Investigation of Proteomic Responses and Anticancer Activities of an Isolated Strain of Cordyceps sinensis Under Different Nutritional Fermentation Conditions
Cordyceps sinensis is a fungal parasite on the larvae of Lepidoptera and has been used as a tonic herb in the traditional Chinese medicines. The fruit bodies of C. sinensis are limited because of the difficulty in cultivation. The mycelia produced by the fermentation have already become the best substitute for C. sinensis. The fungus strain called G308-1 strain used in this study was isolated from the fruit bodies of C. sinensis in Yunan Province of People Republic of China. From the comparisons of rDNA sequences against Gene Bank, the G308-1 strain was identified as Paecilomyces farinosus. In this study, the proteomic profiles of the G308-1 strain grown in the different liquid media containing glucose, sucrose, maltose or lactose as the sole carbon source were established using two-dimensional gel electrophoreses. The proteomes of the mycelia in the glucose media were compared respectively to those in the other three media. The comparative results showed that two, five and seventeen protein spots were repressed respectively in the mycelia in the sucrose, maltose and lactose media while six, three and eight protein spots were induced respectively. In order to explore the correlation between the proteomes and the anticancer activities of the mycelia, the water or ethanol extracts of the G308-1 strain cultivated in various media were examined for their abilities to suppress the HepG2, Du145, Hela and MCF-7 cells. The anticancer results demonstrated that the water extracts showed the selectivity to Hela and Du 145 cells and the high concentrations of the water extracts made from the mycelia in the lactose media had better suppressive effects on the HepG2 cells. Essentially the ethanol extracts prepared from the mycelia in the lactose media showed much better inhibitory activities to each cell line than those from the other three media. The above results suggested the anticancer results might vary with the proteome profile changes which probably resulted in the production of a different array of bio-products in the mycelia under the different nutritional conditions.
Introduction
Cordyceps sinensis of the clavipitaceae family is a medicinal fungus of traditional Chinese medicine. It possesses many pharmacological activities, including regulating the immune system [1, 2, 3, 4], hypotensive effect [5], inhibiting the platelet aggregation [6], stimulating the hepatic energy metabolism [7], improving the renal disorder [8], antioxidant activities [9, 10] and modulating the apoptosis homeostasis [11]. Furthermore, sterols isolated from Cordyceps sinensis were shown to be a great inhibitor to the proliferation of K562, Jukart, WM-1341, HL-60 and RPMI-8226 tumor cell lines [12]. In addition, Yang et al. (2002) [13] reported that H1-A extracted from Cordyceps sinensis can suppress the proliferation of human mesangial cell and may evoke the apoptosis by down-regulating the tyrosine phosphorylation of Bcl-2 and Bcl-XL. The fruit bodies of Cordyceps sinensis are rare and expensive. Therefore, the fermentation-grown mycelia with the pharmacological activities draw many attentions. However, different fermentation conditions may have the different impacts on the therapeutic activities of Cordyceps sinensis. The metabolites synthesized in Cordyceps sinensis may be altered by changing the nutritional fermentation conditions due to the differential protein expressions. Nowadays no any work has been focused on investigating the globe protein expression in Cordyceps sinensis influenced by the nutritional fermentation condition. Two-dimensional (2D) gel electrophoresis can visualize hundreds of proteins at a time, which coupled with mass spectrometry results in their identification. This technique has been applied to examine the global changes in the proteome map in Saccharomyces cerevisiae in response to the fermentation stress [14, 15] as well as environmental stimuli such as cadmium [16], lithium [17], H2O2 [18], or sorbic acid [19] and to the metabolic engineering in E. coli [20]. In this study, different liquid media containing various carbon sources were used for the cultivation of a strain of Cordyceps sinensis which was isolated in Yunnan province of People Republic of China and called G308-1 strain. The extracts of G308-1 strain grown in different liquid media were examined for their anticancer activities. Besides, the proteome maps of mycelia collected from the different media were compared to figure out the impact of various carbon sources on the protein expression.
Materials and Methods
Cultivation of the G308-1 Strain
The fungus strain used in this study was provided by Safe Protecta Technology Co., Ltd, Tainan, Taiwan and named as G308-1 strain. The voucher specimens were deposited at Culture Collection Laboratory of Department of Biotechnology, Southern Taiwan University of Technology, Tainan, Taiwan. The G308-1 strain was isolated from the fruit bodies of C. sinensis grown at Deh- Chin city of Yunnan province, People Republic of China. The G308-1 strain was cultivated in 500 ml shake flask containing 100 ml of liquid media [0.5 % (w/v) yeast extract, 0.05 % (w/v) MgSO4, 0.03 % (w/v) KH2PO4, 0.2 % (w/v) Na2HPO4 and 3 % (w/v) each of various carbon sources (glucose, maltose, sucrose or lactose)] with 250 rpm at pH 7.0 and 17°C for 7 days till the exponential phase reached.
Preparation of the Extract from the G308-1 Strain
The water extracts of G308-1 strain were prepared by incubating the mycelia with 60℃ water for 2 hours at the ratio of 1 to 10 (w/v). Then the suspension was centrifuged at 3,000 ×g and the supernatant was passed through a 0.2-μm pore size filter (Millex GP Carrigtwohill Co., Cork, Ireland). The filtrate was evaporated to one- tenth of the original volume. The ethanol extracts of G308-1 were prepared as in preparation of the water extract except that the mycelia were incubated with 95% of ethanol for 16 hour at 25℃ at the ratio of 1 to10 (w/v). 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay: Various concentrations of the water or ethanol extracts were added to a 96-well plate already loaded with HepG2, Du145, Hela or MCF-7 cells per well. After treatment for 72 hours, 20 μl of MTT solution (Merck, Damstadt, German) (5 mg/ml PBS) was added to each well and the plate was incubated at 37ºC for 4 hours. After medium removal, 200 μl of DMSO was added to each well and the plate was gently shaken for 5 minutes. The absorbance was determined at 540 nm. Triplicate wells were applied to each concentration for 72 hours as shown in Figures 4 and 5. The untreated or ethanol-treated cells were employed as the control. Isolation of Genomic DNA and Polymerase Chain Reaction The genomic DNA of G308-1 strain was isolated from the mycelia according to the Chelex method [21]. Briefly, an Lee ST, Huang MY and Wu TF. Investigation of Proteomic Responses and Anticancer Activities of an Isolated Strain of Cordyceps sinensis Under Different Nutritional Fermentation Conditions. J Microbiol Biotechnol, 2019, 4(1): 000137.
