Role of HDL and Scavenger Receptor Class B Type I (SR-BI) Expression in LNCaP and PC-3 Prostate Cancer Cell Lines. Growth Patterns in the Chick Chorioallantoic Membrane Model
Cholesterol metabolism is a key pathway affecting androgen synthesis and prostate cancer cell proliferation. Major receptors and transporters involved in cellular cholesterol homeostasis and cholesterol fluxes are ATP-binding cassette protein A1 (ABCA1), LDL receptor (LDLR) and the HDL receptor scavenger receptor class B type I (SR-BI). Especially SR-BI deserves more and more attention because it has been shown to be overexpressed by the majority of malignant tumours, promoting their proliferation and metastasis. PC-3 and LNCaP are two of the commonly used cell lines for in vitro prostate cancer studies. A comparative overview of the expression of receptors involved in cholesterol uptake and their activities in these two cell lines was missing; therefore we reviewed the current knowledge on this topic. We investigated the expression of SR-BI in these two cell lines and the uptake of labelled HDL. In order to study and compare tumour development, we assessed the growth of both cell lines in the chick chorioallantoic membrane (CAM) model. Although microtumours successfully developed, size increase was limited as confirmed by the analysis of the proliferation marker Ki67. Interestingly prostate-specific antigen (PSA) secreted by LNCaP tumours could be determined in the chick embryo blood. SR-BI is a promising therapeutic target either by its direct inhibition or more importantly as an agent for drug delivery.
Introduction
Cholesterol Metabolism
Cholesterol is essential in cell metabolism as the most abundant lipid component of cell membrane, influencing its fluidity and consequently cellular signalling [1]. Cells obtain cholesterol by two pathways: endogenously by de novo synthesis through the mevalonate pathway, in which the rate-limiting enzyme 3-hydroxy-3-methylglutaryl-coenzyme A reductase (HMGCR) is the target of statins [2]. Exogenously, circulating lipoproteins provide cholesterol to cells mainly through LDL (via LDLR) and HDL (via SR-BI). Intracellularly, especially in steroidogenic tissues, cholesterol is stored in lipid droplets as cholesteryl ester. Cholesterol is a precursor of bile acids and more importantly of hormones for steroid- dependent tissues. Prostate cancer cells use cholesterol for proliferation and steroidogenesis [2]. Diverse mechanisms are impaired in prostate cancer cells leading to cholesterol enrichment:
- Higher cholesterol uptake via SR-BI
- Cholesterol accumulation in cells by ABCA1 downregulation
- Enhance uptake of LDL
- Altered enzymes in cholesterol synthesis particularly HMGCR
- At the transcriptional level by two key regulators, sterol regulatory element-binding protein (SREBP) and liver X receptor (LXR)
- Oxysterols imbalance [3, 4]
- Androgen receptor (AR) pathway activation [5, 6, 7]
- Loss of the PTEN tumor suppressor and upregulation of the PI3K/AKT pathway [8]
Consequently, all these dysregulated elements contribute to altered plasma membrane cholesterol content and cholesterol accumulation in cancer cells. Originally, these pathways are closely interrelated to maintain cholesterol homeostasis. Cholesterol in prostate cancer has been the topic of previous reports [9, 10].
Cholesterol is likely involved in the development of advanced prostate cancer as a substrate in androgen synthesis and in addition, by stimulating signalling pathways by stabilising lipid raft structures [10, 11]. Studies showed that low plasma cholesterol levels and statin use were associated with a decreased risk of prostate cancer development and progression [9, 12].
Cholesterol Metabolism in LNCaP and PC-3 Prostate Cancer Cell Lines
LNCaP cell line is androgen-dependent, expresses AR and is described as poorly tumorigenic; PC-3 cell line is androgen-insensitive and highly metastatic [14, 15]. Several studies addressed these two cell lines because there are representative of the two most significant prostate cancer clinical scenarios; LNCaP cells preserving more prostate specific characteristics than PC-3 [16, 17, 18, 19].
Cholesterol content was found to be similar in both cell lines in culture medium according to Krycer et al. (with little esterified cholesterol), whereas Sekine et al. found more total cholesterol in LNCaP than PC-3 after 24 h starving [5, 20]. Yue et al. assessed about 20% cholesterol ester in LNCaP versus 90% in PC-3 [8]. Obviously, cell culture conditions, experimental settings and cholesterol determination could explain these discrepancies.
Interestingly when comparing normal and tumour tissues, total cholesterol was similar, but with higher oxysterol concentrations in prostate cancer samples [3].
SR-BI
SR-BI is primarily expressed on hepatocytes and steroidogenic tissues. SR-BI is overexpressed in many tumour types, playing a role in cancer progression [21, 22]. More and more studies show increased SR-BI expression in human prostate cancer specimens [3, 12, 23, 24]. Through SR- BI, HDL delivers cholesterol to cells. In prostate cancer cells, HDL and SR-BI receptor were involved in higher cholesterol uptake, aberrant cholesterol metabolism favouring cancer progression [3, 9, 23]. In mouse models, SR-BI and HDL promoted prostate cancer tumour growth [12, 24, 26].
