unable to differentiate in vitro. These studies unequivocally showed that PcG protein are critical for lineage specification. Polycomb group (PcG) proteins have been extensively studied in lower animals such as C.elegans and D.melanogaster [16, 17, 18, 19], however, their role in mammalian development is being increasingly explored. It is ethically and technically challenging to study role of PcG proteins during human embryonic development. Human pluripotent stem cells can be used to investigate role of PcG proteins in lineage specification in vitro. This review gives readers a glimpse into how Polycomb group proteins regulate expression of specific genes required for lineage specification based on the studies carried out with human pluripotent stem cells.
Polycomb group proteins in human pluripotent stem cells
RING1B is the main catalytic protein of the PRC1 and has been shown to occupy promoters of neuronal specific genes during neuronal differentiation of human pluripotent stem cells, while some specific neuronal genes were shown to be regulated by EZH2 [20]. Inhibition of RING1B using a pharmaceutical inhibitor PRT4165 led to increased expression of neuronal genes such as PAX6 and NESTIN [21]. EZH2, the catalytic component of PRC2 complex when inhibited led to reduced H3K27me3 mark; this favors mesoderm differentiation of human embryonic stem cells [22]. Using CRISPR-Cas9 system knockout of EZH2, EED and SUZ12 were created in human embryonic stem cells (hESCs); these cells lacking EZH2, EED and SUZ12 spontaneously differentiated into meso-endoderm lineage, but ectoderm differentiation was not seen [23]. Human embryonic stem cells when differentiated into ectoderm, mesoderm and endomderm, showed a distict expression profile of Polycomb group (PcG) proteins such as RING1B, CBX2, BMI1, EZH2, EED and SUZ12. [24]. Although, it seems certain PcG proteins are associated with a specific lineage, however it is unlikely that Polycomb group (PcG) proteins alone can bring about lineage specification. The Polycomb repressive complexes (PRCs) made up of various combinations of PcG proteins must be guided by upstream signaling molecules and lineage specific transcription factors.
Polycomb Group Proteins in Human Adult Stem Cells
Human adipose derived mesenchymal stem cells (hASCs) when cultured in osteo-inductive media showed up regulation of Polycomb group (PcG) protein BMI1. Conversely, BMI1 knockdown reduced the ability of hASCs to differentiate into osteogenic lineage. Curiously, the hASCs showed changes in BMI1 protein but not in BMI1 mRNA [25], which is most likely due to analytical methods used. Human neural stem cells are difficult to isolate and expand in culture. shRNA mediated knockdown of BMI1 in neural stem cells led to reduction in their proliferation and self-renewal, however when BMI1 was forcibly overexpressed in the neural stem cells, these cells showed enhanced self-renewal and proliferation. BMI1 was found to regulate the expression of cell cycle inhibitor INK4A/ARF genes, thus when BMI1 was overexpressed it led to reduced INK4A/ARF and consequently enhanced neural stem cell proliferation [26]. BMI1 knockdown in human umbilical cord derived mesenchymal stem cells (hUC-MSCs) reduced their proliferative ability under hypoxia conditions as well as their immunomodulatory properties, hUC-MSCs normally showed increased proliferation under hypoxia. Since, mesenchymal stem cells are used clinically for their immunomodulatory properties; BMI1 overexpression in these cells might have yet untested clinical significance.
EZH2 inhibition by pharmaceutical inhibitor in pancreatic explants led enhanced production of endocrine cells in vitro [3]. Patients suffering from diabetes type I, can be injected human pancreatic islets, however, this is limited due to good quality endocrine cells from cadavers. The endocrine cells from the cadaveric pancreatic tissue could be increased by treating them with EZH2 inhibitor. Addition of EZH2 inhibitor to human mesenchymal stem cells in vitro led to increased osteogenesis but decreased adipogenesis, this was due to expression of RUNX2 (osteogenic master transcription factor) which under normal conditions is repressed by EZH2 [28]. Human bone marrow derived mesenchymal stem cells when treated with EZH2 inhibitor – GSK126 and BMP2 led to enhanced ostogenic differentiation in a 3D scaffold [29]. Polycomb group (PcG) proteins BMI1 and EZH2 are involved in regulating adult stem cell proliferation as well as differentiation. Thus, manipulating the PcG proteins in adult stem cells using either shRNA approach or via pharmaceutical inhibitors has great clinical potential and this should be explored in preclinical models.