aliquot of about 300 µl Chelex 100 (Bio-Rad 36897, 50-100 mesh, Richmond, CA) was deposited in a sterile Eppendorff tube containing 500 µl of sterile water. Then small amount of the mycelia was transferred into the Eppendorff tube, crashed the mixture by Eppendorff stick and incubated at 95°C for 10 minutes. The crashed solution was centrifuged at 13,000 rpm for 2 minutes. The supernatant was collected and diluted with sterile water by 100 times. These samples could be subjected to PCR.
The rDNA (nuclear ribosomal DNA) regions of the genome including 18S, ITS (internal transcribed spacers), 5.8S and 25S are highly conserved among the fungi. Thus their sequences are usually subjected to phylogenetic analyses. Five pairs of primers [21] were used to perform PCR amplification and sequencing of the rDNA regions in G308-1 strain. As illustrated in Figure 1A, the primers ITS1 (5'-TCCGTAGGTGAACCTGCGG)/ITS4 (5'-TCCTCCGCTTATT GATATGC) were used for the amplification of ITS region while NS1 (5'-GTAGTCATATGCTTGTCTC)/NS2 (5'-GGCTGCTGGCACCAGACTTGC), NS3 (5'-GCAAGTCTGGTGCCAGCAGCC)/NS4 (5'-CTTCCGTCAATTGCTTTAAG) and NS5 (5'-AACTTAAAGGAATTGACGGAAG)/NS8 (5'-TCCGCAGGTTCACCTACGGA) were used for the amplification of 18S rDNA and NL1 (5'-GCATATCAATAAGCGGAGGAAAAG)/NL4 (5'-CCAGGCACAAAGTTCTGCC) were used for the amplification of 25S rDNA region.
The PCR reaction was performed in a mixture containing 25ng of DNA template, 10 µg of primer, 2 µl of 25mM dNTP, and 5 µl of 10x reaction buffer (100 mM Tris-HCl, pH9.0, 500mM KCl, 15mM MgCl₂, 1% Triton X-100) and 0.2 µl of Taq polymerase (5 U/µl). The thermal program for PCR amplifications consisted of a single denaturing step for 2 minutes at 95°C, followed by 35 cycles of denaturing at 95°C for 30 seconds, annealing at 47°C~52°C for 30 seconds and elongating at 72°C for 30 seconds. The reaction was completed with a single cycle of 72°C for 7 minutes. The PCR products were separated by electrophoresis and these rDNAs were sequenced by Applied Biosystems Ltd. Co. (Taipei, Taiwan). The resulting DNA sequence were analyzed using the BLAST software and MEGA 3 software based on the nucleotide sequences available in the GeneBank database.
Preparation of Protein Lysate
After fermentation, the mycelia were collected by centrifugation at 3,900 ×g from the liquid media and then dried in Speed Vac (ThermoSavant, Milford, MA) for 5 minutes. 0.2 g of pelleted mycelia were ground in liquid nitrogen and lysed in 1 m lysis buffer [7M urea, 2M thiourea, 4% (v/v) CHAPS, 100 mM dithiothreitol, 40 mM Tris pH 10, 20 µl Roche Complete mini protease inhibitor cocktail per 500 µl buffer] at room temperature for 1 hour. Samples were then centrifuged at 18,300 ×g for 20 minutes. The supernatant was collected and centrifuged again at 349,000×g for 3 hours at 15°C in Optima L-90K ultracentrifuge (Beckman Coulter, Fullerton, CA). After centrifugation, the supernatant was cleaned with 2D clean kit (Amersham-Pharmacia Biotech Inc., Piscataway, NJ) according to the manufacture’s suggestion and the protein pellet was dissolved in rehydration buffer [7M urea, 2M thiourea, 4% (v/v) CHAPS, 26mM dithiothreitol, 0.5% (v/v) IPG buffer and trace amount of bromophenol blue] and stored at -80 °C until use. The protein concentration was measured using a DC protein assay kit (BioRad, Hercules, CA).
Two-Dimensional Gel Electrophoresis (2-DE)
The immobiline dry strips (Pharmacia pl 4-7L, 11 cm) were rehydrated at 20°C for 16 hours with 200 µl rehydration buffer containing 60µg of protein. The proteins were then focused at 200V, 500V, 1000V, 5000V and 8000V with a total of 43,700 voltage-hours. After isoelectric focusing, the gel strips were incubated in dithiothreitol equilibration buffer [0.05M Tris pH 8.8, 6M urea, 30% (v/v) glycerol, 2% (w/v) SDS, and 1% (w/v) dithiothreitol] for 15 min and then in iodoacetamide equilibration buffer [0.05M Tris pH 8.8, 6M urea, 30% (v/v) glycerol, 2% (w/v) SDS, and 2.5% (w/v) iodoacetamide] for a further 15 min. The equilibrated gels were loaded onto the top of 12.5% acrylamide gel and sealed with 0.5% agarose. The proteins were separated until bromophenol blue reaches the bottom of the gel.
Silver Staining
Silver staining were performed according to the protocol described by Yan et al. In brief, the gels were fixed in fixation solution (ethanol/water/acetic acid, 4/5/1, v/v) after electrophoresis and soaked in sensitizing solutions [30 % (v/v) ethanol, 6.8 % (w/v) sodium acetate, 0.2 % (w/v) sodium thiosulphate and 0.125 % (v/v) glutardialdehyde] for 30 minutes. After sensitization, the gels were washed, incubated in silver nitrate solutions [0.25 % (w/v) silver nitrate and 0.0148 % (v/v) formaldehyde] for 20 min and then developed with the developing solution [2.5% (w/v) sodium carbonate and 0.0074 % (v/v) formaldehyde].