Although SR-BI has been published to be similarly expressed in LNCaP and PC-3 cell lines [6, 20, 27], we found that SR-BI was clearly more expressed in LNCaP than PC-3 (Figure 1A).
Of particular interest is the cell proliferation inhibition by blocking SR-BI chemically or by knock down. The SR-BI inhibitor ITX5061 blocked the uptake of HDL and proliferation of LNCaP [26]. Furthermore SR-BI silencing prevented prostate-specific antigen (PSA) secretion and cell proliferation; this PSA decrease reflected the impact of SR-BI on AR activity [24, 28].
We investigated the binding of HDL to prostate cancer cell lines; cells were pre-treated with HDL and the SR-BI inhibitor BLT-1 followed by incubation with fluorescent- labelled HDL (Figure 1B). In LNCaP, BLT-1 decreased HDL uptake by 39% but only 19% in PC-3. As expected, native HDL potently competed with fluorescent HDL in both cell lines.
(a) PC-3 LNCaP
PC-3
SR-BI/GAPDH band density
SR-BI
$$ \therefore \mathrm {即} \mathrm {当} x = 0 \mathrm {时} \mathrm {有} \mathrm {点} (0, 0) $$ GAPDH SR-BI/GAPDH band density 0.031 0.036 0.309 (b) Dil-HDL uptake Control 10 µM BLT-1 100 µg/ml HDL
Figure 1: A Representative immunoblot of SR-BI expression in LNCaP and PC-3. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used for normalisation. Band density is presented as a bar graph at the right of the blot illustration. B Fluorescent-labelled HDL binding to LNCaP and PC-3. Cells were pre-incubated with BLT-1 and HDL before incubation with DiI-HDL (40µg/ml). Fluorescence in LNCaP and PC-3 were determined by flow cytometry.
LDL Receptor
LDL is the major lipoprotein to deliver cholesterol to the cells [9]. LDLR was reported to be upregulated in PC-3 compared to LNCaP leading to a most prominent SR-BI expression PC-3 LNCaP
0.0 LDL uptake in PC-3 cells compared to LNCaP [8, 16, 17, 29]. This finding was consistent with the higher activity of SREBP2 in PC-3; this transcription factor is a cellular lipid sensor that controls LDLR expression (Figure 2) [9, 17].
Cholesterol accumulation

Figure 2: A schematic representation of cholesterol pathways in prostate cancer cells. Cholesterol uptake is mediated mainly by LDL via LDLR and partly by HDL via SR-BI. ABCA1 pumps cholesterol out. Cholesterol can be produced de novo via the mevalonate pathway with its rate-limiting HMGCR enzyme, target of statins. Cholesterol can be used for proliferation, converted in oxysterols and further to hormones like androgens and stored in lipid droplets. The transcription factors SREBP2 and LXR regulate cholesterol homeostasis. CYP17A1 is a rate-limiting enzyme in androgen synthesis from cholesterol. AR signalling plays a crucial role in prostatic diseases by inducing proliferation. In red pathways leading to cholesterol accumulation favouring cancer development, in green pathways decreasing intracellular cholesterol, restraining cancer progression. Adapted from [9, 13].
ABCA1 and ABCG1
ABCA1 plays a major role in cellular cholesterol homeostasis by exporting cellular cholesterol and phospholipids to plasma lipoproteins such as native HDL [30, 31]. ABCA1 protein levels were found to be decreased in LNCaP compared to normal prostate epithelial cells and other prostate cancer cell lines; consequently cellular cholesterol content was increased and efflux reduced [5, 20, 32]. However, Ruscica et al. reported contradicting data [27]. Indeed, they found similar ABCA1 levels in LNCaP and PC-3 which could be explained by differences in the experimental settings. Furthermore ABCA1 expression is regulated post- transcriptional [30]. Lee et al. described silencing of ABCA1 gene in LNCaP leading to cholesterol accumulation and confirmed this observation in prostate cancer specimens [11].
ABCG1 is another cholesterol transporter expressed by prostate cancer cell lines [11, 20, 27]. Its implication in tumour progression deserves further clarifications but its increased expression in prostate tumours has been reported [12].
SREBP2, LXR and AR
The nuclear hormone receptors SREBP and LXR are key regulators of cholesterol metabolism, SREBP2 favours cholesterol uptake and synthesis, whereas LXR prevents cholesterol accumulation [9]. LDLR and HMGCR are target genes of SREBP2; ABCA1 is controlled by LXR [9, 30, 31]. SREBP2 and LXR are positive regulators of SR-BI [21]. However its regulation seems complex and not specifically described in prostate cancer.
Androgens through their binding to AR transcriptionally regulate multitude pathways in prostate cells like growth, survival and physiology, including the PSA marker [6, 33, 34]. AR activates SREBP2 leading to cellular cholesterol accumulation and ultimately to proliferation (Figure 2). AR and LXR are interconnected; LXR controls AR activity and vice versa [9, 33]. Interestingly, Celhay et al. found modified oxysterol content in tumours compared to normal tissues [3]. These oxysterols regulate SREBP2 and LXR activity, which ultimately lead to cholesterol dysregulation in cancer cells through all these interconnected pathways [9].