Future Perspectives
Epigenetic mechanisms play a major role in lineage specification and histone modifications can fine tune gene expression. Several of the Polycomb group (PcG) proteins regulate cell proliferation, hence a lot of work on PcG proteins is being done in cancer biology. Recently, the use of small molecule inhibitors for PcG proteins such as EZH2 and BMI are in various clinical trial stages for treatment of various kinds of cancers (https://www.clinicaltrials.gov/). Since, PcG proteins regulate lineage specification; the small molecule inhibitors such as DZNep, PRT4164, GSK126 could be explored to achieve differentiation of human pluripotent stem cells into desired cell type. Although human pluripotent stem cells can be used to understand role of PcG protein in early lineage specification, but due to inherent limitation of 2D culture it has been difficult to explore their role that would be seen physiologically. Organoids generated from human pluripotent stem cells have shown to mimic in vivo development, and the role of Polycomb group (PcG) protein in lineage specification in humans could be explored using organoids. It would be essential to understand the proteins that regulate various PcG proteins by post translational modifications that regulate their activity. Polycomb group (PcG) proteins are recruited to specific locations on the chromatin, hence the upstream signals that recruit PcGs to specific gene promoters should be investigated and this would help in furthering our understanding the role of Polycomb group (PcG) proteins in lineage specification.
The PcG localization at specific gene promoters is retained through several rounds of mitosis during differentiation, but the mechanisms of PcG “memory” needs to be explored further. Understanding how Polycomb group proteins regulate lineage specification may have great implication in cell and developmental biology as well as from clinical perspective. Image created with BioRender.com
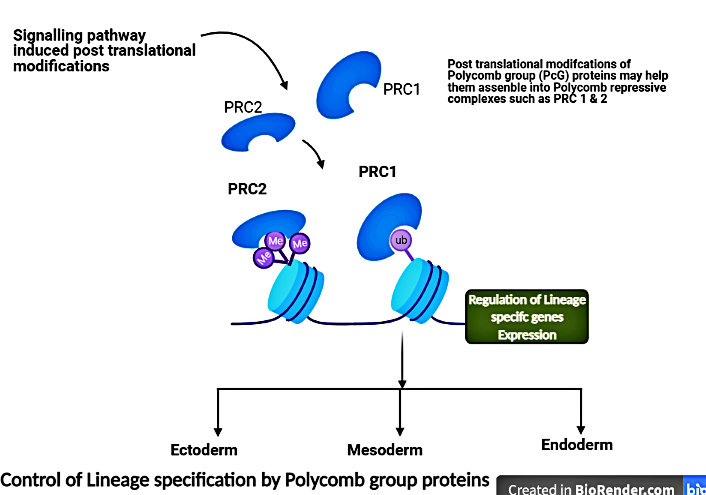
Figure 1: The schematic shows that signalling pathways may lead to post translational modifications of Polycomb group (PcG) proteins, these modifications may help assemble into large multimeric protein complexes - Polycomb repressive complexes (well known are PRC1 and PRC2) at specific gene. The PRCs catalyze histone modifications that regulate lineage specific genes, this leads to gene expression characteristic of germ lineages such as ectoderm, mesoderm and endoderm.
Acknowledgement
The author acknowledges support from Symbiosis Centre for Stem Cell Research (SCSCR), Symbiosis International University (SIU).
Conflict of Interest
Author declares no conflict of interest.
References
-
Zhang Y, Sun Z, Jia J, Du T, Zhang N, et al. (2021) Overview of Histone Modification. Adv Exp Med Biol 1283: 1-16.
-
Völker Albert M, Bronkhorst A, Holdenrieder S, Imhof A (2020) Histone Modifications in Stem Cell Development and Their Clinical Implications. Stem Cell Reports 15(6): 1196-1205.
-
Xu R, Li C, Liu X, Gao S (2021) Insights into epigenetic patterns in mammalian early embryos. Protein Cell 12(1): 7-28.
-
Hu Q, Greene CS, Heller EA (2020) Specific histone modifications associate with alternative exon selection during mammalian development. Nucleic Acids Res 48(9): 4709-4724.
-
de Napoles M, Mermoud JE, Wakao R, Tang YA, Endoh M, et al. (2004) Polycomb group proteins Ring1A/B link ubiquitylation of histone H2A to heritable gene silencing and X inactivation. Dev Cell 7(5): 663-676.
-
Mikkelsen TS, Ku M, Jaffe DB, Issac B, Lieberman E, et al. (2007) Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 448(7153): 553-560.
-
Pirrotta V (1997) PcG complexes and chromatin silencing. Curr Opin Genet Dev 7(2): 249-258.
-
Chan HL, Morey L (2019) Emerging Roles for Polycomb- Group Proteins in Stem Cells and Cancer. Trends Biochem Sci 44(8): 688-700.
-
Healy E, Mucha M, Glancy E, Fitzpatrick DJ, Conway E, et al. (2019) PRC2.1 and PRC2.2 Synergize to Coordinate H3K27 Trimethylation. Mol Cell 76(3): 437-452.
-
Faust C, Schumacher A, Holdener B, Magnuson T (1995) The eed mutation disrupts anterior mesoderm production in mice. Development 121(2): 273-285.
-
Voncken JW, Roelen BA, Roefs M, de Vries S, Verhoeven E, et al. (2003) Rnf2 (Ring1b) deficiency causes gastrulation arrest and cell cycle inhibition. Proc Natl Acad Sci U S A 100(5): 2468-2473.
-
Coré N, Bel S, Gaunt SJ, Aurrand Lions M, Pearce J, et al. (1997) Altered cellular proliferation and mesoderm patterning in Polycomb-M33-deficient mice. Development 124(3): 721-729.
-
Bracken AP, Dietrich N, Pasini D, Hansen KH, Helin K, et al. (2006) Genome-wide mapping of Polycomb target genes unravels their roles in cell fate transitions. Genes Dev 20(9): 1123-1136.
-
Shen X, Liu Y, Hsu YJ, Fujiwara Y, Kim J, et al. (2008) EZH1 mediates methylation on histone H3 lysine 27 and complements EZH2 in maintaining stem cell identity and executing pluripotency. Mol Cell 32(4): 491-502.
-
van der Lugt NM, Alkema M, Berns A, Deschamps J(1996) The Polycomb-group homolog Bmi-1 is a regulator of murine Hox gene expression. Mech Dev 58(1-2): 153- 164.
-
Chetverina DA, Lomaev DV, Erokhin MM (2020) Polycomb and Trithorax Group Proteins: The Long Road from Mutations in Drosophila to Use in Medicine. Acta Naturae 12(4): 66-85.
-
Kassis JA, Kennison JA, Tamkun JW (2017) Polycomb and Trithorax Group Genes in Drosophila. Genetics 206(4): 1699-1725.
-
Ross JM, Zarkower D (2003) Polycomb group regulation of Hox gene expression in C. elegans. Dev Cell 4(6): 891- 901.
-
Déjardin J, Cavalli G (2005) Epigenetic inheritance of chromatin states mediated by Polycomb and trithorax group proteins in Drosophila. Prog Mol Subcell Biol 38: 31-63.
-
Desai D, Khanna A, Pethe P (2020) Inhibition of RING1B alters lineage specificity in human embryonic stem cells. Cell Biol Int 44(6): 1299-1311.
-
Desai D, Khanna A, Pethe P (2020) PRC1 catalytic unit RING1B regulates early neural differentiation of human pluripotent stem cells. Exp Cell Res 396(1): 112294.
-
Lee JY, Yu KR, Kim HS, Kang I, Kim JJ, et al. (2016) BMI1 inhibits senescence and enhances the immunomodulatory properties of human mesenchymal stem cells via the direct suppression of MKP-1/DUSP1. Aging (Albany NY) 8(8): 1670-89.
-
Shan Y, Liang Z, Xing Q, Zhang T, Wang B, et al. (2017) PRC2 specifies ectoderm lineages and maintains pluripotency in primed but not naïve ESCs. Nat Commun 8(1): 672.
-
Pethe P, Pursani V, Bhartiya D (2015) Lineage specific expression of Polycomb Group Proteins in human embryonic stem cells in vitro. Cell Biol Int. 39(5): 600- 610.
-
Becker M, Potapenko T, Niklaus A, Bieback K, Ho AD, et al. (2016) Polycomb Protein BMI1 Regulates Osteogenic Differentiation of Human Adipose Tissue-Derived Mesenchymal Stem Cells Downstream of GSK3. Stem Cells Dev 25(12): 922-933.
-
Wang Y, Guan Y, Wang F, Huang A, Wang S, et al. (2010) Bmi-1 regulates self-renewal, proliferation and senescence of human fetal neural stem cells in vitro. Neurosci Lett 476(2): 74-78.
-
Hemming S, Cakouros D, Isenmann S, Cooper L, Menicanin D, et al. (2014) EZH2 and KDM6A act as an epigenetic switch to regulate mesenchymal stem cell lineage specification. Stem Cells 32(3): 802-815.
-
Lui H, Samsonraj RM, Vaquette C, Denbeigh J, Kakar S, et al. (2021) Combination of BMP2 and EZH2 Inhibition to Stimulate Osteogenesis in a 3D Bone Reconstruction Model. Tissue Eng Part A
-
Cao R, Zhang Y (2004) SUZ12 is required for both the histone methyltransferase activity and the silencing function of the EED-EZH2 complex. Mol Cell 15(1): 57- 67.
- Research Progress of Induced Pluripotent Stem Cells and Their Clinical Application Prospects
- Nishan Al-Kamal is the Starting Point of A Feminist Scientist
- Current Concepts and Future Perspectives of Stem Cell Therapy in Peripheral Arterial Disease
- Stem Cell and Oxidative Stress-Inflammation Cycle
- Adipose Derived Mesenchymal Stem Cells Origin, Characteristics and Promises
- Mitochondria Targeted Antioxidants can Improve In Vitro Embryo Production in Buffalo