Image Analysis
The proteome profiles of the G308-1 strain were analyzed by PDQuest (BioRad) software to search for differentially expressed proteins. Six replica well-focused gels collected from three batches of the mycelia in each liquid medium were compared. Differentially expressed spots found by computer analysis were further examined by visualization. The intensity of the spot was computed and normalized as a percentage of the total intensity of all spots in a gel and analyzed with student’s t-test (Pharm PCS Version 4.2, Springer-Verlag, New York, USA). In all cases, statistical variance of the spot intensity ratio within 95% (Student’s t-test; p<0.05) was considered to be significantly different.
Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
Electrospray ionization (ESI) tandem mass spectrometry was performed as described previously [22, 23] using a ThermoFinnigan LCQ Deca ion trap mass spectrometer interfaced with an Agilent 1100 HPLC system. The spots of interest were excised from the gel and digested with modified trypsin (Amersham- Pharmacia Biotech Inc., Piscataway, NJ) as described previously [21]. The digested peptides were separated in an Agilent ZORBAX 300SB-C18 column using a mobile phase of solution A (0.1% formic acid in water) and solution B (0.085% formic acid in acetonitrile). The peptides eluted from the HPLC were introduced on-line to the ESI source and the spectra were acquired as successive sets of three scan modes (MS, Zoom and MS/MS scans). The acquired collision-induced dissociation spectra were interpreted using TurboSequest software (ThermoFinnigan, San Jose, CA) which matches predicted tandem mass spectra against non-redundant protein database.
Results Determination of the phylogenetic relationship of the G308-1 strain to Cordyceps sinensis
Since the G308-1 strain was isolated from the fruit bodies (teleomorph; sexual stage) of C. sinensis, the G308- 1 strain cultivated in the liquid media might be the anamorph (asexual stage). In order to demonstrate that the G308-1 strain was the anamorph, the mycelia were authenticated using the conventional fungal classification methods by Dr. Roland Kirschner in Botany Institute University of Frankfurt, German. The classification results suggested that the G308 strain belonged to Paecilomyces farinosus (data not shown). To further confirm this identification, various regions of rDNA of the G308-1 strain including the internal transcribed spacers (ITS) were amplified with the polymerase chain reaction. As shown in Figure 1A, five pairs of PCR primers (NL1&NL4, ITS1&ITS4, NS1&NS2, NS3&NS4 and NS5&NS8) were utilized to amplify the rDNA regions as described by Chen et al. (2001)[22]. The PCR fragments with the expected sizes could be amplified from rDNA using each primer pair (Figure 1B, lanes 2-6). The nucleotide sequence of each PCR fragment was compared against GeneBank using the BLAST software. The comparative results showed that each nucleotide sequence was nearly 100 % identical to that of Paecilomyces farinosus. In order to further analyze the link of the G308-1 strain to P. farinosus, the distance values of ITS sequences between P. farinosus and its allies were estimated using MEGA 3 software and presented in Table 1. The zero distance value between the G308-1 strain and P. farinosus indicated that the G308-1 strain should belong to P. farinosus. In addition, the phylogenetic tree was reconstructed from the distance values using the neighbor-joining (JN) method to confirm the relationship between the G308-1 strain and P. farinosus. As demonstrated in Figure 1C, the G308-1 strain and P. farinosus were combined into a single clade.
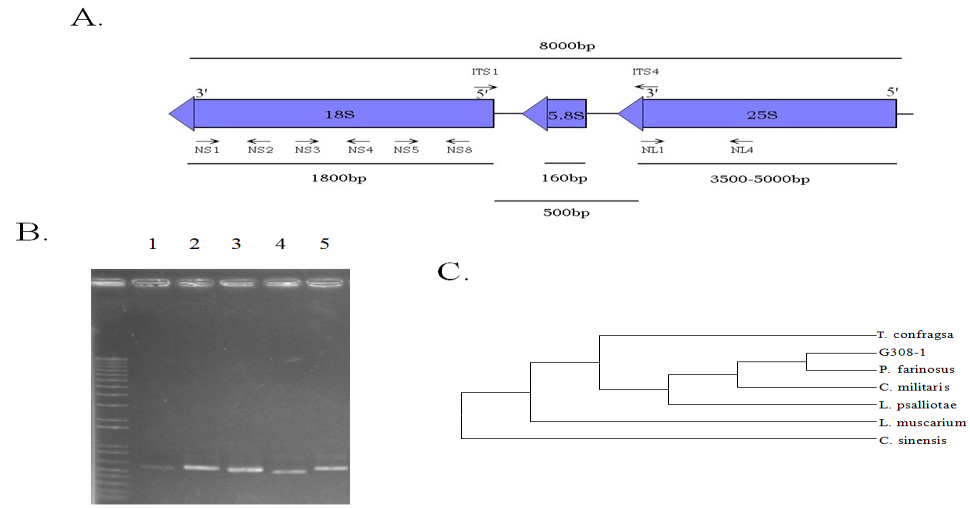
Figure 1: The molecular identification of Paecilomyces farinosus using rDNA regions. (A) The locations of the primers used in the polymerase chain reactions. (B) The PCR products amplified from the genomes of P. farinosus using the above primers. Lane 1, PCR with NL1 and NL4; lane 2, ITS1 and ITS4; lane 3, NS1 and NS2; lane 4, NS3 and NS4; lane 5, NS5 and NS8. (C) Molecular phlyogenetic tree inferred from the ITS of G308-1 strain and its allies.