LXR expression was found to be higher in LNCaP compared to PC-3 and vice versa [32, 33, 34, 35]. Overall LXR activity and AR complex interaction has been described [9, 33].
The Chick Chorioallantoic Membrane (CAM) Model for Prostate Cancer
The CAM model is a well-established in vivo assay to study tumour growth and to evaluate efficacy of anticancer drugs [36, 37]. It is an attractive alternative to animal experiments. Although prostate cancer has been described in the CAM model, tumour growth assays have not been reported so far [38, 39].
After inoculation of LNCaP and PC-3 cells on the CAM, microtumours successfully developed. Tumour growth was monitored from day 2 to 6; from day 2, the flat cell layers got vascularised by the CAM vessels, compacted and by day 6 the tumours were mainly located under the CAM. Neither weight nor volume increased during this phase.
PC-3 and LNCaP tumours were excised from the CAM from day 2 to 6 and processed for immunohistochemistry and stained for the proliferation marker Ki67. Visually LNCaP and PC-3 tumour structures differed (Figure 3A). LNCaP presented clusters of cells with no lumen surrounded by extracellular matrix essentially composed of collagen as suggested by Masson’s trichrome staining. PC-3 tumours presented a trabecular structure composed of more dedifferentiated malignant cells and less extracellular matrix.
Ki67 immunostaining revealed a drop of 5.6-fold in LNCaP from day 2 to 6 (Figure 3B). As LNCaP cells secreted PSA, it could be determined in the blood of the chick embryo; PSA decreased 138-fold (from 8.28 µg/ml by day 2 to 0.06 µg/ml by day 6), (Figure 3C).
LNCaP and PC-3 cell lines had both the capacity to efficiently implant in the CAM and form vascularised tumours, but no further volume gain could be observed unlike other tumour types [40, 41]. ‘Small nodules were evident at the site of inoculation of the cells, but they did not increase in size’ reported Kim et al. about prostate cell lines implanted in the CAM [39]. By calculating tumour volume and unlike their in vitro assays, Reuter et al. were unable to demonstrate an antiproliferative effect of their intervention in LNCaP tumours implanted in the CAM [25].
Fang et al. were able to monitor the weight of PC-3 tumours after 9 days of growth in the CAM [42]. But without special authorization, egg development had to be stopped at embryonic development day (EDD) 14, two third of the embryo development before hatching, allowing 6 days of tumour growth in the CAM.
Nonetheless, Ki67 drastically decreased in LNCAP tumours with a simultaneous drop of PSA blood levels. Estrada et al. reported Ki67 expression in PC-3 and LNCaP in the CAM [43]. These authors found similar expression of Ki67 at day 6 of tumour development for both cell lines whereas we observed 9 times lower levels in LNCaP versus PC-3 (Figure 3B).
