Broad Spectrum Antiviral Therapy can Supersede Vaccination Strategy for Combating COVID19 Pandemic
As of today seven coronaviruses were identified to infect humans, out of which only 4 of them belongs to beta family of coronavirus, like HCoV-HKU1, SARS-CoV-2, MERS-CoV and SARS-CoV. SARS family viruses were known to cause severe respiratory disease in humans. SARS-CoV-2 has recently considered as a pandemic with over 100 M confirmed cases and over 1.5 M death worldwide. Vaccines and antibodies appeared as record pace. However, generation of variants has already been demonstrated as an escape of virus from a particular vaccine therapy. Therefore, a broad-spectrum anti-CoV therapies should be emphasized.
Introduction
What is COVID-19
A new threat to human life, in recent days, is the outbreak of SARSCoV-2 human coronavirus at Wuhan City of China in late December 2019. COVID-19 is the severe lower tract infectious disease caused by SARS-CoV-2, a beta family of human respiratory coronavirus. COVID-19 also presents several other pathologies in different patients [1]. As of April 9, 2021, the world-wide SARS-CoV-2 infected cases are more than 134M and death number is 3M. In USA, there have been over 30 million confirmed cases with 554K deaths, and worldwide there have been more than 134 M cases with 2.9K fatalities [2].
The infection capabilities of SARS-CoV-2 are due to its increased affinity for the angiotensin-converting enzyme 2 (ACE2) receptor presents on the recipient body’s cell surface [3, 4]. We have recently written an article referring the zoonosis, susceptibility and different strategies to develop therapeutics [5].
Transmission
Route of person-to-person transmission — Direct person-to-person respiratory transmission is the primary means of transmission of SARS-CoV-2 [6]. Infection might also occur if a person’s hands are contaminated by droplets or by touching contaminated surfaces and then they touch their eyes, nose, or mouth, although contaminated surfaces are not thought to be a major route of transmission. SARS- CoV-2 also has been detected in non-respiratory specimens, including stool, blood, ocular secretions, and semen, but the role of these sites in transmission is uncertain [7, 8, 9, 10, 11, 12, 13]. In particular, several reports have described detection of SARS- CoV-2 RNA from stool specimens, even after viral RNA could no longer be detected from upper respiratory specimens, and replicative virus has been cultured from stool in rare cases [8, 10, 11, 14].
Period of greatest infectiousness
Infected individuals are more likely to be contagious in the earlier stages of illness, when viral RNA levels from upper respiratory specimens are the highest [15, 16, 17, 18, 19].
Immunity following infection
Evidence suggests that some of these responses are protective and generally last at least several months. However, it is unknown whether all infected patients mount a protective immune response and how long protective effects last beyond the first few months after infection.
Humoral immunity
Following infection with SARS-CoV-2, the majority of patients develop detectable serum antibodies to the receptor-binding domain of the viral spike protein and associated neutralizing activity [16, 17]. However, the magnitude of antibody response may be associated with severity of disease, and patients with mild infection may not mount detectable neutralizing antibodies [20, 21]. When neutralizing antibodies are elicited, they generally decline over several months after infection, although most studies suggest that neutralizing activity is maintained for up to six to eight months [22, 23, 24, 25]. These antibody responses are associated with protection from SARS-CoV-2 infection.
Cell-mediated immunity
Studies have also identified SARS-CoV-2-specific CD4 and CD8 T cell responses in patients who had recovered from COVID-19 and in individuals who had received an investigational COVID-19 vaccine, which suggest the potential for a durable T cell immune response [26, 27, 28].
Protective immune response after infection or vaccination in primate studies
Animal studies have suggested that the immune response to infection may offer some protection against reinfection, at least in the short term [29, 30, 31, 32]. Antibodies that neutralize SARS-CoV-2 and SARS-CoV-2-reactive CD4 T cells have been identified in some individuals without known exposure to SARS-CoV-2, and some of these appear to be cross-reactive with antigens from common cold coronaviruses [28, 33, 34, 35]. These pre-existing immune responses whether impact the risk or the severity of COVID-19 and whether they will influence COVID-19 vaccine responses remain unknown.
Risk of reinfection
Overall, the short-term risk of reinfection (eg, within the first few months after initial infection) appears low. Nevertheless, sporadic cases of reinfection have been documented [36, 37, 38, 39, 40, 41, 42]. The first infection elicited detectable neutralizing activity (evaluated using a reference strain- based neutralizing assay) that was undetectable at the time of the second infection [43].
Therapeutic Approaches
To find a proper therapeutics for SARS-CoV-2, it is better to know first the structural and functional biology of the said virus. Coronaviruses are enveloped positive-stranded RNA viruses. Full-genome sequencing and phylogenic analysis indicated that human Coronavirus including SARS-CoV-2 have largest genome among all known RNA viruses (≥27 Kb) [44, 45]. Its nonstructural gene product replicase is required for viral replication and modulation of the antiviral effects [46, 47, 48].
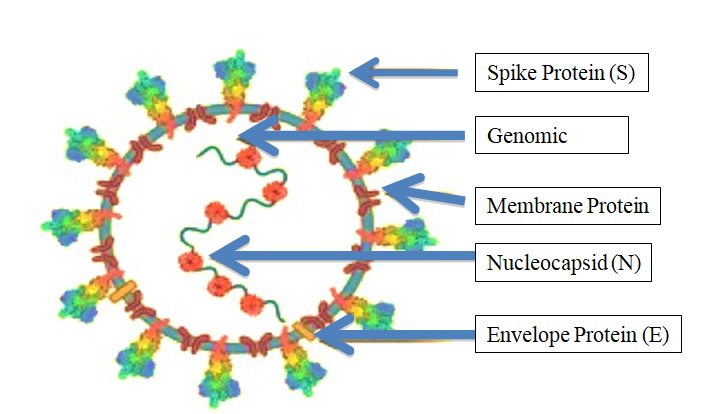
The structural gene region encodes spike protein (S), envelope (E), membrane (M), and nucleocapsid protein (N) [49, 50]. The spike (S) protein of SARS binds with the host cell surface angiotensin-converting enzyme 2 (ACE2), and then uses the host trans membrane serine protease-2 (TMPRSS2) for S priming for efficient fusion and cellular entry of SARS- CoV-2 [49, 51] (Figure 1). Mature Coronavirus as a double membrane compartment particles are COVID-19 causing virus, SARS-CoV-2 is similar to SARS-CoV but dissimilar in posing threat to humans transported to the cell [49].
Present Therapy
Over a year ago, when the very first SARS-CoV-2 cases in the USA were discovered, Nano Viricides was already working on developing a broad-spectrum anti-coronavirus drug. Yet the scientific community at large and regulatory efforts have remained focused on (a) vaccines, (b) antibodies, and (c) re- development of pre-existing drugs as antivirals [52].
A: Vaccine and/or Antibody
The strong government support led to rapid emergency approval, and later full approval, of an already known antiviral drug called remdesivir (Gilead). Strong fiscal support from the government also led to the emergency approval of two different antibody drugs, one from Regeneron, and one from Eli Lilly in the fastest ever drug development timeframe.
Even stronger commitments and strong government support led to the fastest ever emergency approval of two vaccines, both employing nanotechnology; both developed against the original 2019-nCoV-Wuhan variant, one by Pfizer-BioNTech, and one by Moderna.
Vaccine Technologies
mRNA Vaccine
mRNA vaccines combine desirable immunological properties with an outstanding safety profile and the unmet flexibility of genetic vaccines. Based on in situ protein expression, mRNA vaccines are capable of inducing a balanced immune response comprising both cellular and humoral immunity while not subject to MHC haplotype restriction. In addition, mRNA is an intrinsically safe vector as it is a minimal and only transient carrier of information that does not interact with the genome. Because any protein can be expressed from mRNA without the need to adjust the production process, mRNA vaccines also offer maximum flexibility with respect to development. Taken together, mRNA presents a promising vector that may well become the basis of a game-changing vaccine technology platform [53]. BioNtech and Pfizer already announced the final results of their COVID-19 vaccine phase 3 clinical trial on 18 November 2020 [54].
Only a couple of days earlier, Moderna had also revealed the preliminary outcome of their phase 3 study [55]. In both cases the claimed efficacy in preventing infection was 95% and 94.5%, respectively. Besides, BNT162b2, the vaccine developed by the small German start-up and the giant American pharma, and mRNA-1273 developed by the Cambridge-based biotech company in collaboration with the National Institutes of Health, are en-route to becoming the first prophylactic measures against SARS-CoV-2 infection.
Target Protein for vaccines
The virus has 15 non-structural proteins and 12 structural and accessory proteins. The spike (S) protein are responsible for recognition of the host cellular receptor to initiate virus entry. M proteins are embedded in the envelope and shape the virion envelope. E proteins are small polypeptides that are crucial for CoV infectivity. N proteins make up the helical nucleocapsid and bind structural rearrangement of the S protein and the insertion of FP required for virus– host membrane fusion [56, 57, 58, 59].
The spike protein (S)
Being composed of two subunits (S1 and S2), it mediates both virus-host attachment via its receptor-binding domain (RBD) and virus entry into the host cell [60]. Therefore, it could be the major target of neutralizing antibodies, which are most closely associated with protective responses against viral infection in humans.
M, E and N proteins
Unlike the S protein, CoV M and E proteins are poorly immunogenic for humoral responses, presumably owing to their small ecto-domains for immune cell recognition and small molecular sizes [61, 62] (Figure 1). Adoptive transfer of sera from donors immunized with a virus vector expressing M or E protein did not protect mice against SARS- CoV-2 infection [63]. Therefore, M and E proteins have never been explored as vaccine targets alone against SARS- CoV-2 or other CoVs.
Nonetheless, the sequence identity of M or E proteins among SARS- CoV, MERS- CoV and SARS- CoV-2 is much higher than for the S protein and RBD, suggesting the potential of M and E proteins as targets for cross- reactive T cells. Indeed, several T cell epitopes have been identified in M and E proteins in previous studies of SARS- CoV and MERS- CoV immunity [64]. In this regard, M and E proteins may help to broaden the T cell response and improve cross- protection if included in a SARS- CoV-2 vaccine.
The N protein
Is the most abundant viral protein and is highly immunogenic during CoV infections [65]. It is a major target for antibody responses and also contains T cell epitopes [66]. N- specific antibodies were reported to protect mice against mouse hepatitis virus, a mouse CoV, via Fc- mediated effector functions [67, 68]. However, anti- N immune sera did not protect against SARS- CoV-2 infection in a mouse model [63]. Immunization with N protein can also elicit CD4+ and CD8+ T cell responses in mice [69]. N- specific CD8+ T cell epitopes are known to protect chickens against IBV infection [70].
Venezuelan equine encephalitis virus replicon particles expressing an N- specific CD4+ T cell epitope showed complete protection against SARS-CoV infection [71]. However, its potential as a CoV vaccine target was largely undermined by early studies of SARS- CoV showing that vaccines expressing N protein did not provide protection and, on the contrary, enhanced infection-induced pneumonia via increased pulmonary eosinophil infiltration and TH2 cell-biased responses [72, 73], causing ERD. Therefore, the inclusion of N protein in CoV vaccines is complicated by balancing viral clearance and immune-pathogenesis and no N protein-based vaccine has been reported for COVID-19, so far.
The RBD
The RBD binds to the host receptor via a receptor- binding motif (RBM) on its external subdomain in SARS- CoV, MERS- CoV or SARS- CoV-2 [74, 75, 76]. The surface of the S protein is extensively shielded from antibody recognition by glycans, with the notable exception of the RBD, which explains the immune-dominance of RBD epitopes [77].