Lee ST, Huang MY and Wu TF. Investigation of Proteomic Responses and Anticancer Activities of an Isolated Strain of Cordyceps sinensis Under Different Nutritional Fermentation Conditions. J Microbiol Biotechnol, 2019, 4(1): 000137.
| G308-1 | P. farinosus | T. confragosa | V. psalliotae | L. muscarium | C. militaris | C. sinensis | |||||||||||||||
| G308-1 | - | 0 | 0.04 | 0.04 | 0.04 | 0.07 | 0.54 | ||||||||||||||
| P. farinosus | - | 0.04 | 0.04 | 0.04 | 0.07 | 0.54 | |||||||||||||||
| T. confragosa | - | 0.03 | 0 | 0.06 | 0.49 | ||||||||||||||||
| V. psalliotae | - | 0.03 | 0.07 | 0.53 | |||||||||||||||||
| L. muscarium | - | 0.06 | 0.49 | ||||||||||||||||||
| C. militaris | - | 0.55 | |||||||||||||||||||
| C. sinensis | - |
Table 1: Distance values between the G308-1 and its allies.
The Proteome Profile Changes of the G308-1 Strain Grown Under the Different Nutritional Media
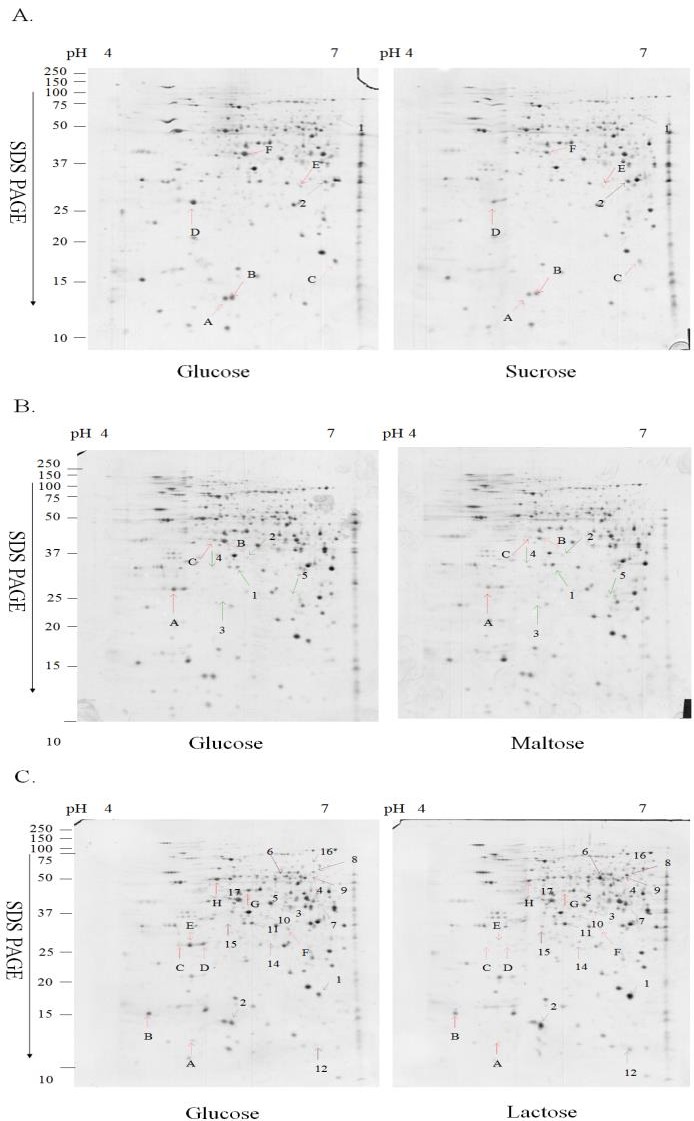
Upon exposure to a specific carbon source, cells adopt a corresponding metabolic pathway by adjusting the proteome profiles to assimilate the carbon source for the energy. In order to evaluate the proteome profile changes in response to various carbon sources, two-dimensional gel electrophoreses (2-DE) were carried out to analyze the proteome profiles of the G308-1 strain grown in the liquid media containing glucose, sucrose, maltose or lactose as the sole carbon source. The proteome profiles of the G308-1 strain grown in the glucose media were compared respectively with those in the other three media. The representative 2-DE maps of the G308-1 strain cultivated under the different nutritional conditions were illustrated in Figure 2. On average each gel resolved up to 850 protein spots. To prevent the variation of silver staining, six replica gels were collected from three batches. The differentially expressed protein spots were scored which is higher than 1.5-fold in magnitude as observed in all six replica gels. Eight, eight and twenty- five differentially expressed proteins were observed respectively in the mycelia derived from the sucrose, maltose and lactose media. Among these differentially expressed protein spots, two, five and seventeen protein spots were repressed respectively in the mycelia grown in the sucrose, maltose and lactose media while six, three and eight protein spots were induced respectively. The total normalized volume (ppm), _p_I, molecular weight and the statistical result of each protein spot were presented in Tables 2-4.
Lee ST, Huang MY and Wu TF. Investigation of Proteomic Responses and Anticancer Activities of an Isolated Strain of Cordyceps sinensis Under Different Nutritional Fermentation Conditions. J Microbiol Biotechnol, 2019, 4(1): 000137.
Figure 2: The 2-D maps of the mycelia grown in the liquid media containing glucose, sucrose, maltose and lactose. (A) The comparisons of the proteomes of the mycelia cultivated in glucose media to those in sucrose media. (B) The proteomes in glucose media to those in maltose media. (C) The proteomes in glucose media to those in Spot No pI/Mra glucoseb (ppm) Foldc P value 1 5.98/60.7 16.18±4.03 30.20±6.02 +1.87 P<0.05 2 5.91/32.1 77.5±27.98 131.16±19.94 +1.69 P<0.05 A 5.14/13.5 102.93±6.87 56.32±6.06 -1.83 P<0.01 B 5.21/13.7 127.92±12.23 81.97±6.61 -1.56 P<0.05 C 5.95/17.5 159.33±38.38 39.15±8.74 -4.07 P<0.05 D 4.78/26.8 476.72±159.22 176.6±61.37 -2.7 P<0.05 E 5.74/30.7 23.33±8.17 10.27±3.92 -2.27 P<0.05 F 5.33/44.7 239.03±61.76 165.83±66.19 -1.44 P<0.01 (ppm) lactose media. The differentially expressed protein spots were indicated by the arrows. The upregulated proteins in sucrose, maltose or lactose media were shown by the Arabic numbers while the down-regulated proteins were indicated by the letters. The figures were the representative gels of six replica gels from three batches.