![Figure 2: A schematic representation of cholesterol pathways in prostate cancer cells. Cholesterol uptake is mediated mainly by LDL via LDLR and partly by HDL via SR-BI. ABCA1 pumps cholesterol out. Cholesterol can be produced de novo via the mevalonate pathway with its rate-limiting HMGCR enzyme, target of statins. Cholesterol can be used for proliferation, converted in oxysterols and further to hormones like androgens and stored in lipid droplets. The transcription factors SREBP2 and LXR regulate cholesterol homeostasis. CYP17A1 is a rate-limiting enzyme in androgen synthesis from cholesterol. AR signalling plays a crucial role in prostatic diseases by inducing proliferation. In red pathways leading to cholesterol accumulation favouring cancer development, in green pathways decreasing intracellular cholesterol, restraining cancer progression. Adapted from [9,13].](/fulltextimages/7601/fig_2.jpeg)
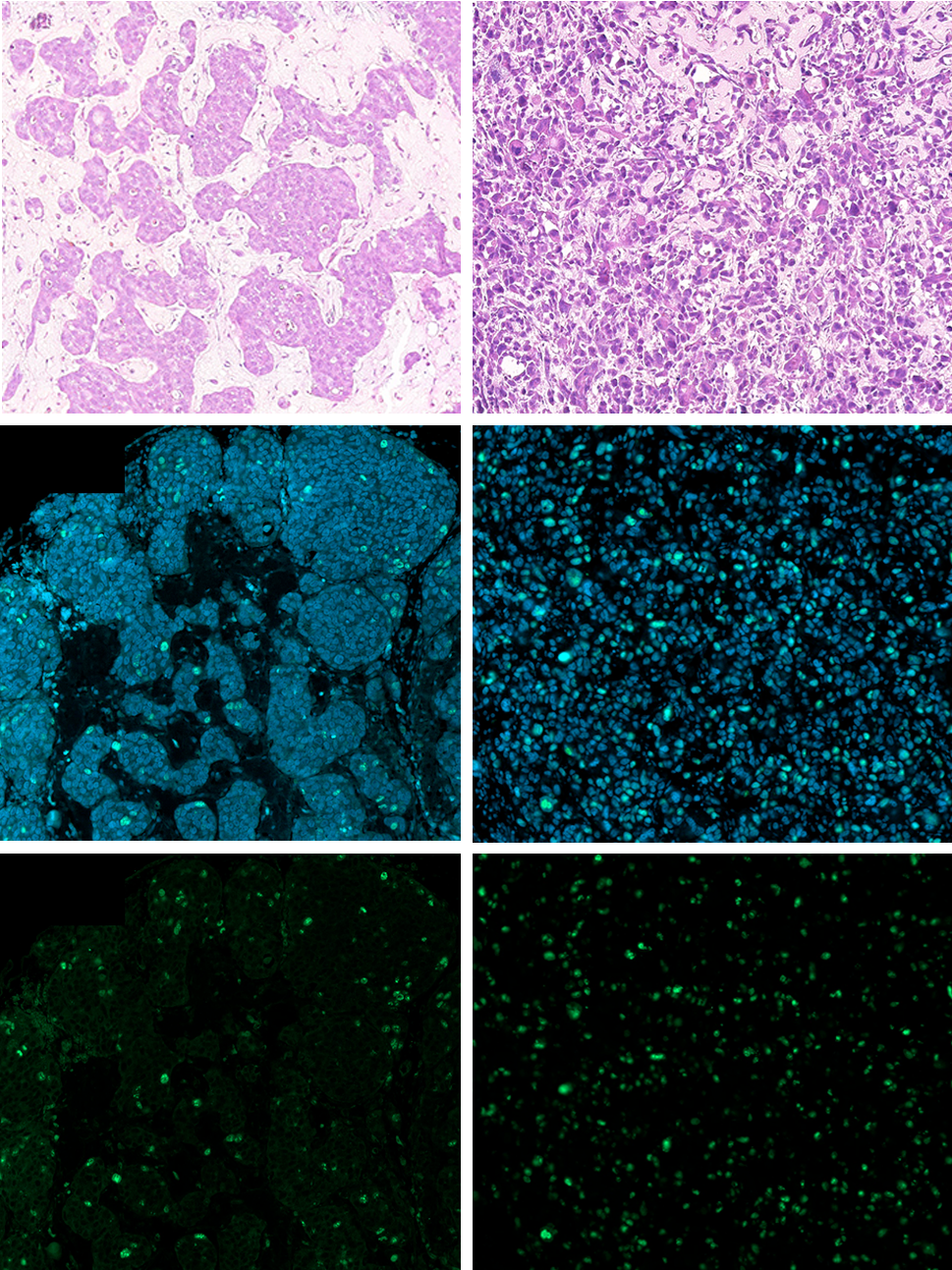
Figure 3: Ki67 immunostaining in LNCaP and PC-3 tumours grown in the CAM. A Representative HE, Ki67 and DAPI staining. B Relative quantification of Ki67 positive nuclei reported to DAPI stained nuclei (n = 4 to 6 tumours per conditions, *p < 0.05, ** p < 0.01, Student’s t-test). C PSA secreted by LNCaP tumours in chick plasma. TDD: tumour development day.
| Target | LNCaP and PC-3 comparison | References |
|---|---|---|
| SR-BI | Similar or increased in LNCaP | [20,27], our data |
| LDLR | Increased in PC-3 | [8,16,17,29] |
| ABCA1 | Increased in PC-3 | [5,20,32] |
| ABCG1 | Increased in PC-3 | [27,32] |
| LXR | No consensus | [32,34] |
| SREBP2 | No consensus | [17,35] |
| Cholesterol content | Depending on cells and experimental conditions | [5,8,20] |
Table 1: Comparison of receptor expression and transcription factor activity in LNCaP and PC-3 cell lines.
Discussion
Cellular cholesterol concentrations are critical for proper function of plasma membrane proteins. Cholesterol reduction and enrichment lead to the alteration of their activity [1]. Cholesterol homeostasis to support tumour growth and androgen independency is disrupted in prostate cancer cells. Cholesterol receptor and transporter activities (SR-BI, LDLR and ABCA1) are altered favouring cellular cholesterol accumulation. We focussed our study on SR-BI as growing evidence has shown an overexpression of this receptor in malignant tumours [21, 22]. This observation was confirmed by the analysis of prostate cancer tissue specimens [3, 12, 23, 24].
In animal models, SR-BI inhibitors (ITX5061, BLT-1) and SR-BI knock down have been shown to reduce progression of prostate tumours [12, 24, 26]. Another promising approach is to use SR-BI overexpression in tumours as a gateway for drug delivery. Indeed SR-BI is a permissive receptor, which binds a large variety of ligands [22]. Nanoparticles and reconstitute HDL drug delivery technology are under investigation [21, 44, 45]. For instance, HDL could be used to deliver antioxidants to prostate cancer cells [27].