Most SARS- CoV-2 neutralizing antibodies (nAbs) bind to RBD and block the RBD–hACE2 interaction, thus inhibiting virus attachment [78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90]. RBD is an attractive vaccine target because it elicits high- quality, functionally relevant antibodies, while avoiding the potential risk of ADE, which is generally thought to be mediated by weak nAbs or non- nAbs [91, 92, 93].
The RBD also contains epitopes for T cell responses, as shown in studies of SARS- CoV, MERS- CoV and SARS- CoV- 2 [64, 94, 95, 96]. RBD- based antigens have been described in previous studies for SARS- CoV and MERS- CoV vaccine development [64, 94, 97, 98, 99]. To date, several RBD- based vaccines for COVID-19 have entered clinical trials. Yang, et al. reported an RBD- based COVID-19 vaccine candidate generated using a protein subunit strategy [95]. Consistent with this, a recent report of an RBD- based DNA vaccine also showed that the nAbs, and not the induced T cells, are immunogenic protects NHPs from COVID-19 [100]. An RBD- based mRNA vaccine is being developed in China and is currently in phase I trials.
This candidate vaccine, ARCoV, which expresses SARS- CoV RBD delivered by lipid nanoparticles, induced both nAb production and TH1 cell- biased responses in mouse and NHP models. Vaccination protected mice against challenge with a mouse- adapted SARS- CoV-2 strain [94]. However, the use of RBD in vaccines is compromised by its limited immunogenicity owing to its small molecular size and possible mixed forms of multiple complexes (as monomers, dimers or trimers). Strategies to overcome these drawbacks include increasing antigen size (for example, by fusing the RBD with an Fc domain), or by RBD multi-merization (for example, by displaying multiple copies of RBD on particles [101, 102, 103, 104, 105, 106, 107, 108]. Recently, to address these limitations, a dimeric form of the RBD of beta- CoV antigens suitable for use against SARS- CoV-2, MERS- CoV and SARS- CoV are in the process of development [109].
In addition to the RBD- dimer, an mRNA vaccine, BNT162b1 (BioNTech/Pfizer), was reported to express an RBD- trimer stabilized by the fold on trimerization domain (Figure 1). The phase I/II studies of this vaccine encouragingly showed that two doses of the vaccine induced nAbs to levels higher than those in convalescent patients as well as inducing TH1 cell- biased responses [110, 111].
Indeed, it is assumed that multivalent antigens would allow the crosslinking of B cell receptors for better B cell activation [112]. Interestingly, a recent study described a vaccine candidate comprising multiple copies of the SARS- CoV-2 RBD displayed in arrays on nanoparticles that induced a markedly lower binding to nAb ratio than a S-2P- based vaccine, indicating the humoral responses are focused on epitopes recognized by nAbs for the RBD- based vaccines, which are believed to have lower ADE potential [108]. S1- NTD and the S2 subunit. The S1- NTD also contains epitopes for CoV nAbs found in infected patients and has been considered a potential target in CoV vaccines [56, 57, 85, 113, 114, 115]. NTD- targeting nAbs generally do not directly block receptor binding but rather interfere with receptor binding or restrain the S protein conformational changes required for the pre- fusion to post- fusion transition [56, 114, 115]. It is notable that SARS- CoV-2 NTD- targeting nAbs generally exhibit lower neutralizing potency than RBD- specific nAbs [56]. We previously reported an NTD- based vaccine against MERS- CoV [116].
Vaccination with NTD protein elicited nAbs and NTD- specific T cell responses. Furthermore, it reduced lung abnormalities in a MERS-CoV challenge mouse model, although the immunogenicity and protective efficacy of the NTD protein were weaker than the RBD protein [116]. The inclusion of NTD in a COVID-19 vaccine would broaden the neutralizing epitopes and reduce the potential of viral escape of host immunity. Yet, so far, NTD- based vaccines against COVID-19 have not been reported.
S2 Subunit
Peptides derived from HR1 or HR2 of the S2 subunit from SARS-CoV, MERS- CoV and SARS- CoV-2 have been described that inhibit viral fusion with target cells and thereby prevent virus infection. Moreover, nAbs have been reported to target the S2 subunit of CoVs, including SARS- CoV-2 suggesting the S2 subunit as a COVID-19 vaccine target [39, 56, 117, 118]. However, the membrane- proximal S2 subunit contains more extensive N- glycan shielding and is less accessible for immune recognition than the S1 subunit and is therefore less immunogenic [119, 120]. Rabbits immunized with SARS- CoV-2 S2 protein showed much lower nAb titres than those immunized with the S1 subunit or RBD proteins [121]. S2 subunit- targeting antibodies isolated from convalescent patients showed weaker neutralizing activities against SARS- CoV-2 than RBD- targeting antibodies [86]. These studies suggest that the S2 subunit alone may not be an effective target for humoral responses. Nevertheless, because of the relative sequence conservation of the S2 subunit between virus species, the S2 subunit is targeted by cross- reactive antibodies and CD4+ T cells recognizing both SARS- CoV- 2 and other human CoVs, suggesting a potential target in universal CoV vaccines [86, 38, 39, 122].
Nanoparticle-based Vaccines
Two of the nanoparticles-based vaccines are close to obtaining approval by the US Food and Drug Administration could represent a giant step in the fight against the COVID-19 pandemic. Nanomedicine approaches, especially for cancer therapies, have often led to underwhelming results when translated from the pre-clinical to the clinical arena due to the complex and still poorly understood nature of the nano– bio interactions. The latest evidence suggests that in areas such as vaccine development the odds for a nanomedicine- based approach are more favorable [123, 124]. These strategies moreover are scalable and versatile, since mRNA can be engineered using standard laboratory techniques. This means that they can be easily and quickly adapted to produce new vaccines against future epidemics.
With respect to other approaches, mRNA-based therapies have several advantages. mRNA delivery is safer than whole virus or DNA delivery as mRNA is not infectious and cannot be integrated into the host genome; while DNA needs to reach the nucleus to be decoded, mRNA is processed directly in the cytosol; mRNA has a short half-life, which can be regulated by molecular design; finally, it is immunogenic, which might represent an advantage for vaccine design, yet its immunogenicity can be modulated with molecular engineering techniques [125]. However, to be safely and efficiently transported in vivo without being degraded in the circulation, and to reach the cytosol across the cellular plasma membrane, mRNA needs a carrier. For many mRNA- based therapeutics, including BNT162b2 and mRNA-1273, the vehicles of choice are lipid nanoparticles (although other materials have also been used) [125]. Complexed with positively-charged lipids, mRNA is more stable and resistant to RNase-mediated degradation and forms self-assembled virus-sized particles that can be administered via different routes [126]. Once endocytosed, the lipid nanoparticles promote endosomal escape, and release their genetic cargo in the cytosol, where the mRNA is translated into antigenic proteins, kick-starting the immune system machinery into producing neutralizing antibodies. Both BNT162b2 and mRNA-1273 deliver mRNA encoding genetic variants of the SARS-CoV-2 spike protein that are more stable and immunogenic than the natural protein. A current drawback of these formulations is that their long-term storage requires low temperatures, posing logistical hurdles to their potential distribution and administration, in particular for regions of the global south [127, 128, 129].
Adenovirus Cassette
Adenoviral vectors are the new COVID-19 vaccine front- runners. As with all vaccines, the idea is to trick our body into thinking it’s been infected. Those self-made spike proteins would train our bodies to detect and terminate any real SARS- CoV-2 infections. The technique has been in development for more than 3 decades, but thanks to COVID-19, it is about to be put to the test like never before.
As soon as the genetic sequence of SARS-CoV-2 was posted online in January, three groups began independently working on adenoviral vector vaccines for COVID-19: CanSino Biologics, the University of Oxford, and Johnson & Johnson. All three teams are chock full of vaccine veterans, and their COVID-19 programs have garnered global attention for their scale and speed.
CanSino’s adenoviral vector vaccine made it into human trials in China in March ‘20. Later that month, the US government pledged more than $500 million to help J&J make up to 1 billion doses of its vaccine, which isn’t expected to enter clinical testing until September. Oxford University, meanwhile, is taking the most ambitious approach: in late April, it started a 1,100-person trial to prove its vaccine’s safety while looking for signs that it works. Its goal is to complete that study in just a month and then begin a Phase III trial of 5,000 people as soon as June. If successful, Oxford’s program would leapfrog all other COVID-19 vaccines in development.
And all genetic vaccines—DNA vaccines, mRNA vaccines, and adenoviral vector vaccines—mimic a natural viral infection by forcing our bodies to produce viral proteins inside our cells. That spurs the T cells of our immune system to attack these vaccinated cells, and in the process, they learn to seek and destroy cells infected with the real virus in the future. Traditional vaccines, made from weakened viruses or viral proteins, stimulate B cells to make antibodies against the virus. Those antibodies latch onto invading viruses and prevent them from entering our cells. The problem is that once the virus infiltrates our cells, the antibodies from a traditional vaccine are useless. It’s at that stage that T cells need to swoop in. Adenovirus vectors “are the best of all vaccines at inducing a T-cell response,” Wistar’s Ertl says.
The National Institutes of Health, which partly funded the trials, called a meeting to decide whether to proceed with trials of Ad5-based vaccines. In 2009, it decided to push forward with a modified version of a planned HIV vaccine trial so long as the participants didn’t have preexisting immunity to Ad5. Results from the 2,500-person study showed that the vaccine was safe, but it still didn’t work.
That study curbed enthusiasm for Ad5, but didn’t eliminate it altogether. CanSino, a Chinese company founded by former Sanofi vaccine developers, developed an Ad5-based vaccine for Ebola during the 2014 outbreak, and a Phase II study showed that the vaccine induced an antibody response 4 weeks after injection. Several smaller firms are also developing COVID-19 vaccines based on Ad5. One of them is ImmunityBio, which uses Ad5 vectors with additional gene deletions. CEO Patrick Soon-Shiong says the modification drastically curtails the body’s toxic immune responses to the virus and even allows the vector to be dosed multiple times. The firm has tested the vector in about 200 people in several small clinical trials, mostly for cancer.
Other companies, including Altimmune, Stabilitech BioPharma, and Vaxart, believe they can circumvent preexisting immunity to Ad5 in the bloodstream by administering their vaccines as nasal sprays or pills rather than injections. The experimental formulations could also be easier to manufacture, store, distribute, and use.
To date, three COVID-19 vaccine candidates based on adenovirus expressing the full- length S protein have entered phase III clinical trials. One, which is being developed in China, is based on human adenovirus type 5 (Ad5) [31, 130].
Ad-Chimp used by Oxford (A-Z) to COVID
In 2012, the Oxford group developed its own chimpanzee-derived vector, dubbed ChAdOx1, based on an adenovirus discovered in chimpanzee feces. The Oxford team went on to create the spin-off company Vaccitech in 2016 and has developed experimental vaccines for a number of diseases, including AIDS, malaria, tuberculosis, and Middle East respiratory syndrome, which is caused by the MERS coronavirus. A small safety study of that MERS vaccine was conducted in 2018. The results, published this April, showed that most of the 24 people in the trial still had T cells that targeted the MERS virus 12 months after a single injection of the vaccine. They also still had elevated levels of antibodies a year later. But only about half the people who got the highest dose of the vaccine had antibodies that neutralized the MERS virus in lab experiments. That MERS work allowed the Oxford team to move fast on a COVID-19 vaccine, which essentially swaps in the genetic instructions for the SARS-CoV-2 spike protein.