surcose
- apI: isoelectric point; Mr: molecular weight
- bLglu: protein spots in the 2D gels of the mycelia grown in glucose media
- c+: proteins up-regulated in the mycelia grown in sucrose media; -: protein down regulated in the mycelia grown in sucrose media
Table 2: The differentially expressed protein spots between the mycelia grown in glucose and sucrose media.
| pI/Mr | glucose (ppm) | maltose (ppm) | Fold | P value | |
|---|---|---|---|---|---|
| 1 | 5.45/34.0 | 46.92±10.45 | 96.47±9.7 | 2.06 | P<0.05 |
| 2 | 5.53/34.2 | 3.03±0.48 | 9.8±1.55 | 3.23 | P<0.01 |
| 3 | 5.31/24.9 | 0.4±0.2 | 12.48±2 | 31.2 | P<0.01 |
| 4 | 5.22/31.5 | 1.57±0.41 | 32.37±5.17 | 20.62 | P<0.01 |
| 5 | 5.85/24.4 | 1.93±0.7 | 7.97±2.45 | 4.13 | P<0.05 |
| A | 4.78/26.8 | 256.5±80 | 56.77±9.74 | -4.52 | P<0.05 |
| B | 5.33/40.6 | 160.93±20.1 | 84.45±12.33 | -1.9 | P<0.01 |
| C | 5.23/40.8 | 34.25±6.65 | 11.35±3.92 | -3.02 | P<0.01 |
Table 3: The differentially expressed protein spots between the mycelia grown in glucose and maltose media.
| Spot No | pI/Mr | glucose (ppm) | lactose (ppm) | Fold | P value | |||||||||||
| 1 | 5.95/17.5 | 81.86±8.34 | 502.2±83.9 | 6.13 | P<0.01 | |||||||||||
| 2 | 5.21/13.7 | 82.3±10.6 | 150.7±30.53 | 1.83 | P<0.05 | |||||||||||
| 3 | 5.66/35.7 | 0.52±0.23 | 27.6±5.61 | 53.08 | P<0.01 | |||||||||||
| 4 | 5.89/40.9 | 207.08±35.02 | 259.43±33.63 | 1.25 | P<0.05 | |||||||||||
| 5 | 5.64/54.2 | 18.18±4 | 109.8±36.15 | 6.04 | P<0.05 | |||||||||||
| 6 | 5.74/54.2 | 57.23±9.97 | 267.88±73.92 | 4.68 | P<0.05 | |||||||||||
| 7 | 5.93/39.3 | 45.82±9.81 | 136.55±17.96 | 2.98 | P<0.01 | |||||||||||
| 8 | 5.94/60.0 | 21.42±6.4 | 69.15±18.56 | 3.23 | P<0.05 | |||||||||||
| 9 | 5.91/55.8 | 3.65±0.88 | 23.83±5.86 | 6.53 | P<0.05 | |||||||||||
| 10 | 5.53/34.2 | 3.1±0.68 | 20.9±2.89 | 6.74 | P<0.01 |
Lee ST, Huang MY and Wu TF. Investigation of Proteomic Responses and Anticancer Activities of an Isolated Strain of Cordyceps sinensis Under Different Nutritional Fermentation Conditions. J Microbiol Biotechnol, 2019, 4(1): 000137.
| 11 | 5.45/34.0 | 47.03±8.53 | 120.92±12.07 | 2.57 | P<0.01 |
|---|---|---|---|---|---|
| 12 | 5.96/11.6 | 0.96±0.47 | 11±3.74 | 11.46 | P<0.05 |
| 14 | 5.58/26.8 | 0.65±0.18 | 10.78±2.31 | 16.58 | P<0.01 |
| 15 | 5.22/31.7 | 3.73±0.46 | 14.9±1.65 | 3.99 | P<0.01 |
| 16 | 5.91/66.0 | 0.57±0.009 | 12.9±4.32 | 22.63 | P<0.05 |
| 17 | 5.36/34.0 | 4.92±1.79 | 105.43±27.81 | 21.43 | P<0.05 |
| A | 4.82/12.6 | 4.22±0.76 | 0.32±0.2 | -13.5 | P<0.01 |
| B | 4.37/14.8 | 72.87±24.07 | 43.32±17.8 | -1.68 | P<0.05 |
| C | 4.69/28.8 | 12.52±2.52 | 0.75±0.27 | -16.7 | P<0.01 |
| D | 4.96/27.0 | 157.38±50.34 | 3.03±0.75 | -51.94 | P<0.05 |
| E | 4.78/26.8 | 140.1±23.61 | 2.9±0.75 | -48.31 | P<0.01 |
| F | 5.74/30.7 | 18.6±2.34 | 2.92±0.79 | -6.37 | P<0.05 |
| G | 5.42/44.5 | 107.32±17.06 | 15.2±2.07 | -7.06 | P<0.01 |
| H | 5.12/50.8 | 161.02±15.16 | 96.92±21.64 | -1.66 | P<0.01 |
mass spectrometry (LC-MS/MS) and TurboSequest software. No significant hits were acquired for the protein spots 1 and 17. Nevertheless, the de novo sequencing was performed to interpret the peptide sequences of two proteins (Figure 3). Two peptides of the protein spot 11 were identified to be related to the aldehyde dehydrogenase of Alternaria alternata (alternaria rot fungus).

Lee ST, Huang MY and Wu TF. Investigation of Proteomic Responses and Anticancer Activities of an Isolated Strain of Cordyceps sinensis Under Different Nutritional Fermentation Conditions. J Microbiol Biotechnol, 2019, 4(1): 000137.