LNCaP and PC-3 prostate cancer cell lines are representative of early and late stages of prostate cancer respectively; LNCaP is androgen-responsive and PC-3 androgen-independent. By gathering information in literature on the major cholesterol transporters (SR-BI, LDLR and ABCA1) in these two cell lines, no clear picture could be drawn (Table 1). Discordant experimental results using these cell lines are recurrent as mentioned above. Extra care should be taken when culturing the weakly adherent LNCaP cell line as mentioned by Sobel et al. [14]. Furthermore, low-passage and high-passage LNCaP were used as models for prostate cancer progression, showing the instability of these cells [8]. Prostate cancer cell lines require rigorous culture conditions to avoid inconsistent results. Obviously studying cholesterol metabolism in vitro in cells in different experimental settings (for instance in culture medium in presence or absence of serum) led to different outcomes. This point should be considered when interpreting the results. Thus we found higher levels of SR-BI expression in LNCaP than PC-3, which translated in a stronger inhibition by BLT-1 of HDL uptake in LNCaP compared to PC-3. This increased SR-BI expression in LNCaP could come from low passages and culture handling as a non-adherent cell line. On the other hand PC-3 appears to promote the LDL pathway, as measured by the increased uptake of fluorescent LDL in PC-3 compared to LNCaP [8].
In order to study prostate tumour growth, we used the CAM model. LNCaP and PC-3 were implanted, though their tumour growths were limited. Importantly we showed that in the LNCaP model, PSA levels could be measured in the chick embryo blood. The PSA decrease over the observed period would come from the concomitant cell proliferation arrest and blood volume increase during the chick embryo development [46].
Kelley et al. studied the lipoprotein composition during the chick embryo development [47]. Interestingly chick embryo plasma was essentially composed of VLDL and LDL, HDL remaining low until hatching. The lipoprotein composition of the chick embryo plasma could alter the growth of the prostate tumours in the CAM, especially LNCaP. Of course tumorigenicity and microenvironment are essential parameters for tumour progression, which might not be gathered for prostate cancer cell line development in the CAM model.
Although cholesterol and steroid hormone metabolism has been shown to play a major role in prostate cancer cell proliferation in preclinical models, cholesterol synthesis, storage and uptake pathways have not been thoroughly and successfully investigated in the clinial setting. Obviously additional actors come into play when considering cholesterol metabolism in prostate cancer and deserve further investigations.
Materials and Methods
Cell Lines and Cell Culture
Cell lines were purchased from LGC Standards, culture media and supplements were from Gibco (Thermo Fischer Scientific). PC-3 (ATCC CRL-1435) were maintained in Ham’s F-12K medium, LNCaP clone FGC (ATCC CRL-1740) in RPMI. Culture media were supplemented with 10% FBS and 50 units/ml penicillin and 50 µg/ml streptomycin. LNCaP were weakly adherent and consequently handled like non- adherent cells, although they still needed to be trypsinised to dissociate aggregates.
CAM Tumour Grafts
Fertilised hen eggs (animal facility, Geneva University) were incubated at 37.7°C with ≤ 47% humidity (Incubator MG140/200 rural, Fiem). From EDD 1 to 4 eggs were placed the wide apex up and sporadically rotated. On EDD 4 eggs were turned upside down and a small hole was made in the narrow apex. At EDD 8 the hole was enlarged and the CAM gently damaged with a cotton bud near the junction of blood vessels. 1-2*106 cells in 60 µl were applied in a cryovial o-ring placed on the injured membrane. The hole in the eggshell was sealed with a double layer of Parafilm and eggs put back into the incubator.
Cell were prepared in culture medium and placed on ice. They were mixed with half a volume of cold Matrigel (Corning) before seeding on the CAM.
From EDD 10 to 14 tumours were cut out of the CAM. Blood was collected in heparin, centrifuged and plasma was frozen.
Immunohistochemistry
Tumours were briefly washed in PBS and fixed in 10% buffered formaldehyde for 24 h. Samples were dehydrated and paraffin embedded.
Immunohistochemical staining was performed on 5 µm- thick sections after deparaffinisation in Neo Clear (Merck) and rehydration through a graded series of ethanol. Antigen retrieval was performed in Tris-borate-EDTA buffer (pH 8) for 20 min at 100°C in a water bath following cooling to allow antigen site to reform after exposure to high temperature. After a permeabilisation step in TBS 10% Triton for 10 min and a blocking step in 5% BSA in TBS 10% Triton for 20 min, the sections were incubated overnight at 4°C with the anti- Ki67 primary antibody (Cell Signaling 9027) diluted 500 times in antibody signal enhancer [48]. The sections were washed with PBS 0.2% Tween and incubated for 1 h at RT with the secondary antibody (anti-rabbit IgG, Alexa fluor 488 conjugate, Life technologies A21206, Thermo Fischer Scientific) 1/1000 dilution and 250 ng/ml DAPI (AppliChem). Slides were washed and mounted in fluorescence mounting medium (Dako, Agilent) and images acquired with Axio Scan. Z1 slide scanner (Zeiss). Quantification was performed with open source Cell Profiler software, counting Ki-67 positive cells divided by the number of DAPI-stained nuclei.
PSA measurement
Human PSA in chick plasma was determined by the medical lab of the Geneva University Hospitals (Cobas 8000 analyser, Roche).
Immunoblotting
Western blot were made as described [49]. Primary antibody: anti-SR-BI 1/5000 dilution (Novus NB400- 101), anti-GAPDH 1/500 dilution (Millipore MAB374) and secondary antibodies anti-rabbit immunoglobulins/HRP (Dako P0448, Agilent) and anti-mouse immunoglobulins/ HRP (Dako P0447, Agilent) both 1/10000 dilution.
Lipoprotein Isolation and Labelling
Blood was harvested in EDTA blood collection tubes (Vacutainer, BD), centrifuged at 3500 rpm for 15 min at 4°C. Plasma was transferred to centrifuge tubes (OptiSeal, Beckman Coulter) and overlaid with NaBr density 1.006 g/ cm3 and ultracentrifuged at 50000 rpm for 16 h at 20°C in a 60 Ti rotor (Beckman Coulter). Top fraction containing chylomicron and VLDL was discarded. Density of bottom fraction was raised to 1.063 g/cm3 with solid NaBr, transferred to centrifuge tubes, overlaid with NaBr d = 1.063 g/cm3 and ultracentrifuged at 50000 rpm for 18 h at 20°C. Top fraction containing LDL was collected. Bottom fraction was processed at density 1.21 g/cm3 as described for d = 1.063 g/cm3 with ultracentrifugation extended to 25 h. Top fraction contained HDL and bottom fraction lipoprotein- deficient serum (LPDS). LDL, HDL and LPDS samples were dialysed against PBS supplemented with 100 µM EDTA. Fraction protein content was determined (Protein assay 500-0006, Bio-Rad).
HDL were labelled as described [50]. 1 mg HDL were mixed with 1 ml LPDS and 1 ml PBS supplemented with 100 µM EDTA, while vortexing 50 µl 1,1′-dioctadecyl-3,3,3′,3′- tetramethylindocarbocyanine perchlorate (DiI, 3 mg/ml in DMSO, Sigma, Merk) was added. Solution was incubated overnight at 37°C and ultracentrifuged as described above at density 1.21 g/cm3 at 50000 rpm for 25 h at 20°C. Top fraction was dialysed, concentrated (Vivaspin columns, 10000 MWCO, Satorius) and protein concentration determined.
Lipoprotein Uptake Assay
PC-3 and LNCaP were detached with Accutase (Gibco, Thermo Fischer Scientific). Cells were pre-incubated for 50 min at 37°C with 10 µM BLT-1 (EMD Millipore), 100 µg/ml unlabelled HDL in culture medium supplement with 0.2% BSA. Then 40 µg/ml HDL was added for 4 h at 37°C. Cells were washed with PBS supplemented with 0.2% BSA and counterstained with 3 µM DAPI. Fluorescence was acquired by flow cytometry (Attune NxT, Life Technologies). 30000 events were analysed.
Data Analysis
Data plotting and statistical analysis were done using Prism GraphPad 6.
Acknowledgements
We are grateful to Laurence Zulianello for designing the figures.
These investigations were supported by Fundació Privada CELLEX and Fondation Nutrition 2000 Plus.
References
-
Meza U, Delgado-Ramírez M, Romero-Méndez C, Sánchez-Armass S, Rodríguez-Menchaca AA (2020) Functional marriage in plasma membrane: Critical cholesterol level-optimal protein activity. Br J Pharmacol 177(11): 2456-2465.
-
Kim JH, Cox ME, Wasan KM (2014) Effect of simvastatin on castration-resistant prostate cancer cells. Lipids Health Dis 13: 56.
-
Celhay O, Bousset L, Guy L, Kemeny JL, Leoni V, et al. (2019) Individual comparison of cholesterol metabolism in normal and tumour areas in radical prostatectomy specimens from patients with prostate cancer: Results of the chomecap study. Eur Urol Oncol 2(2): 198-206.
-
Dambal S, Alfaqih M, Sanders S, Maravil la E, Ramirez Torres A, et al. (2020) 27 hydroxycholes terol impairs plasma membrane lipid raft signaling as evi denced by inhibition of il6-jak-stat3 signaling in prostate cancer cells. Mol Cancer Res 18(5): 671-684.
-
Krycer JR, Brown AJ (2011) Cross-talk between the androgen receptor and the liver x receptor: Implications for cholesterol homeostasis. J Biol Chem 286(23): 20637-20647.
-
Tousignant KD, Rockstroh A, Taherian Fard A, Lehman ML, Wang C, et al. (2019) Lipid uptake is an androgen- enhanced lipid supply pathway associated with prostate cancer disease progression and bone metastasis. Mol Cancer Res 17(5): 1166-1179.
-
Han W, Gao S, Barrett D, Ahmed M, Han D, et al. (2018) Reactivation of androgen receptor-regulated lipid biosynthesis drives the progression of castration- resistant prostate cancer. Oncogene 37(6): 710-721.
-
Yue S, Li J, Lee SY, Lee HJ, Shao T, et al. (2014) Cholesteryl ester accumulation induced by pten loss and pi3k/ akt activation underlies human prostate cancer aggressiveness. Cell Metab 19(3): 393-406.
-
Krycer JR, Brown AJ (2013) Cholesterol accumulation in prostate cancer: A classic observation from a modern perspective. Biochim Biophys Acta 1835(2): 219-229.
-
Hryniewicz-Jankowska A, Augoff K, Sikorski AF (2019) Highlight article: The role of cholesterol and cholesterol- driven membrane raft domains in prostate cancer. Exp Biol Med (Maywood) 244(13): 1053-1061.
-
Lee BH, Taylor MG, Robinet P, Smith JD, Schweitzer J, et al. (2013) Dysregulation of cholesterol homeostasis in human prostate cancer through loss of abca1. Cancer Res 73(3): 1211-1218.
-
Traughber CA, Opoku E, Brubaker G, Major J, Lu H, et al. (2020) Uptake of high-density lipoprotein by scavenger receptor class b type 1 is associated with prostate cancer proliferation and tumor progression in mice. J Biol Chem 295(24): 8252-8261.
-
Stopsack KH, Gerke TA, Andren O, Andersson SO, Giovannucci EL, et al. (2017) Cholesterol uptake and regulation in high-grade and lethal prostate cancers. Carcinogenesis 38(8): 806-811.
-
Sobel RE, Sadar MD (2005) Cell lines used in prostate cancer research: A compendium of old and new lines-- part 1. J Urol 173(2): 342-359.
-
van Bokhoven A, Varella-Garcia M, Korch C, Johannes WU, Smith EE, et al. (2003) Molecular characterization of human prostate carcinoma cell lines. Prostate 57(3): 205-225.
-
Locke JA, Wasan KM, Nelson CC, Guns ES, Leon CG (2008) Androgen-mediated cholesterol metabolism in lncap and pc-3 cell lines is regulated through two different isoforms of acyl-coenzyme a:Cholesterol acyltransferase (acat). Prostate 68(1): 20-33.
-
Krycer JR, Kristiana I, Brown AJ (2009) Cholesterol homeostasis in two commonly used human prostate cancer cell-lines, lncap and pc-3. PLoS One 4(12): e8496.
-
Singh AN, Sharma N (2020) Quantitative swath-based proteomic profiling for identification of mechanism- driven diagnostic biomarkers conferring in the progression of metastatic prostate cancer. Front Oncol 10: 493.
-
Dozmorov MG, Hurst RE, Culkin DJ, Kropp BP, Frank MB, et al. (2009) Unique patterns of molecular profiling between human prostate cancer lncap and pc-3 cells. Prostate 69(10): 1077-1090.
-
Sekine Y, Demosky SJ, Stonik JA, Furuya Y, Koike H, (2010) High-density lipoprotein induces proliferation and migration of human prostate androgen-independent cancer cells by an abca1-dependent mechanism. Mol Cancer Res 8(9): 1284-1294.
-
Gutierrez-Pajares JL, Ben Hassen C, Chevalier S, Frank PG (2016) Sr-bi: Linking cholesterol and lipoprotein metabolism with breast and prostate cancer. Front Pharmacol 7: 338.
-
Mooberry LK, Sabnis NA, Panchoo M, Nagarajan B, Lacko AG (2016) Targeting the sr-b1 receptor as a gateway for cancer therapy and imaging. Front Pharmacol 7: 466.
-
Schörghofer D, Kinslechner K, Preitschopf A, Schütz B, Röhrl C, et al. (2015) The hdl receptor sr-bi is associated with human prostate cancer progression and plays a possible role in establishing androgen independence. Reprod Biol Endocrinol 13: 88.
-
Gordon JA, Noble JW, Midha A, Derakhshan F, Wang G, et al. (2019) Upregulation of scavenger receptor b1 is required for steroidogenic and nonsteroidogenic cholesterol metabolism in prostate cancer. Cancer Res 79(13): 3320-3331.
-
Reuter A, Sckell A, Brandenburg LO, Burchardt M, Kramer A, et al. (2019) Overexpression of microrna-1 in prostate cancer cells modulates the blood vessel system of an in vivo hen’s egg test-chorioallantoic membrane model. In Vivo 33(1): 41-46.
-
Patel R, Fleming J, Mui E, Loveridge C, Repiscak P, et al. (2018) Sprouty2 loss-induced il6 drives castration- resistant prostate cancer through scavenger receptor b1. EMBO Mol Med 10(4): e8347.
-
Ruscica M, Botta M, Ferri N, Giorgio E, Macchi C, et al. (2018) High density lipoproteins inhibit oxidative stress-induced prostate cancer cell proliferation. Sci Rep 8(1): 2236.
-
Twiddy AL, Cox ME, Wasan KM (2012) Knockdown of scavenger receptor class b type i reduces prostate specific antigen secretion and viability of prostate cancer cells. Prostate 72(9): 955-965.
-
Furuya Y, Sekine Y, Kato H, Miyazawa Y, Koike H, et al. (2016) Low-density lipoprotein receptors play an important role in the inhibition of prostate cancer cell proliferation by statins. Prostate Int 4(2): 56-60.
-
Phillips MC (2018) Is abca1 a lipid transfer protein? J Lipid Res 59(5): 749-763.
-
Xiong T, Xu G, Huang XL, Lu KQ, Xie WQ, et al. (2018) Atp- binding cassette transporter a1: A promising therapy target for prostate cancer. Mol Clin Oncol 8(1): 9-14.
-
Trasino SE, Kim YS, Wang TT (2009) Ligand, receptor, and cell type-dependent regulation of abca1 and abcg1 mrna in prostate cancer epithelial cells. Mol Cancer Ther 8(7): 1934-1945.
-
Cariello M, Ducheix S, Maqdasy S, Baron S, Moschetta A, et al. (2018) Lxrs, shp, and fxr in prostate cancer: Enemies or menage a quatre with ar? Nuclear receptor signaling 15: 1550762918801070.
-
Tsui KH, Chung LC, Feng TH, Lee TY, Chang PL, et al. (2015) Divergent effect of liver x receptor agonists on prostate-specific antigen expression is dependent on androgen receptor in prostate carcinoma cells. Prostate 75(6): 603-615.
-
Li X, Wu JB, Li Q, Shigemura K, Chung LW, et al. (2016) Srebp-2 promotes stem cell-like properties and metastasis by transcriptional activation of c-myc in prostate cancer. Oncotarget 7(11): 12869-12884.
-
Ribatti D (2017) The chick embryo chorioallantoic membrane (cam) assay. Reprod Toxicol 70: 97-101.
-
DeBord LC, Pathak RR, Villaneuva M, Liu HC, Harrington DA, et al. (2018) The chick chorioallantoic membrane (cam) as a versatile patient-derived xenograft (pdx) platform for precision medicine and preclinical research. Am J Cancer Res 8(8): 1642-1660.
-
Kunzi-Rapp K, Genze F, Kufer R, Reich E, Hautmann RE, et al. (2001) Chorioallantoic membrane assay: Vascularized 3-dimensional cell culture system for human prostate cancer cells as an animal substitute model. J Urol 166(4): 1502-1507.
-
Kim J, Yu W, Kovalski K, Ossowski L (1998) Requirement for specific proteases in cancer cell intravasation as revealed by a novel semiquantitative pcr-based assay. Cell 94(3): 353-362.
-
Kleibeuker EA, Ten Hooven MA, Castricum KC, Honeywell R, Griffioen AW, et al. (2015) Optimal treatment scheduling of ionizing radiation and sunitinib improves the antitumor activity and allows dose reduction. Cancer Med 4(7): 1003-1015.
-
Hagedorn M, Javerzat S, Gilges D, Meyre A, de Lafarge B, et al. (2005) Accessing key steps of human tumor progression in vivo by using an avian embryo model. Proc Natl Acad Sci U S A 102(5): 1643-1648.
-
Fang J, Ding M, Yang L, Liu LZ, Jiang BH (2007) Pi3k/pten/ akt signaling regulates prostate tumor angiogenesis. Cell Signal 19(12): 2487-2497.
-
Estrada AC, Syrovets T, Pitterle K, Lunov O, Buchele B, et al. (2010) Tirucallic acids are novel pleckstrin homology domain-dependent akt inhibitors inducing apoptosis in prostate cancer cells. Mol Pharmacol 77(3): 378-387.
-
Raut S, Mooberry L, Sabnis N, Garud A, Dossou AS, et al. (2018) Reconstituted hdl: Drug delivery platform for overcoming biological barriers to cancer therapy. Front Pharmacol 9: 1154.
-
Niora M, Pedersbæk D, Münter R, Weywadt MFV, Farhangibarooji Y, et al. (2020) Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega 5(33): 21162-21171.
-
Mueller C, Burggren W, Tazawa H (2015) The physiology of the avian embryo. In: Sturkie’s avian physiology. Academic Press, San Diego, USA, pp: 739-766.
-
Kelley J, Schjeide O, Schjeide S, Milius R, Alaupovic P (1980) Quantification of the major apolipoproteins in chicken and turkey serum during embryonic development. Comparative Biochemistry and Physiology--Part B: Biochemistry and Molecular Biology 65(2): 239-242.
-
Rosas-Arellano A, Villalobos-Gonzalez JB, Palma- Tirado L, Beltran FA, Carabez-Trejo A, et al. (2016) A simple solution for antibody signal enhancement in immunofluorescence and triple immunogold assays. Histochem Cell Biol 146(4): 421-430.
-
Colin DJ, Cottet-Dumoulin D, Faivre A, Germain S, Triponez F, et al. (2018) Experimental model of human malignant mesothelioma in athymic mice. International journal of molecular sciences 19(7): 1881.
-
Pitas RE, Innerarity TL, Weinstein JN, Mahley RW (1981) Acetoacetylated lipoproteins used to distinguish fibroblasts from macrophages in vitro by fluorescence microscopy. Arteriosclerosis 1(3): 177-185.
- Cancer Diagnosis from RNA Sequence of Blood Cells by Using AI
- Field Cancerization in Oral Cavity, Case Report and Review of Literature. Oncologic Program Salud Integral Hospital, Managua, Nicaragua
- Identification of B Lymphocytes in Cancer Patient’s Blood
- A Case Report of a Breast Cancer Patient Developing Pneumonitis as a Result of Abemaciclib Therapy
- Immune Checkpoint Therapeutics for Today’s Fight and Beyond
- The Amalgamated Sophomore-Gonadoblastoma