A single dose of ChAdOx1 nCoV-19, an investigational vaccine against SARS-CoV-2, has protected six rhesus macaques from pneumonia caused by the virus, according to National Institutes of Health scientists and University of Oxford collaborators. The findings are not yet peer-reviewed but are being shared to assist the public health response to COVID-19. Based on these data, a Phase 1 trial of the candidate vaccine began on April 23 in healthy volunteers in the United Kingdom.
Oxford University has entered into a partnership with UK-based global biopharmaceutical company AstraZeneca for the further development, large-scale manufacture and potential distribution of the vaccine [131]. The second, being developed in the UK, uses recombinant chimpanzee adenovirus, ChAdOx1 [132, 133].
Inactivated virus: Besides Pfizer’s and Moderna- developed COVID-19 vaccine, a third one was developed by China’s Sinovac Biotech company from inactivated SARS-CoV-2 to stimulate our bodies immune response. This vaccine is similar to the flu and chickenpox vaccines, and is currently in late-stage trials [134].
Live attenuated virus: Live attenuated virus (LAV) vaccines have been a reliable means of inducing effective long-term immunity against various specific viral pathogens, such as polio and measles. These are developed by passaging the pathogen under heterogeneous conditions. For instance, the LAV measles strain called AIK, used for vaccine production, was produced by passaging in sheep kidney cell lines and then in chicken embryo cells at 33°C. This allows it to replicate at 35°C but not at 37°C or 39°C. Such temperature- sensitive strains are suitable for LAV vaccines.
An end result is a weakened form of the virus that cannot replicate easily within human cells. This allows the virus to generate an immune response but not to establish a productive infection. The mutant enzymes may be temperature-dependent in their activity. Alternatively, the structure of the NSP may be altered by temperature, hindering complex formation with other NSPs and therefore preventing viral replication. A50-18 thus isolated as a novel live attenuated vaccine candidate for the prevention of COVID-19 because they less pathogenicity but retained the immunogenicity [135].
Validity of S protein as target: The SARS-CoV-2 S protein binds to the host cell receptor and induces virus–cell membrane fusion, which plays a vital role in the process of virus invasion. Moreover, the high affinity between the S protein and ACE2 increases the infectivity of SARS-CoV-2. Mammals including pangolins, pets (dogs and cats), and members of Cricetidae may be important for determining key residues for association with S from SARS-CoV and SARS-CoV-2 understanding of the structure and function of SARS-CoV-2 S will allow for additional information regarding invasion and pathogenesis of the virus, which will support the discovery of antiviral therapeutics and precision vaccine design [136].
Structural information will also assist in evaluating mutations of the SARS-CoV-2 S protein and will help in determining whether these residues have surface exposure and map to known antibody epitopes of S proteins from other coronaviruses. In addition, structural knowledge ensures that the proteins produced by constructs are homogeneous and participate in the prefusion conformation, which should maintain the most neutralization sensitive epitopes when used as a candidate vaccine or B-cell probe for isolating neutralizing human mAbs. Furthermore, atomic-level details will enable the design and screening of small molecules that inhibit fusion. Since SARS-CoV-2 and SARS-CoV RBD domains share 75% amino acid sequence identity, future work will be necessary to evaluate whether any of these Abs neutralize newly emerged coronavirus. Overall, interaction between the S protein of SARS-CoV-2 and ACE2 should be further studied to contribute elucidation of the mechanism of SARS- CoV-2 infection. Similarly, focusing on high expression of the S protein or its receptor binding region is also of great significance for the development of vaccines.
The S2 subunit of SARS-CoV-2 shows 88% sequence homology with the SARS-CoV S2 domain and is structurally conserved. Therefore, the development of antibodies targeting this functional motif may cross-bind and neutralize these two viruses and related CoVs. Antiviral peptides prevent SARS- CoV-2 membrane fusion and can potentially be used for the prevention and treatment of infection. It is worth mentioning that EK1C4, which targets the highly conserved HR1 domain of the S2 subunit, is expected to have therapeutic potential against SARS-CoV-2. More importantly, EK1C4 can be used as a nasal drop, which increases its medicinal properties, it possesses a high genetic barrier to resistance, and does not easily induce drug-resistant mutations. On the other hand, peptide fusion inhibitors may not be widely used clinically and have low bioavailability. Therefore, the development of oral small molecule fusion inhibitors is a major direction.
In the course of virus epidemics, the ability to adapt to external pressure is an important factor affecting the spread of the virus. Regarding the envelope S protein, recombination or mutation in the gene of its RBD can occur to promote transmission between different hosts and lead to a higher fatality rate [137]. Mutation of the aspartate (D) at position 614 to glycine (G614) results in a more pathogenic strain of SARS-CoV-2 which makes it more difficult to develop antibodies or vaccines that target non-conservative regions [138]. To effectively prevent disease, combinations of different mAbs that identify different epitopes on the SARS- CoV-2 S surface can be assessed to neutralize a wide range of isolates, including escape mutants [139].
Potential Issues in using S-protein as target
- Autoantibodies against ACE and ACE-like hormones are possible.
- It has been hypothesized (suggested) by some experts that ACE inhibitors could make COVID-19 worse. But because COVID-19 is a new disease, we do not know if they actually do.
- ACE inhibitors reportedly increase levels of ACE-2 which is the protein that the SARs-CoV-2 virus binds to, and long-term use may also suppress the immune response.
- However, an overactive renin-angiotensin system may be another reason people with high blood pressure are more likely to develop pulmonary complications from COVID-19.
- There is currently no evidence that ACE inhibitors do make COVID-19 worse. But it is something that needs to be investigated further, to help guide practice during this current COVID-19 pandemic and for any new coronaviruses in the future that use ACE-2 as an entry point as well.
- Do not stop taking your ACE inhibitor because this may increase your chance of having a heart attack or stroke. If you have any concerns, talk to your doctor.
Variants are really considered: SARS-CoV-2 Variants a new virus variant has one or more mutations that differentiate it from the wild-type or predominant virus variants already circulating among the general population. As expected, multiple variants of SARS-CoV-2 have been documented in the United States and globally throughout this pandemic. To inform local outbreak investigations and understand the national picture, scientists compare genetic differences among viruses to identify variants and how closely they are related to each other.
Several new variants, most notably
B.1.1.7: In the United Kingdom (UK), a variant of SARS-CoV-2 known as B.1.1.7 emerged., and has a transmission advantage of 0.4 to 0.7 points higher in reproduction number, meaning it’s reproduction numbers could vary from 1.4 to 1.8 [140].
This variant carries a large number of mutations and has since been detected around the world, including in the United States (US). This variant was first detected in the US at the end of December 2020. In January 2021, scientists from the UK reported early evidence that suggests the B.1.1.7 variant may be associated with an increased risk of death compared with other variants [141, 142]. More studies are needed to confirm this finding.
B.1.351: In South Africa, another variant of SARS-CoV-2 known as B.1.351 emerged independently of B.1.1.7. According to a non-peer-reviewed preprint article, this variant shares some mutations with B.1.1.7 [143]. Cases attributed to B.1.351 have been detected outside of South Africa, and this variant was first detected in the US at the end of January 2021. Preliminary evidence from non-peer- reviewed publications suggests that the Moderna mRNA- 1273 vaccine currently used in the US may be less effective against this variant but additional studies are needed [144].
P.1: In Brazil, a variant of SARS-CoV-2 known as P.1 emerged; it was first identified in January 2021 in travelers from Brazil who arrived in Japan. This variant was detected in the US at the end of January 2021 [145, 146].
The P.1 variant has 17 unique mutations, including three in the receptor binding domain of the spike protein (K417T, E484K, and N501Y), according to non-peer-reviewed preprint articles [147, 148]. There is evidence to suggest that some of the mutations in the P.1 variant may affect the ability of antibodies (from natural infection or vaccination) to recognize and neutralize the virus but additional studies are needed [149].
One specific mutation, called D614G, is shared by these three variants. It gives the variants the ability to spread more quickly than the predominant viruses, as described in a non-peer-reviewed preprint article [150, 151]. There also is epidemiologic evidence that variants with this specific mutation spread more quickly than viruses without the mutation [152].
This mutation was one of the first documented in the US in the initial stages of the pandemic, after having initially circulated in Europe [153].
D-1: Other target protein for COVID-19 vaccine: New research shows why the N protein might be a good target. The latest results from the phase 3 COVID-19 vaccines trials have been very positive. These have shown that vaccinating people with the gene for SARS-CoV-2 spike protein can induce excellent protective immunity.
The spike protein is the focus of most COVID-19 vaccines as it is the part of the virus that enables it to enter our cells. Virus replication only happens inside cells, so blocking entry prevents more virus being made. If a person has antibodies that can recognize the spike protein, this should stop the virus in its tracks.
The three most advanced vaccines (from Oxford/ AstraZeneca, Pfizer/BioNTech and Moderna) all work by getting our own cells to make copies of the virus spike protein. The Oxford vaccine achieves this by introducing the spike protein gene via a harmless adenovirus vector. The other two vaccines deliver the spike protein gene directly as mRNA wrapped in a nanoparticle. When our own cells make the spike protein, our immune response will recognize it as foreign and start making antibodies and T cells that specifically target it.
However, the SARS-CoV-2 virus is more complicated than just a spike protein. There are, in fact, four different proteins that form the overall structure of the virus particle: spike, envelope (E), membrane (M) and nucleocapsid (N). In a natural infection, our immune system recognizes all of these proteins to varying degrees.
Following SARS-CoV-2 infection, researchers have discovered that we actually make the most antibodies to the N protein – not the spike protein. This is the same for many different viruses that also have N proteins. But how N protein antibodies protect us from infection has been a long-standing mystery. This is because N protein is only found inside the virus particle, wrapped around the RNA. Therefore, N protein antibodies cannot block virus entry, will not be measured in neutralization assays that test for this in the lab, and so have largely been overlooked.
New mechanism discovered: A new mechanism for how N protein antibodies can protect against viral disease has emerged from the MRC Laboratory of Molecular Biology in Cambridge. An unusual antibody receptor called TRIM21 is only found inside the cell. They have shown that N protein antibodies that get inside cells are recognized by TRIM21, which then shreds the associated N protein. Tiny fragments of N protein are then displayed on the surface of infected cells. T cells recognize these fragments, identify cells as infected, then kill the cell and consequently any virus.
This suggests that vaccines that induce N protein antibodies, as well as spike antibodies, could be valuable, as they would stimulate another way by which our immune response can eliminate SARS-CoV-2.
Adding N protein to SARS-CoV-2 vaccines could also be useful because N protein is very similar between different coronaviruses – much more so than the spike protein. This
means it’s possible that a protective immune response against SARS-CoV-2 N protein could also offer some protection against other related coronaviruses, such as MERS.
Another potential benefit that may arise from including N protein in SARS-CoV-2 vaccines is due to the low mutation rates seen in the N protein sequence. Some changes to the sequence of SARS-CoV-2 have been reported over the course of this pandemic, with the most significant changes occurring in the spike protein. There is some concern that if the spike sequence alters too much, then new vaccines will be required. This could be similar to the current need for annual updating of influenza vaccines. However, as the N protein sequence is much more stable than the spike, vaccines that include a component targeting the N protein are likely to be effective for longer [154].
E: Are vaccines a good strategy for using in late stage of this pandemic?: The coronavirus disease 2019 (COVID-19) pandemic disrupted our lives and our healthcare systems and accounted for millions of illnesses across the globe ranging from mild to severe to deadly. COVID-19 vaccination is a critical tool in stopping this pandemic.