The Anticancer Activities of the G308-1 Strain Vrown in various Liquid Media
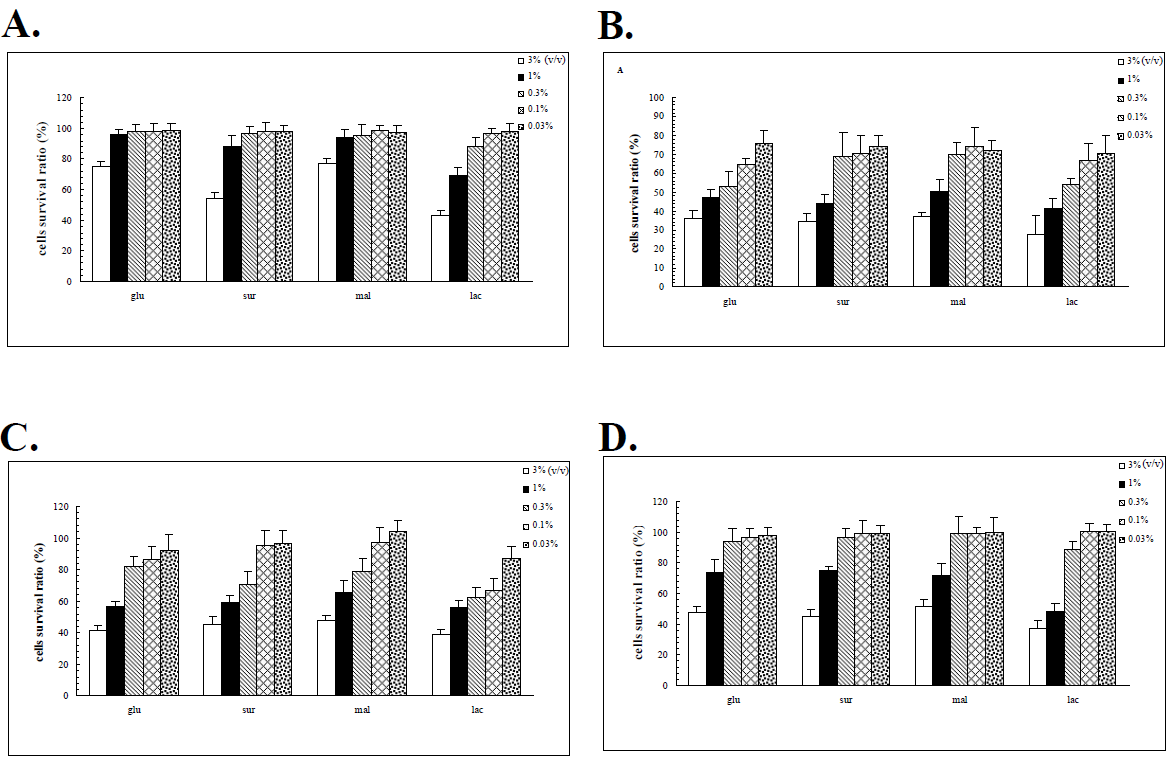
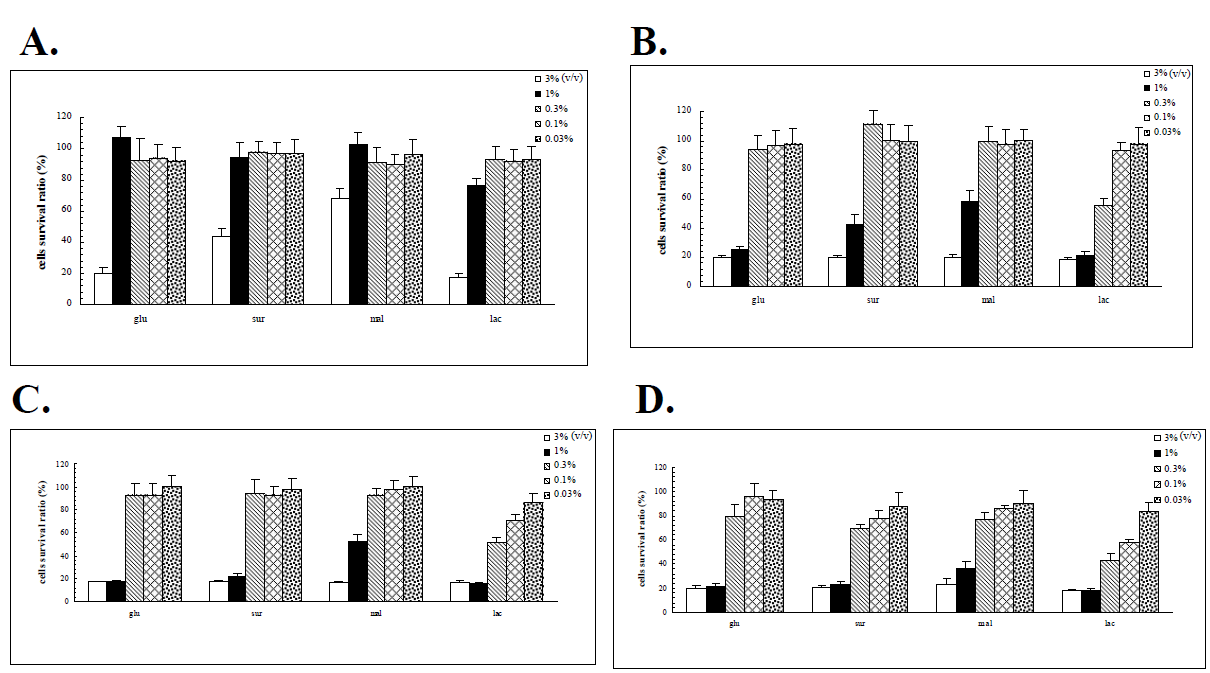
The proteomic responses of the G308-1 strain in various liquid media suggested that the G308-1 strain might adopt a corresponding proteome in response to a specific carbon source. This response, in turn, might elicit a specific array of bio-products including primary and secondary metabolites which are synthesized in the mycelia and possibly relevant to the anticancer activities of the fungi. In order to examine the anticancer activities of the G308-1 strain cultured under various nutritional conditions the water or ethanol extracts of the G308-1 strain cultivated in the glucose, sucrose, maltose or lactose media were explored for their abilities to suppress the HepG2, Du145, Hela and MCF-7 cells. The preliminary results demonstrated that the water extracts had the best inhibitory results to the cancer cells when the extracts were made by incubating the mycelia with the 60°C water at 1:10 ratio (v/v) (data not shown). As illustrated in Figure 4, the suppressive effects augmented essentially with the increasing concentrations of the water extracts [0.03 % to 3 % (v/v)] when each cancer cell line was treated with each water extract. Hela and Du 145 cells were more sensitive to the water extracts than HepG2 and MCF-7 cells at all the applied concentrations in terms of each water extract. At low concentrations (0.03% to 0.3%) the HepG2 and MCF-7 cells were barely harmed by each water extract. However, high concentrations (1% and 3%) of each water extract showed the repressive impacts nearly on all the cell lines. At high concentrations the water extracts made from the mycelia in the lactose media had better inhibitory results to the HepG2 cells than those from the other three media. Figure 5 demonstrated all the cell lines were essentially insensitive to low concentrations of the ethanol extracts made from the mycelia in glucose, sucrose or maltose. However, at low concentrations the ethanol extracts produced from the mycelia in the lactose media exerted the better inhibitory activities to the Du145, Hela and MCF-7 cells. In contrast to low concentrations, high concentrations of each ethanol extract showed much stronger suppressive influences on all the cell lines. Except for the HepG2 cell at high concentrations there were no significant differences between the efficacies of all four ethanol extracts for each cell line (Figure 5). The ethanol extracts produced from the mycelia in the lactose media showed much better repressive activities to the HepG2 cell than those from the other three media (Figures 4 and 5). In addition, the HepG2 cells were only inhibited by the highest concentration of each ethanol extract.
Lee ST, Huang MY and Wu TF. Investigation of Proteomic Responses and Anticancer Activities of an Isolated Strain of Cordyceps sinensis Under Different Nutritional Fermentation Conditions. J Microbiol Biotechnol, 2019, 4(1): 000137.
Discussion The fungus strain utilized in this study was isolated in Yunnan province of People Republic of China and identified as Paecilomyces farinosus. Although the link of Paecilomyces farinosus to the anamorph of C. sinensis is still under controversy, P. farinosus possesses the pharmacological activities [23, 24]. To gain insight into the biological effects of various carbon sources and to evaluate the proteome profile changes evoked by cell’s adaptation to the different nutritional conditions, two-dimensional gel electrophoreses were utilized to monitor the proteome changes of a Paecilomyces farinosus strain (G308-1 strain) cultivated in liquid media containing glucose, sucrose, maltose and lactose. Because glucose is the best carbon source for the fungi, the proteomes of the mycelia cultivated in the liquid media containing glucose were compared with those of the mycelia in the sucrose, maltose or lactose media. The comparative results demonstrated that some proteins were induced or repressed under the different nutritional conditions (Tables 2-4). The most proteome profile change was elicited in the mycelia in the lactose media with 17 down- regulated and 8 up-regulated proteins (Table 4).
However, much fewer differentially expressed proteins (eight and eight) were found in the mycelia derived from the sucrose and maltose media (Tables 2 and 3). Consistent with this result, Trabalzini, et al. (2003) [15] reported that 54 proteins are up-regulated and 138 proteins are down-regulated during the physiological fermentation stress under which glucose concentration gradually decreases in the YPD media. The dramatic proteome profile change induced by the lactose media suggested that the fungi might adopt a more complicated metabolic pathway to metabolize the lactose. In contrast, it might be easier for the fungi to metabolize the sucrose and maltose. In this study, the G308-1 strain at exponential stage was utilized for the proteomic analyses and therefore the proteome profile change was not due to the physiological fermentation stress. The proteomic comparisons of the G308-1 strain in various nutritional media indicated that a specific carbon source might induce a corresponding proteome in the mycelia and probably, in turn, a specific array of bio- products which likely contribute to the anticancer activities of the fungi. The anticancer results demonstrated that the water extracts showed the selectivity to Hela and Du 145 cells and for HepG2 cells at high concentrations the water extracts made from the Lee ST, Huang MY and Wu TF. Investigation of Proteomic Responses and Anticancer Activities of an Isolated Strain of Cordyceps sinensis Under Different Nutritional Fermentation Conditions. J Microbiol Biotechnol, 2019, 4(1): 000137.
mycelia in the lactose media had better suppressive effects (Figure 4). As to the ethanol extract, all the cell lines were almost insensitive to low concentrations of the extracts but high concentrations of the extracts showed much stronger inhibitory abilities. Conclusively the ethanol extracts prepared from the mycelia in the lactose media showed much better inhibitory activities to each cell line than those from the other three media (Figure 5). The above results implicated that the water or ethanol extracts of the G308-1 strain showed the selectivity to the cancer cell lines. In addition, the anticancer results might vary with the proteome profile changes which were evoked by a specific carbon source and probably resulted in the production of a different array of bio-products in the mycelia under the different nutritional conditions. This study was the first paper that described the proteome changes of a fungus induced by the different nutritional conditions. If the differentially expressed proteins could be identified in the time when the genome annotations of fungi are available in the protein database, the metabolic pathways adopted in response to a specific carbon source can be interpreted. Therefore, relying upon the interpreted metabolic pathway, the metabolites correlated with the pharmacological activities may be predicted. In addition, the metabolic pathways may be changed genetically using various molecular biological tools to improve the production of the metabolites.
Conflict of Interest Disclosure The authors declare that there is no conflict of interest regarding the publication of this paper Funding Acknowledgement This work was supported by from the Ministry of Science and Technology of Taiwan (grant number MOST 105-2320-B-218-001-MY3).
References 1. Chen GZ, Chen GL, Sun T, Hsieh GC, Henshall JM
(1991) Effects of Cordyceps sinensis on murine T lymphocyte subsets. Chin Med J (Engl) 104(1): 4-8.