There are several COVID-19 vaccines that are in the late stages of development. Currently, two vaccines have been authorized for use by the U.S. Food and Drug Administration and recommended by the Centers for Disease Control and Prevention. These vaccines are designed to teach your body’s immune system to recognize and fight off the virus that causes COVID-19.
Both of these vaccines require two doses. The first shot starts building protection. A second shot a few weeks later is needed to get the most protection the vaccine has to offer. Neither of these vaccines will give you COVID-19 [155].
Why are COVID-19 vaccines important? COVID-19 has sickened and killed millions of people worldwide. COVID-19 vaccines have been tested in clinical trials to determine that they are highly effective in stimulating our bodies to develop immunity and protect us from the disease. Therefore, COVID-19 vaccines are a critical tool in stopping the pandemic, resuming normal life, and protecting ourselves and others from this disease.
- Psychological comfort
- May reduce severity of diseases even from a variants.
• However, increase of ADE cannot be neglected. In case of SARS-CoV-1, ADE was demonstrated in animal model.
• Vaccines are protective prior to going to exposure event. However, the virus is constantly mutated.
• Is frozen vaccine going to alter our normal life, may be questionable.
F: Compare plasma therapy and Antibody therapy: MABs repeatedly escaped. Several approved MABs are not very effective based on the amount of dosing and clinical results.
Demerits of Vaccine/Antibody therapy: However, as the vaccines are undergoing deployment, several new virus variants of tremendous concern have already emerged. Additional virus variants will continue to emerge at an even faster rate because of the widespread dissemination of the virus with many patient bodies serving as factories.
It is well known that viruses, particularly RNA viruses, mutate rapidly, and that such changes produce “variants” that escape from vaccines as well as from antibody drugs. SARS- CoV-2 has a repair mechanism that retains some fidelity during reproduction, and therefore it changes less rapidly than Influenza A viruses or HIV. Nevertheless, given the significant penetration of the virus into human population, and the very high viral loads achieved in severe cases, the virus has a tremendous ability to change. This important concern, voiced by several eminent scientists, has been overlooked until recently.
Meanwhile, SARS-CoV-2 has generated at least four important variants that have had significant impact. The very first one, namely D614G, replaced the original Wuhan strain completely and rapidly during the first wave of the pandemic itself. In the current second wave, we have seen emergence of the B.1.1.7 variant from UK, the N501Y-V.2 (also called B.1.1.7 variant) from South Africa, and the P.1 variant from Brazil. Each of them arose independently and in distinctly different geographic spaces, and yet there are many common features. It appears that the 501Y mutation in the spike protein leads to stronger binding to the human cellular receptor ACE2, allowing the virus to infect more productively. It appears that the E484K mutation, along with other mutations, may result in escape from vaccines and antibodies.
Even more of concern is the fact that the variants now being found have an accumulation of multiple mutations. This is predictive of such variants being more resistant to drugs and vaccines. They are likely to have been selected against drug pressure or immune system pressure, and thus would have resistance to antibody drugs, as well as other commonly used drugs such as remdesivir, It has already been found that most of the new variants are resistant to existing antibody drugs. Some of the new variants may also be resistant to remdesivir, since a WHO report stated that remdesivir was not very effective in the field, in contrast to its established effectiveness in randomized clinical trials by Gilead.
Of note, the currently approved drugs, namely remdesivir, the Regeneron antibody cocktail, or the Eli Lilly antibody drug, had demonstrated only moderate effectiveness in clinical trials. Remdesivir reduced the length of hospital stay in marginally sick hospitalized patients by approximately one to two days. The Eli Lilly antibody drug, given alongside remdesivir, improved this by another one day. The Regeneron drug dosing is at 8g of antibody, indicating relatively weak effectiveness.
Further, it is known that a new variant can cause infection of a previously recovered coronavirus patient, and sometimes may be more severe than the earlier infection. Thus, further loss of effectiveness of the existing drugs as new variants emerge would have devastating consequences Vaccines, it appears, are not the great hope nor panacea that the scientific community had once thought for this epidemic. The South African variant, 501Y.v2 is of great concern as scientists believe it may escape current vaccines. Its mutations are also shared by the Brazilian variant, P.1. Vaccinated persons coming down with SARS-CoV-2 infection has already been witnessed. As more variants emerge, existing vaccines would probably be practically useless.
An additional concern is that some of the variants are expected to result in greater total fatality numbers. A more contagious variant would cause more number of cases and thus greater number of fatalities. A more lethal variant would lead to a greater proportion of infected patients dying (i.e. a greater case fatality rate). The UK B.1.1.7 variant has both of these features, and thus presents a great risk. It is reportedly more lethal than the earlier prevalent strain, and, additionally, it is significantly more contagious.
Failure of vaccines and antibody drugs is therefore certain, the only question is how long will it be before the original vaccines become completely useless. Replacing current vaccine with a new vaccine, would be an endless game of chasing a rapidly changing epidemic that would be both costly and would be substantially non-responsive.
G: Host Defence Therapy
Dexamethasone: A moderate dose of dexamethasone (6 mg daily for 10 days) reduced mortality in hospitalized patients with COVID-19 who required supplemental oxygen therapy or mechanical ventilation [156, 157]. These favorable findings are supported by three other trials of glucocorticoids for COVID-19, which stopped enrolment in early June, 2020, when the RECOVERY trial results were released [158, 159, 160]. Each of these trials showed some evidence of benefit, although none had completed enrolment. A prospective meta-analysis of these and other trials, totaling 1703 participants (59% from the RECOVERY trial), confirmed a reduction in 28-day mortality with minimal heterogeneity across studies [22].
While confirming beneficial effects of corticosteroids for critically ill hospitalized patients with COVID-19, some unanswered questions and issues remain that deserve discussion and should be addressed in future research.
Corticosteroids: Corticosteroids, such as hydrocortisone and dexamethasone, have anti-inflammatory, antifibrotic, and vaso-constrictive effects with acute respiratory distress syndrome (ARDS) and septic shock. The publication of these 3 randomized trials of corticosteroids and the prospective meta-analysis in this issue of JAMA represents an important step forward in the treatment of patients with COVID-19. However, numerous other studies limits the confidence in using corticosteroids in hospitalized patients with COVID-19 [161]. IL and Interferons: Effective therapeutic approaches are urgently needed to reduce the spread of the virus and its death toll. An unique role of IFNλ in fine-tuning antiviral immunity in the respiratory tract prevents the devastating consequences of COVID-19-mediated pneumonia and acute respiratory distress syndrome (ARDS) [162].
H: Traditional Antivirals • Antiviral agents that target membrane-bound corona-viral RNA synthesis: Represent a novel and attractive approach [163, 163]. Inhibition of membrane- bound corona viral RNA synthesis byK22, [(Z)-N-(3- (4-(4-bromophenyl)-4-hydroxypiperidin-1-yl)-3-oxo- 1-phenylprop-1-en-2-yl) benzamide], showed potent antiviral activity of MERS-CoV however, never applied to other CoVs including SARS-CoV-2 [163, 165]. • Antiviral effect of protease inhibitors: Disulfiram, generally used for alcohol-aversion therapy, can inhibit papain-like protease and can block viral evasion [166, 167]. However, Disulfiram inhibits papain-like proteases in MERS and SARS via different mechanisms [168]. • Replicase Inhibitors: Virus replication in host cells depends on a crucial protein Helicase (nsP13) [163, 169]. Development of a small 1,2,4-triazole derivative demonstrated an inhibition of NTPase/helicase of SARS- and MERS-CoVs and low cytotoxicity [170, 171]. • Antivirals Targeting the viral entry: Camostat and Nafamostat, inhibitors of TMPRSS2, have shown effectiveness against coronaviruses entry to the host cells (e.g. MERS-CoV) [172, 173]. Further, endocytic entry of β-coronaviruses can be blocked by Imatinib, an Abelson (Abl) kinase inhibitor [174]. All of these potential therapeutics inhibited the replication of MERS- CoV, SARS-CoV, and HCoV-229E in the low-micromolar range, which is an indicative of their use as a broad- spectrum antiviral remedies [163]. • Anti-ebola drug: Remdesivir (GS-5374), a nucleotide analog that was developed as a therapy for Ebola and Marburg virus infections showed antiviral activity against Respiratory Syncytial virus, Junin virus, Lassa Fever virus, and MERS-CoV [163, 175]. This analog is also effective against Bangladesh genotypes of Nipah virus, in vitro [176]. Indeed, very recent it was published that Remdesivir can inhibit SARSCoV- 2 in cell culture studies [177, 178]. • Antimalarial drugs: Chloroquine, generally known as an anti-malarial drug, interferes with the terminal glycosylation of ACE2, and thus inhibits virus-receptor binding and thereby entry to the host cell [179]. Furthermore, an endosomal acidification which is needed for viral genome release in the host-cell cytoplasm, may be inhibited with Chloroquine or its more potent chemical derivate Hydroxychloroquine which have shown promise in testing with SARS-CoV-2 in vitro [179, 180, 181]. Recently, Wang et al. and Liu et al. have shown that the chloroquine can inhibit SARS-CoV-2 in vitro effectively, though the efficacy vs. toxicity remains as a big issue [182, 183]. • Some concerns of the above therapeutic approaches: The anti-malarial drugs, chloroquine, and hydroxychloroquine were found to be effective in limited cases of COVID-19, and also have significant side effects, including increased case fatality rates [177, 184, 185].
Prospective Therapies
Attention needs to be focused on broad-spectrum antiviral therapeutics that minimize the possibility that virus variants would escape the drug. If such an effective drug is available, vaccines that require constant changes would not be needed.
NanoViricides, Inc. (Shelton, CT, USA) is probably the only company that has developed such a technology to have broad spectrum two arms antiviral capacity. It is the only company to the best of our knowledge that can develop antiviral drugs that (a) directly attack the virus and disable them from infecting human cells, and simultaneously, (b) block the reproduction of the virus that has already gone inside a cell. Together, this double-whammy would result in a cure.
Additionally, the Nanoviricides® platform technology is based on biomimetic engineering that copies the features of the human cellular receptor of the virus. No matter how much the virus mutates, all virus variants bind to the same receptor in the same fashion. In fact, the later variants bind to the receptor more effectively, in general. Thus, if these features of the cellular receptor are appropriately copied, the resulting Nanoviricide drug would remain effective against current and future variants of a virus.
In brief, we have developed a “Venus-FlyTrap” for virus particles which is designed with such an unique concept that a when a virus encounters our Nanoviricide “Venus-FlyTrap” thinking it is binding to its target cell receptor(s), and would get engulfed and destroyed in the process (Figure 2). Using a plug-and-play approach, we can change the virus binding ligand portion of this nanomedicine to attack a different virus. This allows for rapid design & construction, synthesis, manufacture, and testing of novel drug candidates against an emerging outbreak such as the SARS-CoV-2 that has now become pandemic.
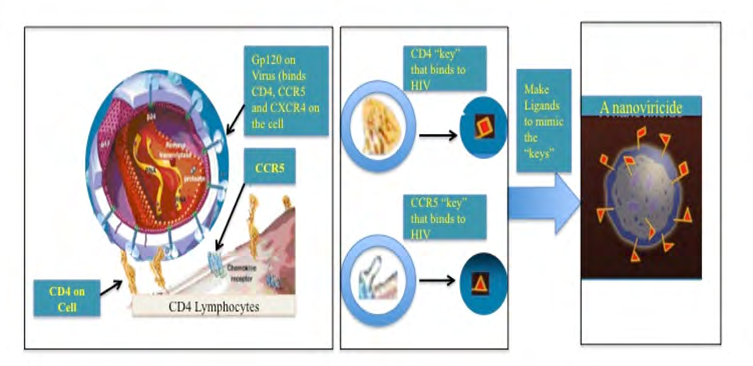
A nanoviricide “look like” a human cell o the virus. Nanoviricide is large enough for a virus particle to latch onto it. Yet small enough to circulate readily in the body. Rather than the virus particle entering into a nanoviricide, a nanoviricide wraps around the virus particle and encapsulates it, by using the virus particle’s very same ability to enter a cell.
Viral resistance to the nanoviricide drug is unlikely because even as the virus mutates, it still binds to the same cell surface receptor(s), in the same fashion. Our Recent Results We have already found SARS-CoV-2 binding ligands using molecular modeling.
- We have synthesized several Nanoviricides based on these virtual hits.
- We have already tested them for broad-spectrum anti- coronavirus effectiveness in cell culture studies. One of the coronavirus strains (NL63) we studied uses the same cell surface receptor ACE2 (angiotensin converting enzyme-2) that is shared by SARS-CoV-2 and the older coronavirus SARS-CoV-1.
- In order to assess how broad-spectrum effect our drug candidates may possess we have tested our compounds against two different coronaviruses, 229E and NL63, in this cell culture study.
- Out of several of our drug candidates, one drug showed as much as 15-TIMES more effective than favipiravir (an RNA polymerase inhibitor used here as a positive control) against two different coronaviruses in this study (Data is in the process of writing an experimental paper).
- Safety and Tolerability of anti-coronavirus drug candidates observed in an animal model, announced on July 8, 2020, further advances NanoViricides’ SARS-CoV-2 therapeutics program. Three different drug candidates at three different dosage levels (low, medium, and high) and vehicle control were administered to separate groups of mice intravenously in the Safety-Tolerability study reported here. Clinical observations and gross post-mortem studies have been completed. The tested drug candidates were safe and well tolerated, thereby clearing the path for further development towards a treatment for SARS-CoV-2 infection that has caused the current COVID-19 pandemic.
- There were no clinical signs of immune or allergic reactions such as itching, biting, twitching, rough coat, etc. Further, there were no observable changes in any organs including large intestine or colon on post mortem in gross histology. The only reportable changes observed were, in the high dosage groups of two of the three drug candidates tested, associated with the non-absorption of water, in the colon. This is consistent with the clinical observation of very loosened stools in the same groups. In clinical usage, the drug candidates are not anticipated to be administered in such high levels. The objective of this study was to discover the dosage level at which such an effect may occur. Sixteen mice in each group (8 males, 8 females), were administered one of the three drug candidates at one of the three dose levels, and additionally, one group was administered vehicle control, for seven days by daily tail-vein intravenous infusion in this blinded study with additional evaluations on 8th day. This non-GLP safety/tolerability study was conducted under GLP-like conditions by AR BioSystems, Inc., Odessa, Tampa, FL. Further microscopic histology and blood work analyses are in progress.
We have already scaled up production of key portions to multi-kilogram scales. We are now initiating an animal model study to finalize two to three best candidates for further testing. We intend to perform certain animal model safety studies, in order to further advance the final candidate for limited human clinical (compassionate) use scenario. This is the fastest timeframe that a drug candidate truly directed at the SARS-CoV-2 has been developed by any Company to date [186].
NanoViricides has diligently engaged in the fight against SARS-CoV-2 since very early in the epidemic. Our drug candidates began core safety pharmacology studies required for entering human clinical trials around October/ November, 2019. The studies have been completed and we are anticipating the report from the external CRO shortly.
We are now working on filing a pre-IND application with the US FDA as soon as possible. We are actively seeking opportunities to engage appropriate sites for human clinical trials in preparation of clinical trial protocols and filing of an IND with the US FDA.
Discussion and Conclusion
We have previously reported that these drug candidates have shown strong effectiveness in a lethal lung infection model in rats using a coronavirus that uses the same ACE2 receptor as SARS-CoV-2 which causes COVID-19, namely hCoV-NL63. We have found that hCoV-NL63, which causes a milder disease than SARS-CoV-2, causes substantially similar clinical pathology in this efficacy animal model as has been reported for SARS-CoV-2 associated lung infections in humans [187].
In those previous reports with lethal direct-lung-infection model efficacy study, animals in all groups developed lung disease which later led to multi-organ failures, a clinical pathology resembling that of the SARS-CoV-2. Reduction in loss of body weight at day 7 was used as the primary indicator of drug effectiveness. Rats were infected directly into lungs with lethal amounts of hCoV-NL63 virus particles and then different groups were treated separately with five different nanoviricides drug candidates, remdesivir as a positive control, and the vehicle as a negative control. The treatment was intravenous by tail-vein injection once daily for five days, except in the case of remdesivir wherein it was by tail-vein injection twice daily. In that efficacy study, animals treated with the five different nanoviricides showed significantly reduced body weight loss. The body weight loss in female animals ranged from only 3.9% to 11.2% in the different nanoviricide-treated groups, as compared to 20% in vehicle-treated control group, and 15.2% in a remdesivir- treated group (n=5 in each group). The body weight loss in male animals ranged from 8.0% to 10.9% in the different nanoviricides-treated groups, as compared to 25% in the vehicle-treated control group, and 18.6% in remdesivir- treated group (n=5 in each group). Smaller numbers mean less loss in body weight compared to starting body weight in the group, and indicate greater drug effectiveness.
The strong effectiveness of nanoviricide drug candidates in the lung-infection model is consistent with the effectiveness observed in cell culture studies against infection of both hCoV-NL63, which was used in the efficacy study, and hCoV- 229E, another circulating coronavirus that uses a distinctly different receptor, namely APN (aminopeptidase).
Prior to filing for human clinical trials, we plan on conducting studies, towards clinical candidate selection, to further determine the effectiveness against SARS-CoV-2, perform additional drug development studies as may be necessary, and request a pre-IND Meeting with the US FDA for regulatory guidance.
Taken together, our approach and results give a strong confidence that: (a) our broad-spectrum anti-coronavirus drug candidates would most likely be effective against SARS- CoV-2, and (b) potential mutations in the virus are unlikely to enable it to escape these drug candidates.
It is needless to say that in the present days, rapid design & construction, as well as synthesis and manufacture of anti SARS-CoV-2 has a crying need. Although our approach are very potential for the COVID-19 therapy, but we have been limited to studying effectiveness against available BSL2 level strains of coronaviruses, as they do not cause the severe pathology in humans. No other drug currently available in the development for SARS-CoV-2 virus in the USA, be it antibody, small chemical, or otherwise, has been tested against any coronaviruses at present, to the best of our knowledge.
Conflict of interests
Authors, Anil Diwan, Brian, Jayant Tatake, and Randall Barton were employed by the company Nanoviricides, Inc. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The study received funding from Nanoviricides, Inc. The funder had the following role in study conception, design, execution, analysis and decision to publish: Nanoviricides, Inc.
Acknowledgement
We acknowledge all our colleagues, Secretaries for their help during the preparation of the manuscript by providing all the relevant information. Funds are from Nanoviricide, Inc., company and they are hereby acknowledged.
References
-
Feng Y, Ling Y, Bai T, Xie Y, Huang J, Li J, et al. (2020) COVID-19 with Different Severities: A Multicenter Study of Clinical Features. Am J Respir Crit Care Med 201(11): 1380-1388.
-
(2020) WHO Coronavirus Disease (COVID-19) Dashboard.
-
Andersen KG, Rambaut A, Lipkin WA, Holmes EC, Garry RF (2020) The proximal origin of SARS-CoV-2. Nature Medicine 26(4): 450-452.
-
Singhal T (2020) A Review of Coronavirus Disease-2019 (COVID-19). Indian J Pediatr 87(4): 281-286.
-
Chakraborty A, Friedrich B, Tatake J, Chiniga V, Pandey R, et al. (2020) COVID-19: Search for Therapeutics. Integrative Molecular Medicine (Open access). Integr Mol Med 7: 1-9.
-
Meyerowitz EA, Richterman A, Gandhi RT, Sax PE (2021) Transmission of SARS-CoV-2: A Review of Viral, Host, and Environmental Factors. Ann Intern Med 174(1): 69- 79.
-
Birgand G, Peiffer-Smadja N, Fournier S, Kerneis S, Lescure FX, et al. (2020) Assessment of Air Contamination by SARS-CoV-2 in Hospital Settings. JAMA Netw Open 3(12): e2033232.
-
Ng K, Poon BH, Kiat Puar TH, Shan Quah JL, Loh WJ, et al. (2020) 2020COVID-19 and the Risk to Health Care Workers: A Case Report. Ann Intern Med 172(11): 766- 767.
-
Wong SCY, Kwong RT, Wu TC, Chan JWM, Chu MY, et al. (2020) Risk of nosocomial transmission of coronavirus disease 2019: an experience in a general ward setting in Hong Kong. J Hosp Infect 105(2): 119-127.
-
Chen W, Lan Y, Yuan X, Deng X, Li Y, et al. (2020) Detectable 2019-nCoV viral RNA in blood is a strong indicator for the further clinical severity. Emerg Microbes Infect 9(1): 469-473.
-
Wang W, Xu Y, Gao R, Lu R, Han K, et al. (2020) Detection of SARS-CoV-2 in Different Types of Clinical Specimens. JAMA 323(18): 1843-1844.
-
Colavita F, Lapa D, Carletti F, Lalle E, Bordi L, et al. (2020) SARS-CoV-2 Isolation From Occular Secretions of a Patient With COVID-19 in Italy With Prolonged Viral RNA Detection. Ann Intern Med 173(3): 242-243.
-
Cheung KS, Hung IFN, Chan PPY, Lung KC, Tso E, et al. (2020) Gastrointestinal Manifestations of SARS-CoV-2 infection and Virus Load in Fecal Samples From a Hong Kong Cohort: Systematic Review and Meta-analysis. Gastroenterology 159(1): 81-95.
-
Zheng S, Fan J, Yu F, Feng B, Lou B, et al. (2020) Viral load dynamics and disease severity in patients infected with SARS-CoV-2 in Zhejiang province, China, January-March 2020: retrospective cohort study. BMJ 369: m1443.
-
Yu F, Yan L, Wang N, Yang S, Wang L, et al. (2020) Quantitative Detection and Viral Load Analysis of SARS- CoV-2 in Infected Patients. Clin Infect Dis 71(15): 793- 798.
-
Xu D, Zhou F, Sun W, Chen L, Lan L, et al. (2020) Relationship Between serum SARS-CoV-2 nucleic acid (RNAemia) and Organ Damage in COVID-19 Patients: A Cohort Study. Clin Infect Dis pp: ciaa1085.
-
AABB. AABB’s Coronavirus Resources.
-
Zou L, Ruan F, Huang M, Liang L, Huang H, et al. (2020) SARS-CoV-2 Viral Load in Upper Respiratory Specimens of Infected Patients. N Engl J Med 382(12): 1177-1179.
-
To KK, Tsang OT, Leung WS, Leung WS, Tam AR, et al. (2020) Temporal profiles of viral load in posterior oropharyngeal saliva samples and serum antibody responses during infection by SARS-CoV-2: an observational cohort study. Lancet Infect Dis 20(5): 565- 74.
-
Munnink BBO, Sikkema RS, Nieuwenhuijse DF, Molenaar RJ, Munger E, et al. (2021) Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans. Science 371(6525): 172-177.
-
European Ministry of Agriculture, Nature and Food Quality. SARS-CoV-2 infections of mink in the Netherlands.
-
WHO (2020) SARS-CoV-2 mink-associated variant strain –Denmark.
-
Rijkers G, Murk JL, Wintermans B, van Looy B, van den Berge M, et al. (2020) Differences in Antibody Kinetics and Functionality Between Severe and Mild Severe Acute Respiratory Syndrome Coronavirus 2 Infections. J Infect Dis 222(8): 1265-1269.
-
Lynch KL, Whitman JD, Lacanienta NP, Beckerdite EW, Kastner SA, et al. (2021) Magnitude and kinetics of anti- SARS-CoV-2 antibody responses and their relationship to disease severity. Clin Infect Dis 72(2): 301-308.
-
Long QX, Tang XJ, Shi QL, Li Q, Deng HJ, et al. (2020) Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections. Nat Med 26(8):1200-1204.
-
Wang K, Long QX, Deng HJ, Hu J, Gao QZ, et al. (2020) Longitudinal dynamics of the neutralizing antibody response to SARS-CoV-2 infection. Clin Infect Dis ciaa1143.
-
Crawford KHD, Dingens AS, Eguia R, Wolf CR, Wilcox N, et al. (2021) Dynamics of Neutralizing Antibody Titers in the Months After Severe Acute Respiratory Syndrome Coronavirus 2 Infection. J Infect Dis 223(2): 197-205.
-
Wajnberg A, Amanat F, Firpo A, Altman DR, Bailey MJ, et al. (2020) Robust neutralizing antibodies to SARS-CoV-2 infection persist for months. Science 370(6521):1227- 1230.
-
Rodda LB, Netland J, Shehata L, Pruner KB, Morawski PA, et al. (2021) Functional SARS-CoV-2-Specific Immune Memory Persists after Mild COVID-19. Cell 184(1):169- 183.e17.
-
Lumley SF, O’Donnell D, Stoesser NE, Matthews PC, Howarth A, et al. (2021) Antibody Status and Incidence of SARS-CoV-2 Infection in Health Care Workers. N Engl J Med 384(6): 533-540.
-
Zhu FC, Li YH, Guan XH, Hou LH, Wang WJ, et al. (2020) Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: a dose- escalation, open-label, non-randomized, first-in-human trial. Lancet 395(10240): 1845-1854.
-
Grifoni A, Weiskopf D, Ramirez SI, Mateus J, Dan JM, et al. (2020) Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 181(7):1489-1501.e15.
-
Deng W, Bao L, Liu J, Xiao C, Liu J, et al. (2020) Primary exposure to SARS-CoV-2 protects against reinfection in rhesus macaques. Science 369(6505): 818-823.
-
Corbett KS, Flynn B, Foulds KE, Francica JR, Boyoglu- Barnum S, et al. (2020) Evaluation of the mRNA-1273 Vaccine against SARS-CoV-2 in Nonhuman Primates. N Engl J Med 383(16): 1544-1555.
-
Mercado NB, Zahn R, Wegmann F, Loos C, Chandrashekar A, et al. (2020) Single-shot Ad26 vaccine protects against SARS-CoV-2 in rhesus macaques. Nature 586(7830): 583-588.
-
van Doremalen N, Lambe T, Spencer A, Belij- Rammerstorfer S, Purushotham JN, et al. (2020) ChAdOx1 nCoV-19 vaccine prevents SARS-CoV-2 pneumonia in rhesus macaques. Nature 586(7830): 578-582.
-
Mateus J, Grifoni A, Tarke A, Sidney J, Ramirez SI, et al. (2020) Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 370(6512): 89- 94.
-
Braun J, Loyal L, Frentsch M, Wendisch D, Georg P, et al. (2020) SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 587(7833): 270- 274.
-
Ng KW, Faulkner N, Cornish GH, Rosa A, Harvey R, et al. (2020) Preexisting and de novo humoral immunity to SARS-CoV-2 in humans. Science 370(6552): 1339-1349.
-
To KK, Hung IF, Ip JD, Chu AW, Chan WM, et al. (2020) COVID-19 re-infection by a phylogenetically distinct SARS-coronavirus-2 strain confirmed by whole genome sequencing. Clin Infect Dis ciaa1275.
-
Van Elslande J, Vermeersch P, Vandervoort K, Wawina- Bokalanga T, Vanmechelen B, et al. (2021) Symptomatic Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Reinfection by a Phylogenetically Distinct Strain. Clinical Infectious Diseases 73(2): 354-356.
-
Gupta V, Bhoyar RC, Jain A, Srivastava S, Upadhayay R, et al. (2020) Asymptomatic reinfection in two healthcare workers from India with genetically distinct SARS-CoV-2. Clinical Infectious Diseases ciaa1451.
-
Tillett RL, Sevinsky JR, Hartley PD, Kerwin H, Crawford N, et al. (2020) Genomic evidence for reinfection with SARS-CoV-2: a case study. Lancet Infect Dis 21(1): 52-58.
-
Common Human Coronaviruses.
-
Cascella M, Rajnik M, Aleem A, Dulebohn SC, Di Napoli R (2021) Features, Evaluation, and Treatment of Coronavirus (COVID-19). In: StatPearls [Internet].
-
Fehr AR, Perlman S (2015) Coronaviruses: An Overview of Their Replication and Pathogenesis. In: Maier H, Bickerton E, Britton P (Eds.), Coronaviruses. Methods in Molecular Biology, Humana Press, New York, NY 1282: 1-23.
-
Ziebuhr J (2005) The Coronavirus Replicase. In: Enjuanes L (Eds.), Coronavirus Replication and Reverse Genetics. Current Topics in Microbiology and Immunology, Springer, Berlin, Heidelberg 287: 57-94.
-
Lim YX, Ng YL, Tam JP, Liu DX (2016) Human Coronaviruses: A Review of Virus–Host Interactions. Diseases 4(3): 26.
-
Schoeman D, Fielding BC (2019) Coronavirus Envelope Protein: Current Knowledge. Virology Journal 16: 69.
-
Dijkman R, Mulder HL, Rumping L, Kraaijvanger I, Deijs M, et al. (2009) Seroconversion to HCoV-NL63 in Rhesus Macaques. Viruses 1(3): 647-56.
-
Salanueva IJ, Carrascosa JL, Risco C (1999) Structural maturation of the transmissible gastroenteritis coronavirus. J Virol 73(10): 7952-7964.
-
Le TT, Andreadakis Z, Kumar A, Román RG, Tollefsen S, et al. (2020) The COVID-19 vaccine development landscape. Nature Reviews (Drug Discovery) 19: 305- 306.
-
Schlake T, Thess A, Fotin-Mleczek M, Kallen KJ (2012) Developing mRNA-vaccine technologies. RNA Biology 9(11): 1319-1330.
-
(2020) Pfizer Press Release.
-
(2020) Moderna news release.
-
Chi X, Yan R, Zhang J, Zhang G, Zhang Y, et al. (2020) A neutralizing human antibody binds to the N- terminal domain of the Spike protein of SARS- CoV-2. Science 369(6504): 650-655.
-
Jiang S, Zhang X, Du L (2020) Therapeutic antibodies and fusion inhibitors targeting the spike protein of SARS- CoV-2. Expert Opin Ther Targets 17: 1-7.
-
Zhu J, Xiao G, Xu Y, Yuan F, Zheng C, et al. (2004) Following the rule: formation of the 6-helix bundle of the fusion core from severe acute respiratory syndrome coronavirus spike protein and identification of potent peptide inhibitors. Biochem Biophys Res Commun 319(1): 283-288.
-
Sun H, Li Y, Liu P, Qiao C, Wang X, et al. (2020) Structural basis of HCoV-19 fusion core and an effective inhibition peptide against virus entry. Emerg Microbes Infect 9(1): 1238-1241.
-
Thomas L, Richards M, Mort M, Dunlop E, Cooper DN, et al. (2012) Assessment of the potential pathogenicity of missense mutations identified in the GTPase-activating protein (GAP)-related domain of the neurofibromatosis type-1 (NF1) gene. Hum Mutat 33(12): 1687-1696.
-
Masters SP, Perlman S (2013) Fields virology. 6th (Edn.), Ch. 28 Knipe DM, Howley PM (Eds.), pp: 825-858
-
Du L, He Y, Jiang S, Zheng BJ (2008) Development of subunit vaccines against severe acute respiratory syndrome. Drugs Today (Barc) 44(1): 63-73.
-
Sun J, Zhuang Z, Zheng J, Li K, Wong RL, et al. (2020) Generation of a Broadly Useful Model for COVID-19 Pathogenesis, Vaccination, and Treatment. Cell 182(3): 734-743.e5.
-
Liu WJ, Zhao M, Liu K, Xu K, Wong G, et al. (2017) T-cell immunity of SARS-CoV: Implications for vaccine development against MERS-CoV. Antiviral Res 137: 82-
-
Long QX, Liu BZ, Deng HJ, Wu GC, Deng K, et al. (2020) Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat Med 26(6): 845-848.
-
Sariol A, Perlman S (2020) Lessons for COVID-19 immunity from other coronavirus infections. Immunity 53(2): 248-263.
-
Nakanaga K, Yamanouchi K, Fujiwara K (1986) Protective effect of monoclonal antibodies on lethal mouse hepatitis virus infection in mice. J Virol 59(1): 168-171.
-
Lecomte J, Cainelli-Gebara V, Mercier G, Mansour S, Talbot PJ, et al. (1987) Protection from mouse hepatitis virus type 3-induced acute disease by an anti-nucleoprotein monoclonal antibody. Archives of Virology 97(1-2): 123- 130.
-
Liu SJ, Leng CH, Lien SP, Chi HY, Huang CY, et al. (2006) Immunological characterizations of the nucleocapsid protein based SARS vaccine candidates. Vaccine 24(16): 3100-3108.
-
Collisson EW, Pei J, Dzielawa J, Seo SH (2000) Cytotoxic T lymphocytes are critical in the control of infectious bronchitis virus in poultry. Dev Comp Immunol 24(2-3): 187-200.
-
Zhao J, Zhao J, Mangalam AK, Channappanavar R, Fett C, et al. (2016) Airway Memory CD4(+) T Cells Mediate Protective Immunity against Emerging Respiratory Coronaviruses. Immunity 44(6): 1379-1391.
-
Deming D, Sheahan T, Heise M, Yount B, Davis N, et al. (2006) Vaccine efficacy in senescent mice challenged with recombinant SARS-CoV bearing epidemic and zoonotic spike variants. PLoS Med 3(12): e525.
-
Yasui F, Kai C, Kitabatake M, Inoue S, Yoneda M, et al. (2008) Prior immunization with severe acute respiratory syndrome (SARS)-associated coronavirus (SARS-CoV) nucleocapsid protein causes severe pneumonia in mice infected with SARS-CoV. J Immunol 181(9): 6337-6348.
-
Chen Q, Quan B, Li X, Gao G, Zheng W, et al. (2020) A report of clinical diagnosis and treatment of nine cases of coronavirus disease 2019. J Med Virol 92(6): 683-687.
-
Lu G, Wang Q, Gao GF (2015) Bat-to-human: spike features determining ‘host jump’ of coronaviruses SARS- CoV, MERS-CoV, and beyond. Trends Microbiol 23(8): 468-478.
-
Lu G, Hu Y, Wang Q, Qi J, Gao F, et al. (2013) Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26. Nature 500: 227-231.
-
Grant OC, Montgomery D, Ito K, Woods RJ (2020) Analysis of the SARS- CoV-2 spike protein glycan shield reveals implications for immune recognition. Sci Rep 10(1): 14991.
-
Guo ZD, Wang ZY, Zhang SF, Li X, Li L, et al. (2020) Aerosol and Surface Distribution of Severe Acute Respiratory Syndrome Coronavirus 2 in Hospital Wards, Wuhan, China, 2020. Emerg Infect Dis 26(7): 1583-1591.
-
Robbiani DF, Gaebler C, Muecksch F, Lorenzi JCC, Wang Z, et al. (2020) Convergent antibody responses to SARS- CoV-2 in convalescent individuals. Nature 584(7821): 437-442.
-
Ju B, Zhang Q, Ge J, Wang R, Sun J, et al. (2020) Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 584(7819): 115-119.
-
Shi R, Shan C, Duan X, Chen Z, Liu P, et al. (2020) A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 584(7819): 120-124.
-
Du S, Cao Y, Zhu Q, Yu P, Qi F, et al. (2020) Structurally resolved SARS- CoV-2 antibody shows high efficacy in severely infected hamsters and provides a potent cocktail pairing strategy. Cell 183(4): 1013-1023.e13.
-
Zost SJ, Gilchuk P, Case JB, Binshtein E, Chen RE, et al. (2020) Potently neutralizing and protective human antibodies against SARS-CoV-2. Nature 584(7821): 443- 449.
-
Wu Y, Wang F, Shen C, Peng W, Li D, et al. (2020) A noncompeting pair of human neutralizing antibodies block COVID-19 virus binding to its receptor ACE2. Science 368(6496): 1274-1278.
-
Brouwer PJM, Caniels TG, van der Straten K, Snitselaar JL, Aldon Y, et al. (2020) Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. Science 369 (6504): 643-650.
-
Wec AZ, Wrapp D, Herbert AS, Maurer DP, Haslwanter D, et al. (2020) Broad neutralization of SARS-related viruses by human monoclonal antibodies. Science 369(6504): 731-736.
-
Rogers TF, Zhao F, Huang D, Beutler N, Burns A, et al. (2020) Isolation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model. Science 369(6506): 956-963.
-
Hansen J, Baum A, Pascal KE, Russo V, Giordano S, et al. (2020) Studies in humanized mice and convalescent humans yield a SARS-CoV-2 antibody cocktail. Science 369(6506): 1010-1014.
-
Barnes CO, West AP, Huey-Tubman KE, Hoffmann MAG, Sharaf NG, et al. (2020) Structures of Human Antibodies Bound to SARS-CoV-2 Spike Reveal Common Epitopes and Recurrent Features of Antibodies. Cell 182(4): 828- 842.e16.
-
Cao Y, Su B, Guo X, Sun W, Deng Y, et al. (2020) Potent Neutralizing Antibodies against SARS-CoV-2 Identified by High-Throughput Single-Cell Sequencing of Convalescent Patients’ B Cells. Cell 182(1): 73-84.e16.
-
Rey FA, Stiasny K, Vaney MC, Dellarole M, Heinz FX (2018) The bright and the dark side of human antibody responses to flaviviruses: lessons for vaccine design. EMBO Rep 19 (2): 206-224.
-
Hohdatsu T, Nakamura M, Ishizuka Y, Yamada H, Koyama H (1991) A study on the mechanism of antibody- dependent enhancement of feline infectious peritonitis virus infection in feline macrophages by monoclonal antibodies. Arch Virol 120(3-4): 207-217.
-
Weiss RC, Scott FW (1981) Antibody-mediated enhancement of disease in feline infectious peritonitis: comparisons with dengue hemorrhagic fever. Comp Immunol Microbiol Infect Dis 4(2): 175-189.
-
Zhang NN, Li XF, Deng YQ, Zhao H, Huang YJ, et al. (2020) A Thermostable mRNA Vaccine against COVID-19. Cell 182(5): 1271-1283.e16.
-
Yang J, Wang W, Chen Z, Lu S, Yang F, et al. (2020) A vaccine targeting the RBD of the S protein of SARS-CoV-2 induces protective immunity. Nature 586(7830): 572- 577.
-
Zhou M, Xu D, Li X, Li H, Shan M, et al. (2006) Screening and identification of severe acute respiratory syndrome- associated coronavirus-specific CTL epitopes. J Immunol 177(4): 2138-2145.
-
Jiang S, Bottazzi ME, Du L, Lustigman S, Tseng CT, et al. (2012) Roadmap to developing a recombinant coronavirus S protein receptor-binding domain vaccine for severe acute respiratory syndrome. Expert Rev Vaccines 11(12): 1405-1413.
-
Wang CB (2020) Analysis of low positive rate of nucleic acid detection method used for diagnosis of novel coronavirus pneumonia. Zhonghua Yi Xue Za Zhi 100(13): 961-964.
-
Zhou Y, Yang Y, Huang J, Jiang S, Du L (2019) Advances in MERS-CoV Vaccines and Therapeutics Based on the Receptor-Binding Domain. Viruses 11(1): 60.
-
Yu J, Tostanoski LH, Peter L, Mercado NB, McMahan K, et al. (2020) DNA vaccine protection against SARS- CoV-2 in rhesus macaques. Science 369(6505): 806-811.
-
Gu H, Chen Q, Yang G, He L, Fan H, et al. (2020) Adaptation of SARS-CoV-2 in BALB/c mice for testing vaccine efficacy. Science 369(6511): 1603-1607.
-
Du L, Zhao G, He Y, Guo Y, Zheng BJ, et al. (2007) Receptor-binding domain of SARS-CoV spike protein induces long-term protective immunity in an animal model. Vaccine 25(15): 2832-2838.
-
Du L, Kou Z, Ma C, Tao X, Wang L, et al. (2013) A truncated receptor-binding domain of MERS-CoV spike protein potently inhibits MERS-CoV infection and induces strong neutralizing antibody responses: implication for developing therapeutics and vaccines. PLoS One 8(12): e81587.
-
Ma C, Wang L, Tao X, Zhang N, Yang Y, et al. (2014) Searching for an ideal vaccine candidate among different MERS coronavirus receptor-binding fragments--the importance of immunofocusing in subunit vaccine design. Vaccine 32(46): 6170-6176.
-
He Y, Zhou Y, Liu S, Kou Z, Li W, et al. (2004) Receptor- binding domain of SARS-CoV spike protein induces highly potent neutralizing antibodies: implication for developing subunit vaccine. Biochem Biophys Res Commun 324(2): 773-781.
-
Wang C, Zheng X, Gai W, Wong G, Wang H, et al. (2017) Novel chimeric virus-like particles vaccine displaying MERS-CoV receptor-binding domain induce specific humoral and cellular immune response in mice. Antiviral Res 140: 55-61.
-
Kim YS, Son A, Kim J, Kwon SB, Kim MH, et al. (2018) Chaperna-Mediated Assembly of Ferritin-Based Middle East Respiratory Syndrome-Coronavirus Nanoparticles. Front Immunol 9: 1093.
-
Walls AC, Fiala B, Scha A, Wrenn S, Pham MN, et al. (2020) Elicitation of potent neutralizing antibody responses by designed protein nanoparticle vaccines for SARS- CoV-2. Cell 183(5): 1367-1382.e17
-
Dai L, Zheng T, Xu K, Han Y, Xu L, et al. (2020) A Universal Design of Betacoronavirus Vaccines against COVID-19, MERS, and SARS. Cell 182(3): 722-733.e11.
-
Mulligan MJ, Lyke KE, Kitchin N, Absalon J, Gurtman A, et al. (2020) Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 586(7830): 589-593.
-
Sahin U, Muik A, Derhovanessian E, Vogler I, Kranz LM, et al. (2020) COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 586(7830): 594-599.
-
Bachmann MF, Zinkernagel RM (1997) Neutralizing antiviral B cell responses. Annu Rev Immunol 15: 235- 270.
-
Liu L, Wang P, Nair MS, Yu J, Rapp M, et al. (2020) Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 584(7821): 450-456.
-
Wang N, Rosen O, Wang L, Turner HL, Stevens LJ, et al. (2019) Structural definition of a neutralization- sensitive epitope on the MERS- CoV S1-NTD. Cell Rep 28(13): 3395-3405.
-
Zhou H, Chen Y, Zhang S, Niu P, Qin K, et al. (2019) Structural definition of a neutralization epitope on the N- terminal domain of MERS- CoV spike glycoprotein. Nat Commun 10(1): 3068.
-
Jiaming L, Yanfeng Y, Yao D, Yawei H, Linlin B, et al. (2017) The recombinant N- terminal domain of spike proteins is a potential vaccine against Middle East respiratory syndrome coronavirus (MERS-CoV) infection. Vaccine 35(1): 10-18.
-
Huang Y, Yang C, Xu XF, Xu W, Liu SW (2020) Structural and functional properties of SARS-CoV-2 spike protein: potential antivirus drug development for COVID-19. Acta Pharmacologica Sinica 41: 1141-1149.
-
Song G, He WT, Callaghan S, Anzanello F, Huang D, et al. (2020) Cross-reactive serum and memory B cell responses to spike protein in SARS-CoV-2 and endemic coronavirus infection. bioRxiv.
-
Watanabe Y, Allen JD, Wrapp D, McLellan JS, Crispin M (2020) Site- specific glycan analysis of the SARS- CoV- 2 spike. Science 369(6501): 330-333.
-
Walls AC, Park YJ, Tortorici MA, Wall A, McGuire AT, et al. (2020) Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 181(2): 281-292.e6.
-
Ravichandran S, Coyle EM, Klenow L, Tang J, Grubbs G, et al. (2020) Antibody signature induced by SARS- CoV-2 spike protein immunogens in rabbits. Sci Transl Med 12(550): eabc3539.
-
Jeyanathan M, Afkhami S, Smaill F, Miller MS, Lichty BD, et al. (2020) Immunological considerations for COVID-19 vaccine strategies. Nat Rev Immunol 20(10): 615-632.
-
Shin MD, Shukla S, Chung YH, Beiss V, Chan SK, et al. (2020) COVID-19 vaccine development and a potential nanomaterial path forward. Nat Nanotechnol 15(8): 646-655.
-
Florindo HF, Kleiner R, Vaskovich Koubi D, Acúrcio RC, Carreira B, et al. (2020) Immune-mediated approaches against COVID-19. Nat Nanotechnol 15(8): 630-645.
-
Pardi N, Hogan MJ, Porter FW, Weissman D (2018) mRNA vaccines - a new era in vaccinology. Nat Rev Drug Discov 17(4): 261-279.
-
Pardi N, Tuyishime S, Muramatsu H, Kariko K, Mui BL, et al. (2015) Expression kinetics of nucleoside- modified mRNA delivered in lipid nanoparticles to mice by various routes. J Control Release 217: 345-351.
-
Akinc A, Maier MA, Manoharan M, Fitzgerald K, Jayaraman M, et al. (2019) The Onpattro story and the clinical translation of nanomedicines containing nucleic acid-based drugs. Nat Nanotechnol 14(12): 1084-1087.
-
Anselmo AC, Mitragotri S (2019) Nanoparticles in the clinic: An update. Bioeng Transl Med 4(3): e1043.
-
(2020) Nanomedicine and the COVID-19 vaccines. Nat Nanotechnol 15: 963.
-
Zhu FC, Guan XH, Li YH, Huang JY, Jiang T, et al. (2020) Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: a randomized, double- blind, placebo- controlled, phase 2 trial. Lancet 396(10249): 479-488.
-
van Doremalen N, Haddock E, Feldmann F, Meade White K, Bushmaker T, et al. (2020) A single dose of ChAdOx1 MERS provides protective immunity in rhesus macaques Science Advances 6(24): eaba8399.
-
van Doremalen N, Lambe T, Spencer A, Belij Rammerstorfer S, Purushotham JN, et al. (2020) ChAdOx1 nCoV-19 vaccine prevents SARS-CoV-2 pneumonia in rhesus macaques. Nature 586: 578-582.
-
Folegatti PM, Ewer KJ, Aley PK, Angus B, Becker S, et al. (2020) Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS- CoV-2: a preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet 396(10249): 467-478.
-
Zhang Y, Zeng G, Pan H, Li C, Hu Y, et al. (2021) Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18-59 years: a randomized, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect Dis 21(2): 181-192.
-
Okamura S, Kashiwabara A, Suzuki H, Ueno S, Miyazato P, et al. (2021) Live attenuated SARS-CoV-2 vaccine candidate: Protective immunity without serious lung lesions in Syrian hamsters. bioRxiv.
-
Abidin AZ, DSouza AM, Nagarajan MB, Wang L, Qiu X, et al. (2018) Alteration of brain network topology in HIV- associated neurocognitive disorder: a novel functional connectivity perspective. Neuroimage Clin 17: 768-777.
-
Wang C, Liu Z, Chen Z, Huang X, Xu M, et al. (2020) The establishment of reference sequence for SARS-CoV-2 and variation analysis. J Med Virol 92(6): 667-674.
-
Becerra Flores M, Cardozo T (2020) SARS-CoV-2 viral spike G614 mutation exhibits higher case fatality rate. Int J Clin Pract 74(8): e13525.
-
Zhou GY, Zhao Q (2020) Perspectives on therapeutic neutralizing antibodies against the novel coronavirus SARS-CoV-2. Int J Biol Sci 16(10): 1718-1723.
-
Molly Walker (2021) Just How Much More Transmissible Is the New Coronavirus Variant?. MedPage Today.
-
Horby P, Huntley C, Davies N, Edmunds J, Ferguson N, et al. (2021) NERVTAG Presented to SAGE.
-
Public Health England (2020) Investigation of SARS- CoV-2 variants of concern: technical briefings. Technical briefing documents on novel SARS-CoV-2 variants.
-
Tegally H, Wilkinson E, Giovanetti M, Iranzadeh A, Fonseca V, et al. (2021) Emergence of a SARS-CoV-2 variant of concern with mutations in spike glycoprotein. MedRxiv.
-
Wu K, Werner AP, Moliva JI, Koch M, Choi A, et al. (2021) mRNA-1273 vaccine induces neutralizing antibodies against spike mutants from global SARS- CoV-2 variants. BioRxiv.
-
National Institute of Infectious Diseases, Japan (2021) Brief report: New Variant Strain of SARS-CoV-2 Identified in Travelers from Brazil.
-
MN Department of Health (2021) MDH lab testing confirms nation’s first known COVID-19 case associate with Brazil P.1 variant.
-
Faria NR, Claro IM, Candido D, Moyses Franco LA, Andrade PS, et al. (2020) Genomic characterisation of an emergent SARS-CoV-2 lineage in Manaus: Preliminary findings. Viological.org.
-
Resende PC, Bezerra JF, De Vasconcelos RHT, Arantes I, Appolinario L, et al. (2021) Spike E484K mutation in the first SARS-CoV-2 reinfection case confirmed in Brazil, 2020. virological.org.
-
Naveca F, de Costa C, Nascimento V, Souza V, Corado A**,** et al. (2018) SARS-CoV-2 reinfection by the new Variant of Concern (VOC) P.1 in Amazonas, Brazil. virological.org
-
Volz E, Hill V, McCrone J, McCrone JT, Price A, Jorgensen D, et al. (2021) Evaluating the Effects of SARS- CoV-2 Spike Mutation D614G on Transmissibility and Pathogenicity. Cell 184(1): 64-75.
-
Korber B, Fischer WM, Gnanakaran S, Yoon H, Theiler J, et. al. (2020) Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 182(4): 812-827 e19.
-
Yurkovetskiy L, Wang X, Pascal KE, Tomkins Tinch C, Nyalile TP, et al. (2020) Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant. Cell 183(3): 739-751.
-
COVID vaccines focus on the spike protein – but here’s another target.
-
HHS (2021) COVID-19 (Coronavirus).
-
Horby P, Lim WS, Emberson JR, Mafham M, Bell JL, et al. (2021) Dexamethasone in hospitalized patients with Covid 19 preliminary report. N Engl J Med 384(8): 693- 704.
-
Keyt H (20251) WHO recommends corticosteroids for patients with severe or critical COVID-19. Ann Intern Med 174(1): JC2.
-
Dequin PF, Heming N, Meziani F, Plantefève G, Voiriot G, et al. (2020) Effect of hydrocortisone on 21- day mortality or respiratory support among critically ill patients with COVID-19: a randomized clinical trial. JAMA 324(13): 1298-1306.
-
Tomazini BM, Maia IS, Cavalcanti AB, Berwanger O, Rosa RG, et. al. (2020) Effect of Dexamethasone on Days Alive and Ventilator-Free in Patients With Moderate or Severe Acute Respiratory Distress Syndrome and COVID-19: The CoDEX Randomized Clinical Trial. JAMA 324(13): 1307-1316.
-
Angus DC, Derde L, Al-Beidh F, Annane D, Arabi Y, et. al. (2020) Effect of Hydrocortisone on Mortality and Organ Support in Patients With Severe COVID-19: The REMAP-CAP COVID-19 Corticosteroid Domain Randomized Clinical Trial. JAMA 324(13): 1317-1329.
-
Prescott HC (2020) Outcomes for Patients Following Hospitalization for COVID-19. JAMA 325(15): 1511- 1512.
-
Andreakos E, Tsiodras S (2020) COVID-19: lambda interferon against viral load and hyperinflammation. EMBO Mol Med 12(6): e12465.
-
Pillaiyar T, Wendt LL, Manickam M, Easwaran M (2021) The recent outbreaks of human coronaviruses: A medicinal chemistry perspective. Med Res Rev 41(1): 72-135.
-
Filone CM, Hodges EN, Honeyman B, Bushkin GG, Boyd K, et al. (2013) Identification of a broad-spectrum inhibitor of viral RNA synthesis: validation of a prototype virus-based approach. Chem Biol 20(3): 424-433.
-
Lundin A, Dijkman R, Bergström T, Kann N, Adamiak B, et al. (2014) Targeting membrane-bound viral RNA synthesis reveals potent inhibition of diverse coronaviruses including the middle East respiratory syndrome virus. PLoS Pathog 10(5): e1004166.
-
Paranjpe A, Zhang R, Ali-Osman F, Bobustuc GC, Srivenugopal KS (2014) Disulfiram is a direct and potent inhibitor of human O6-methylguanine-DNA methyltransferase (MGMT) in brain tumor cells and mouse brain and markedly increases the alkylating DNA damage. Carcinogenesis 35(3): 692-702.
-
Lin MH, Moses DC, Hsieh CH, Cheng SC, Chen YH, et al. (2018) Disulfiram can inhibit MERS and SARS coronavirus papain-like proteases via different modes. Antiviral Res 150: 155-163.
-
Maiti BK (2020) Can Papain-like Protease Inhibitors Halt SARS-CoV-2 Replication? ACS Pharmacol Transl Sci 3(5): 1017-1019.
-
Hertzig T, Scandella E, Schelle B, Ziebuhr J, Siddell SG, et al. (2004) Rapid identification of coronavirus replicase inhibitors using a selectable replicon RNA. J Gen Virol 85(6): 1717-1725.
-
Shen L, Niu J, Wang C, Huang B, Wang W, et al. (2019) High-Throughput Screening and Identification of Potent Broad-Spectrum Inhibitors of Coronaviruses. J Virol 93(12): e00023-19.
-
Adedeji AO, Singh K, Calcaterra NE, DeDiego ML, Enjuanes L, et al. (2012) Severe acute respiratory syndrome coronavirus replication inhibitor that interferes with the nucleic acid unwinding of the viral helicase. Antimicrob Agents Chemother 56(9): 4718- 4728.
-
Markus H (2020) SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 181(2): 271-280.
-
Yamamoto M, Matsuyama S, Li X, Takeda M, Kawaguchi Y, et al. (2016) Identification of Nafamostat as a Potent Inhibitor of Middle East Respiratory Syndrome Coronavirus S Protein-Mediated Membrane Fusion Using the Split-Protein-Based Cell-Cell Fusion Assay. Antimicrob Agents Chemother 60(11): 6532- 6539.
-
Sisk JM, Frieman MB, Machamer CE (2018) Coronavirus S protein-induced fusion is blocked prior to hemifusion by Abl kinase inhibitors. J Gen Virol 99(5): 619-630.
-
Warren T, Jordan R, Lo M. Ray AS, Mackman RL, Soloveva V (2016) Therapeutic efficacy of the small molecule GS-5734 against Ebola virus in rhesus monkeys. Nature 531: 381-385.
-
Lo MK, Feldmann F, Gary JM, Jordan R, Bannister R, et al. (2019) Remdesivir (GS-5734) protects African green monkeys from Nipah virus challenge. Sci Transl Med 11(494): eaau9242.
-
Wang M, Cao R, Zhang L, Yang X, Liu J, et al. (2020) Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res 30(3): 269-271.
-
Lo MK, Jordan R, Arvey A, Sudhamsu J, Shrivastava- Ranjan P, et al. (2017) GS-5734 and its parent nucleoside analog inhibit Filo-, Pneumo-, and Paramyxoviruses. Sci Rep 7: 43395.
-
Vincent MJ, Bergeron E, Benjannet S, Erickson BR, Rollin PE, et al. (2005) Chloroquine is a potent inhibitor of SARS coronavirus infection and spread. Virol J 2: 69-79.
-
Zhou P, Yang XL, Wang XG, Hu B, Zhang L, et al. (2020) A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579: 270-273.
-
Liu C, Zhou Q, Li Y, Garner LV, Watkins SP, et al. (2020) Research and Development on Therapeutic Agents and Vaccines for COVID-19 and Related Human Coronavirus Diseases. ACS Cent Sci 6(3): 315-331.
-
Alanagreh L, Alzoughool F, Atoum M (2020) Risk of using hydroxychloroquine as a treatment of COVID-19. Int J Risk Saf Med 31(3): 111-116.
-
Liu J, Cao R, Xu M, Wang X, Zhang H, et al. (2020) Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro. Cell Discov 6: 16.
-
Gao J, Tian Z, Yang X (2020) Breakthrough: Chloroquine phosphate has shown apparent efficacy in treatment of COVID-19 associated pneumonia in clinical studies. Biosci Trends 14(1): 72-73.
-
Yao X, Ye F, Zhang M, Cui C, Huang B, et al. (2020) In Vitro Antiviral Activity and Projection of Optimized Dosing Design of Hydroxychloroquine for the Treatment of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). Clin Infect Dis 71(15): 732-739.
-
Nanoviricides (2020) NanoViricides Develops Highly Effective Broad-Spectrum Drug Candidates Against Coronaviruses.
-
Nanoviricides (2020) NanoViricides-SARS-CoV.
- Psychogenic Erectile Dysfunction in Late Adulthood: A Case Report on Clinical Intervention and Intimacy Restoration
- Clinical Trials on COVID-19 in 2025: A New Chapter in Global Health Research
- Innovations and Challenges in Contemporary Medical Clinical Trials: An Editorial Perspective
- Innovations and Challenges in Contemporary Medical Clinical Trials: A Critical Perspective
- Reimagining Clinical Trials: The Power of Continuous Feedback from Medical Reports
- Factors Influencing Brain Drain: Perspectives from a Medical School in Turkey