2. Zhang Z, Xia SS (1990) Cordyceps Sinensis-I as an immunosuppressant in heterotopic heart allograft model in rats. J Tongji Med Univ 10(2): 100-103.
3. Zhu JS, Halpern GM, Jones K (1998) The scientific rediscovery of a precious ancient Chinese herbal regimen: Cordyceps sinensis: part II. J Altern Complement Med 4(4): 429-457.
4. Kuo YC, Tsai WJ, Shiao MS, Chen CF, Lin CY (1996)
Cordyceps sinensis as an immunomodulatory agent. Am J Chin Med 24(2): 111-125.
5. Feng MG, Zhou QG, Feng GH (1987) Vasodilating
effect of cultured Cordyceps sinensis (Berk) Sacc. mycelia in anesthetized dogs. Zhong Yao Tong Bao 12(12): 41-45.
6. Shiao MS, Wang ZN, Lin LJ, Lien JY, Wang JJ (1994)
Profiles of nucleosides and nitrogen bases in Chinese medicinal fungus Cordyceps sinensis and related species. Botanical Bulletin of Academia Sinica 35(4): 261-267.
7. Manabe N, Sugimoto M, Azuma Y, Taketomo N,
Yamashita A, et al. (1996) Effects of the mycelial extract of cultured Cordyceps sinensis on in vivo hepatic energy metabolism in the mouse. Jpn J Pharmacol 70(1): 85-88.
8. Zhao X, Li L (1993) Cordyceps sinensis in protection of the kidney from cyclosporine A nephrotoxicity. Chin Med J 73(7): 410-412.
9. Yamaguchi Y, Kagota S, Nakamura K, Shinozuka K,
Kunitomo M (2000) Antioxidant activity of the extracts from fruiting bodies of cultured Cordyceps sinensis. Phytother Res 14(8): 647-649.
10. Li SP, Li P, Dong TT,Tsim KW (2001) Anti-oxidation
activity of different types of natural Cordyceps sinensis and cultured Cordyceps mycelia. Phytomedicine 8(3): 207-212.
11. Buenz EJ, Bauer BA, Osmundson TW, Motley TJ
(2005) The traditional Chinese medicine Cordyceps sinensis and its effects on apoptotic homeostasis. J Ethnopharmacol 96(1-2): 19-29.
12. Bok JW, Lermer L, Chilton J, Klingeman HG, Towers
GH (1999) Antitumor sterols from the mycelia of Cordyceps sinensis. Phytochemistry 51(7): 891-898.
13. Yang LY, Huang WJ, Hsieh HG, Lin CY (2003) H1-A
extracted from Cordyceps sinensis suppresses the proliferation of human mesangial cells and promotes apoptosis, probably by inhibiting the tyrosine phosphorylation of Bcl-2 and Bcl-XL. J Lab Clin Med 141(1): 74-83.
14. Kolkman A, Olsthoorn MM, Heeremans CE, Heck AJ,
Slijper M (2005) Comparative proteome analysis of Lee ST, Huang MY and Wu TF. Investigation of Proteomic Responses and Anticancer Activities of an Isolated Strain of Cordyceps sinensis Under Different Nutritional Fermentation Conditions. J Microbiol Biotechnol, 2019, 4(1): 000137.
Saccharomyces cerevisiae grown in chemostat cultures limited for glucose or ethanol. Mol Cell Proteomics. 4(1): 1-11.
15. Trabalzini L, Paffetti A, Scaloni A, Talamo F, Ferro E et al. (2003) Proteomic response to physiological fermentation stresses in a wild-type wine strain of Saccharomyces cerevisiae. Biochem J 370(1): 35-46.
16. Vido K, Spector D, Lagniel G, Lopez S, Toledano MB, et al. (2001) A proteome analysis of the cadmium response in Saccharomyces cerevisiae. J Biol Chem. 276(11): 8469-8474.
17. Bro C, Regenberg B, Lagniel G, Labarre J, Montero-
Lomeli M, Nielsen, J (2003) Transcriptional, proteomic, and metabolic responses to lithium in galactose-grown yeast cells. J Biol Chem 278(34): 32141-32149.
18. Godon C, Lagniel G, Lee J, Buhler JM, Kieffer S, et al.
(1998) The H2O2 stimulon in Saccharomyces cerevisiae. J Biol Chem 273(35): 22480-22489.
19. de Nobel H, Lawrie L, Brul S, Klis F, Davis M, et al.
(2001) Parallel and comparative analysis of the proteome and transcriptome of sorbic acid-stressed Saccharomyces cerevisiae. Yeast 18(15): 1413-1428.
20. Han MJ, Lee SY (2003) Proteome profiling and its use in metabolic and cellular engineering. Proteomics 3(12): 2317-2324.
21. Chen YQ, Wang N, Qu LH, Li TH, Zhang WM (2001)
Determination of the anamorph of Cordyceps sinensis inferred from the analysis of the ribosomal DNA internal transcribed spacers and 5.8S rDNA. Biochem Syst Ecol 29(6): 597-607.
22. Yan JX, Wait R, Berkelman T, Harry RA, Westbrook JA,
et al. (2000) A modified silver staining protocol for visualization of proteins compatible with matrix- assisted laser desorption/ionization and electrospray ionization-mass spectrometry. Electrophoresis 21(17): 3666-3672.
23. Lang G, Blunt JW, Cummings NJ, Cole AL, Munro MH
(2005) Paecilosetin, a new bioactive fungal metabolite from a New Zealand isolate of Paecilomyces farinosus. J Nat Prod 68(5): 810-811.
24. Cheng Y, Schneider B, Riese U, Schubert B, Li Z, et al.
(2004) Farinosones A-C, neurotrophic alkaloidal metabolites from the entomogenous deuteromycete Paecilomyces farinosus. J Nat Prod 67(11): 1854- 1858.
Lee ST, Huang MY and Wu TF. Investigation of Proteomic Responses and Anticancer Activities of an Isolated Strain of Cordyceps sinensis Under Different Nutritional Fermentation Conditions. J Microbiol Biotechnol, 2019, 4(1): 000137.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania