Bioprospecting of Marine Bacteria for their Diversity and Biotechnological Application
Microorganisms from marine environments are gaining attention for their idiosyncratic ability to survive in these extreme conditions and been evaluated for their application in bioprospecting. From isolation of enzymes and bioactive compounds possessing peculiar abilities, identification of unique metabolic pathways, to application in health care and bioremediation, extensive study on these organisms is been done with tremendous success. Technological advancement in modern science with regards to Omics technologies integrated with genetic engineering technologies are paving way for isolation of novel organisms and their macromolecules. These bioengineered organisms and enzymes have found their application in industries with high commercial value.
Introduction
The oceans, seas, estuaries, and other aquatic systems with high concentrations of dissolved salts collectively comprise the marine environment, which covers ~70% of the Earth’s total surface area and hold more than 97% of the total water on Earth [1]. These environments are characterised by a large gradient that is evident in various environmental factors such as light, pressure, temperature, light intensity, etc [2]. The marine environment, with its habitat complexity and consequent increase in niches, forms numerous ecosystems rich in the diversity of life forms found, with microorganisms accounting for most of this diversity [3]. An important element that distinguishes the diversity and microbial composition of these habitats from terrestrial habitat is salinity. Compared to soil in terrestrial environments, sediments in marine environments have greater phylogenetic diversity [1]. Marine bacteria have acquired various physiological and metabolic modifications with their evolutionary mechanism to adapt to the extremes of pH, temperature, limited substrate concentration and distribution, and high pressure and salt concentration of the marine environment [4]. These organisms play a key role in the flux of organic matter and nutrients, and contribute to the degradation of many organic materials that help to sustain the marine environment [5]. Considering the extremophilic nature of these bacteria, interest in the study of marine bacteria has increased over the years to explore the potentials of these marine organisms for various ecological and commercial purposes.
The marine bacteria produce various complex metabolites to survive in the extreme conditions of the sea such as extremozymes, biopolymers, secondary metabolites and bioactive compounds like flavonoids, antioxidants, antibiotics, etc. These biomolecules with unique structural and functional capabilities hold potentials for novel application and in industries. Further identification of unique biochemical pathways from these may have potentials in the degradation of various organic and chemical compounds [2, 6]. Efforts to bio prospect these marine bacteria for their potentials in the production of various biomolecules have been carried out [7]. Therefore, these bacteria are now acknowledged as valuable resources for the production of various novel biomolecules, with numerous discoveries made in recent years [8]. However, due to the slow pace of technological progress, a large part of the marine ecosystem remains unexplored. The review paper examines the bacterial diversity of marine environment and its role in biotransformation processes that can be exploited for commercial applications such as pharmaceuticals, bioremediation, food supplements, enzyme production etc. The biotechnological advances and challenges in the study of these marine environments will also be discussed to promote a more comprehensive exploration of these environments along with their role in global climate change.
Marine Environment and Bacterial Diversity
The marine environment that stores a large volume of water at depths greater than 11,000 metres is referred to as a “high-pressure refrigerator” because the pressure of deep water increases at a rate of 1 atm/10 metres, with a maximum of 1000 atm in the deepest regions of these water bodies and a temperature below 3°C in regions 100 metres and deeper [9]. In addition to the extreme temperature and pressure conditions, a decreasing intensity of sunlight and a limitation of food supply, a limited supply or absence of oxygen are observed in the deeper seafloor environment [10]. Thus, until the end of the 19th century, it was believed that no life existed in these deep waters and was supported by Forbes’ azoic hypothesis based on his observation where he saw a decrease in the number of species with increasing water depth [11]. However, the discovery of living crinoids (sea lilies) from the depths of Norwegian fjords (∼3000 metres) by biologist Michael and Georg Ossian Sars, followed by research expeditions such as the HMS Challenger expedition, shed light on the diverse life forms in the ocean depths [12].
With the marine environment covering nearly 70% of the Earth’s surface, much of the Earth’s habitat is found in these aquatic systems (approximately 99%), with all of the major phyla of the animal kingdom that evolved from this habitat. Furthermore, much of this marine habitat remains inaccessible to humans. Therefore, the interest in exploring these deep-water systems to identify new organisms has become a fascination, with the discovery of new species regularly reported Kaiser MJ, et al. [13]. Because environmental parameters are constantly changing at different levels of the marine system, a wide variety of habitats can be found in the wilds of the oceans and seas. Based on the availability of sunlight, the ocean zones are divided into photic, mesopelagic, bathypelagic, abyssopelagic, and hadopelagic zones [14]. Other distinct environments in the ocean depths include the abyssal plain, hyperthermal vents, seamounts, mid-ocean ridges, continental margins, oceanic trenches, etc., with deposits of biogenic particles that may originate from various sources such as volcanic, terrigenous, and authigenic on the seafloor [10]. However, all the various habitat cannot be covered in this review so some of the major marine environments have been discussed below.
Environmental extremes and low nutrient availability limit biological productivity, thus large macro-organisms are generally not found in these deep-sea environments, with communities of low biomass species being the predominant life form here [10]. Microorganisms are the main components of the marine habitat with a predicted value of 3.6 ×1029 microbial cells present in the ocean system, of which >105 are present per ml of surface seawater. From bacteria and archaea to other microscopic organisms such as protists and fungi, the microbial diversity of the marine environment includes those that play a critical role in nutrient cycling and maintaining the stability of this ecosystem, contributing 98% of primary production [9]. Considering the ubiquitous nature of microorganisms and their importance in the marine ecosystem, it is expected that evolutionary variation and diversity can be observed in the extremes of the deep- sea habitat.
Abyssal Plain
These are submarine plains that form on the seafloor and account for more than 50% of the Earth’s surface [15]. They are the predominant habitat of the deep-water system and have high diversity with low overall biomass [16]. General environmental conditions of the abyssal plain include temperatures between -0.5 and 3°C, high dissolved oxygen levels, and slow or stagnant water flow. Seafloor sediment deposits consist of sands, clays, and biogenic compounds with a lack of primary production, limiting food availability. The main source of food and energy in these regions is the deposition of detritus, rich in organic matter, formed in the euphotic zones. Manganese nodules associated with hard substrate are also present and support the development of fauna different from that found in sediments [15]. The abyssal plains provide the ecosystem with important services such as biogeochemical cycling and the trapping of a large concentration of CO2 in the form of fossil fuels. Considering their large extent and inaccessibility, knowledge of the abyssal habitat and ecosystem has been limited. However, extensive studies have been conducted in recent decades showing the diversity of life forms in the abyssal plains. For example, nearly 2000 bacterial species have been discovered at a single study site alone, along with 500 invertebrates and 250 protozoan species [17].
Since the main food source of these habitats is detritus, microorganisms play an important role in the transformation of these nutrients and play an important role in the food chain and their diversity is directly correlated with the rate of flux of organic material from the euphotic zones and plays a key role in the various services provided by the abyssal plain ecosystem. Widespread marine microbes include the bacterial order Woeseiales Hoffmann KC, et al, [18] and the archaeal phylum Thaumarchaeota [19], which are under active investigation. However, the seamounts and hyperthermal vents that traverse these large submarine plains are rich in nutrients and form a completely different habitat [20].
Hydrothermal Vents
Hydrothermal vents are a marine ecosystem formed where seawater enters the Earth’s crust through fractures created by the movement of tectonic plates. The geothermal energy of the crust heats this cold water, which then flows out and forms a vent. Chemical leaching of rock on the seafloor alters the hot water so that it has high sulfur and iron content [21]. The mixing of hydrothermal fluids with cold seawater creates a thermal gradient that is accompanied by the geochemistry of the hydrothermal fluid, which also creates a gradient in pH and chemical species like copper, zinc, iron etc. that shape the microbial composition and further subdivide the hyperthermal vent into different habitats, e.g., hydrothermal vent, hydrothermal plumes, etc [22]. For life forms to exist in this region, they should be able to adapt to the gradient of all chemical, physical and biological gradients of this habitat. Microorganisms isolated from the hydrothermal vent show the characteristic property of stability at high temperatures and commitment to anoxic conditions. Chemoautotrophic microorganisms that use sulfur from inorganic compounds as an electron acceptor dominate this habitat, which also supplies the energy needs of other species and thus forms the backbone of the food chain [21]. Other chemosynthetic processes that serve as energy sources include the oxidation of methane, hydrogen, and iron [22]. Microorganisms belonging to different groups of Archaea and Bacteria have been isolated from hydrothermal vents, which include hyperthermophiles: Pyrococcus horikoshii [23], Thermococcus onnurineus [24], hyperthermophilic methanogens: Methanopyrus kandleri strain 116 [25], thermophiles: Thermosulfurimonas marina SU872T [26], Anoxybacillus caldiproteolyticus [27], Methanogens: Methanocaldococcus villosus and Methanothermococcus okinawensis [28], Thermopycophiles: sulfur reducer Marinitoga piezophile strain KA3T [29], Chemolithoautotrophs: Thioprofundum lithotrophicum strain 106 and Piezobacter thermophilus strain 108 [30], Sulfur-oxidizing bacteria: Sulfurimonas sp. NW10 [31], Hydrogenovibrio thermophilus strain S5 [32], etc.
Seamounts
Seamounts are underwater mountains formed by volcanic activity in the seafloor, similar to volcanic mountains but not reaching the sea surface [33]. For a marine system deeper than 200 metres, a minimum elevation of 100 metres above the ocean floor is the criterion for an elevation to be considered a seamount, and includes seamounts associated with the mid-ocean ridge, continental slope, or back-arc basin, as well as intraplate seamounts. In addition, they are subdivided based on elevation into hills and seamounts with elevations of 500 and 1000 metres from the seafloor, respectively [34]. Some geologic topologies in the range of 50 to 100 meters may also be considered seamounts [35]. Oceanic islands are not counted as seamounts, although they are thought to have originated from a seamount because their ocean system is completely different from seamounts due to runoff from the nearby coast, even though their community composition may be the same [34]. Seamounts with 1000 metres relief cover nearly 16% of the seafloor, while those with 1000 metres relief cover l4.7%, and their habitat is highly fragmented, making them a hotspot for biodiversity [34]. From macroscopic to microscopic and benthic to pelagic, various organisms are found in seamounts, which include microorganisms, and seamounts are studied to influence the diversity and distribution of microbial communities [36]. The presence of these topological structures affects the hydrography of water bodies, leading to increases in current velocity, tidal flow, and the generation of internal waves, which increases vertical mixing and the resulting output of nutrients from the ocean depths, thus increasing primary productivity in and around seamounts. In addition, seamounts trap these organic and inorganic materials horizontally by promoting a closed circulation pattern of water like Taylor caps [37].
Given the nutrient richness and association of seamounts with hydrothermal vents, microbial growth and activity is thought to occur via biofilm formation. However, limited study has been conducted on the microbial diversity of the seamounts showing the presence of microbial mats belonging to the bacterial classes Zetaproteobacteria, Epsilonproteobacteria, Gammaproteobacteria and Deltaproteobacteria, along with archaeal phylums such as Crenarchaeota, Euryarchaeota, with the predominant microorganisms belonging to iron-, hydrogen- and sulfur-oxidizing communities. Some sulfur reducers and anaerobic methanogens have also been detected [38]. Mariprofundus ferrooxydans is a Gram-negative, neutrophilic, chemoautotrophic, and Fe-oxidizing bacterium, a new genus and species among the newly discovered Zetaproteobacteria class of Proteobacteria isolated from the microbial mat of the Loihi seamount [39].
Ice-Covered Marine Environments
The surface of the marine environment is largely covered with ice, revealing various microorganisms from the marine environment that occur at the interface between the ice and the water. Numerous species belonging to the genus Psychroflexus, Psychromonas antarcticus, Iceobacter, Polaromonas, Polibacter etc. have been unearthed from these ice-covered regions [9]. These organisms are termed as psychrophilic and have adapted and evolved over the years to function in these extreme temperatures, reaching as low as -10°C e.g. Psychromonas ingrahamii [40]. Because water density is high in these regions, the overall diffusion rate of nutrients and energy is low, suggesting that psychrophiles are oligotrophic in nature [41]. These organisms have high importance for energy-saving industrial applications based on low temperatures, as the biomolecules and the organism itself are stable in these environments.
Biotechnological and Biotransformation Potential of Marine Bacteria
Given the diversity of habitat and bacterial communities present, the marine environment has been considered a compelling source of biocatalysts and other biomolecules with applications in various biotransformation processes [2]. Several novel bacteria and numerous molecules, from enzymes to metabolites, have been isolated from this environment [42]. These bacteria and the compounds find their application in industry, healthcare, and bioremediation processes [2], making the marine environment a prime candidate for bioprospecting.
Enzymes
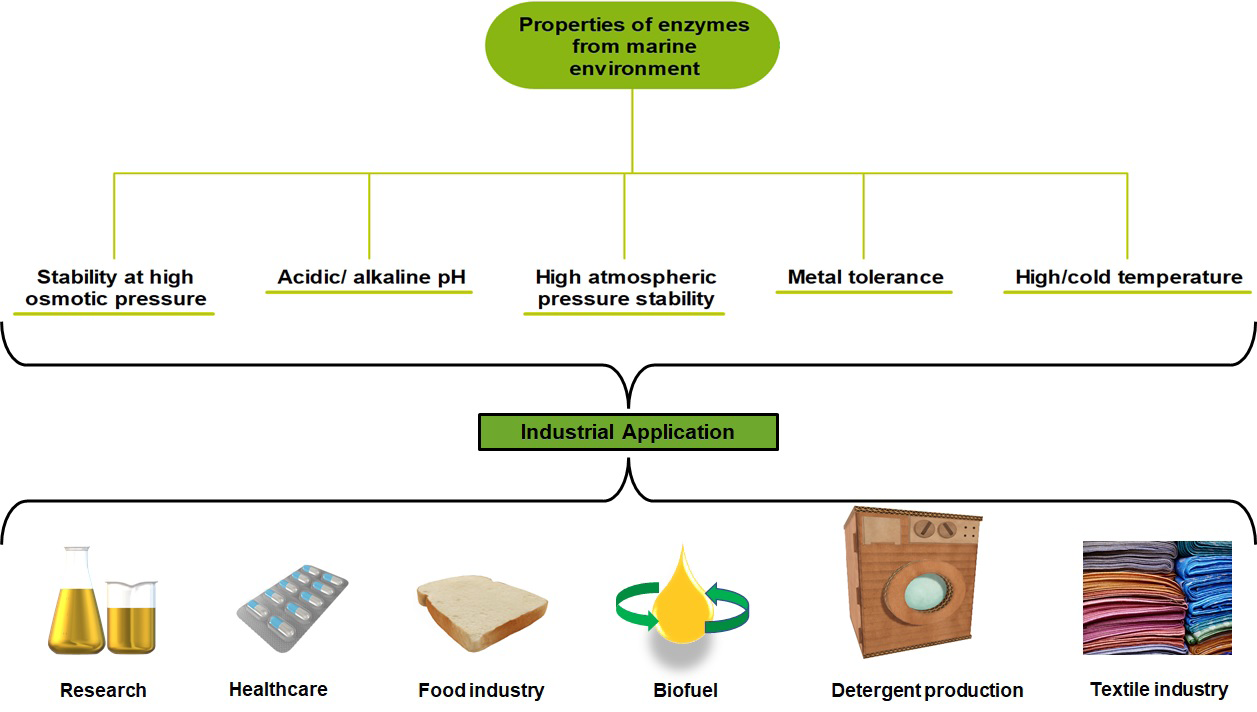
Enzymes are essential biocatalysts that carry out several biochemical processes for the proper functioning of a cell, thus sustaining life [43]. Their catalytic efficiency, environmental friendliness, and economic advantages have made them a suitable alternative to chemical catalysts for industrial processes. From food and agriculture to textiles, pharmaceuticals, biofuel production as well as for bioremediation, all sectors have witnessed an increase in the use of these enzymes [44]. Considering the possibilities of using low-cost and readily available materials along with wastes as substrates for enzymatic biotransformation, their use is of high economic and environmental benefit [45]. The demand and application of these industrial enzymes are progressing steadily and it is estimated that they will reach a global market value of $7.0 billion by 2023 [46]. As extremely diverse physiological conditions prevail in the marine environment, marine bacterial species have evolved accordingly through physiochemical and genetic manipulations, with one prominent feature being the production of highly stable enzyme systems. These enzymes have the ability to function under extreme stress conditions like high temperature or freezing cold, alkaline or acidic pH, and osmotic and hydrostatic pressure. Thus, these extreme enzymes have high potential as biocatalysts in various industrial processes [47] (Figure 1).
Hydrolase enzymes are the most commonly isolated and industrially used class of enzymes derived from marine microorganisms. Lipase is among the highly esteemed enzymes for various industrial purposes and there are numerous reports of marine bacteria producing this enzyme. A marine Janibacter sp. strain HTCC2649 produced mono- and di-acylglycerol lipase enzyme with an optimum of pH 7 at 30 °C. However, the enzyme was still able to maintain 50% of its enzyme activity at a temperature as low as 5 °C and was stable to detergents and various metal ions [48]. Similarly, Lailaja and Chandrasekaran reported a bacterial isolate Bacillus smithii BTMS 11 from marine sediments that produced an alkaline and thermostable lipase with temperature and pH stability in a wide range of 30-80 °C and 7-10, respectively. Moreover, 90% of enzyme activity was retained after treatment with different detergents for almost 3 h [49]. Marine-based lipase has several advantages over traditional mesophilic enzymes, such as its high tolerance to alkaline conditions, detergents, and cold- active/thermostable properties, which make it a preferred candidate for formulating detergents for cold or high temperature washes. Their stability also finds application in pharmaceutical, food, and paper industries [50].
Amylase is another enzyme with a high industrial value that can be obtained from almost all sources and habitats. However, marine-derived amylases exhibit exceptional properties, with marine isolates Nocardiopsis sp. 7326 [51], Bacillus sp. dsh19-1 [52], Wangia sp. C52 [53], Pseudoalteromonas sp. 2-3 [54] exhibiting cold-active amylase. Similarly, pH-tolerant [55], thermostable [56], and salt-tolerant amylase [57] have been isolated from marine bacteria, most of them showing stability towards more than one extreme [58]. Protease is used for cleaning purposes in detergent and leather industries, along with other applications in food, health, and waste management [59]. Several marine bacteria have been used for protease isolation such as Pseudoalteromonas sp. 129-1 [60], Bacillus flexus APCMST-RS2P [61], Pseudomonas sp 145-2 [62] to name a few. Agarase and chitinase enzymes are also isolated from marine bacteria as these polysaccharides are abundant in the marine environment and microorganisms use them as a carbon source [63]. Cellulase, ligninase, esterase, laccase, gelatinase, etc. are other enzymes isolated from the marine environment [64].
Bioactive Compounds
Bioactive molecules and compounds are metabolites derived from living organisms in all aspects of life that have health-promoting properties. Plants have long been considered as a major source of the various bioactive compounds produced as secondary metabolites [65]. Extensive studies have already been carried out on terrestrial life forms to discover and identify the properties of various metabolites. Therefore, other environments are now being explored to find novel bioactive compounds for various health-related benefits, with the marine ecosystem being a prominent option [66]. In addition to the enzyme, marine bacteria produce various bioactive molecules such as antioxidants, antimicrobials, antitumor compounds, etc. to adapt to and utilize the resources available in these environments (Table 1).
| Bacteria | Source | Bioactive compound | Application | Reference |
|---|---|---|---|---|
| Streptomyces sp. VITSJK8 | Cheyyur beach sediment | 1, 2- benzene dicarboxylic acid, mono 2- ethylhexyl ester | Cytotoxicity against human cancer cells | [67] |
| Streptomyces sp. MN41 | Caspian Sea sediment | Pyrrole-derivative | Antimicrobial activity against MRSA Antitumor activity | [68] |
| Streptomyces griseorubens DSD069 | 20–30 m deep Marine sediments | Bisanhydroaklavinone & 1-Hydroxybisanhydroaklavinone | Antibacterial activity | [69] |
| Nocardiopsis sp. HB-J378 | Marine sponge | Nocardiopsistins A-C | Anti-MRSA activity | [70] |
| Streptomyces bingchenggensis ULS14 | Lagos Lagoon sediments | ULDF4 & ULDF5 | Antitumor activity | [71] |
| Saccharomonospora sp. KCTC-19160 | - | Saccharochlorines A & B | BACE1 inhibitors | [72] |
| Streptomyces sp. OUCMDZ-3434 | Marine algae Enteromorpha prolifera | Wailupemycin J & (R)- wailupemycin K | Antiviral activity | [73] |
| M. thermotolerans SCSIO 00652 | South China Sea sediment | Marthiapeptide A | Antimicrobial & anticancer activity | [74] |
| Aequorivita sp. | Antarctic water sediments | Unidentified | Antimicrobial & anthelmintic activity | [75] |
| Erythrobacter sp. | Marine sponge Callyspongia aerizusa | Bromophycolide compound | Antimicrobial activity against Mycobacterium smegmatis | [76] |
| Aerococcus Uriaeequi | Yellow Sea | Exopolysaccharide A | Antioxidant activity | [77] |
| Janthinobacterium spp. strains ZZ145 & ZZ148 | Marine soil | Janthinopolyenemycins A & B | Antifungal activity | [78] |
| Halomonas sp. strain GWS-BW-H8hM (DSM 17996) | Wadden sea water | 3-(4′-hydroxyphenyl)-4- phenylpyrrole-2,5-dicarboxylic acid & 3,4-bis(4′-hydroxy- phenyl) pyrrole-2,5-dicarboxylic acid. | Antitumor activity | [79] |
Table 1: Marine bacteria & their bioactive metabolites.
Since the discovery of cephalosporin C from marine fungi, several other antimicrobial compounds have been isolated from bacteria from the marine environments [80]. A survey of the antibiotics derived from the marine based microorganism showed that 50% of the bacteria isolated from this environment had antibiotic activity against various microorganisms [81]. In a study, fifteen bacteria belonging to the genera Bacillus and Virgibacillus were found to possess antimicrobial activity against the pathogenic Vibrio parahaemolyticus and Staphylococcus aureus strains associated with foodborne illness. The strains were isolated from various marine life forms such as algae, oysters, crabs, and marine sediments [82]. The bioactive metabolite extract prepared from the actinobacteria Streptomyces sp. S2A with three major components, viz. Pyrrolo[1-a] pyrazine-1,4- dione, Hexahydro-3-(2-methylpropyl)- was found to possess inhibit the growth of bacterial strains Escherichia coli, Staphylococcus aureus, Klebsiella pneumoniae, Staphylococcus epidermidis, Bacillus cereus, and Micrococcus luteus, and the fungi Bipolaris maydis and Fusarium moniliforme. The extract also showed antioxidant and cytotoxic activity with inhibitory activity against α-amylase and α-glucosidase enzymes as major components [83]. The analogs of proxicin and polyketide abyssomicin produced by the marine bacterium Verrucosispora sp. MS100137 showed antagonistic activity against the influenza a virus [84].
The bioactive metabolites from marine bacteria may also be considered as chemotherapeutic agents for the treatment of tumors and cancer and numerous antitumor compounds have also been isolated from the marine microorganisms [85]. The depsipeptide thiocoraline derived from the marine Micromonospora sp. has high potential as an anticancer treatment because it inhibits DNA polymerase-α activity [86]. A new metabolite dentigerumycin E was discovered from marine Streptomyces sp. when the strain was co-cultured with Bacillus sp. The metabolite exhibited antimetastatic activity and antiproliferative activity against human carcinoma cells [87]. Other bioactive compounds reported from marine bacteria include antioxidants [77], anti-inflammatory compounds [88], antidiabetic compounds [89], etc.
Bioremediation
Marine bacteria, in addition to producing biocatalysts and bioactive compounds, play an essential part in the bioremediation of this water system. Urbanization and increasing human anthropogenic activities have led to the enrichment of the marine environment with pollutants in the form of plastics, microplastics, heavy metals, chemicals, sewage, oil, and petroleum [90]. Common approaches to bioremediation by marine bacterial and associated species include biofilm production, production of enzymes, biosurfactants, and oxidation of heavy metal ions [4]. Table 2 lists the marine bacteria involved in bioremediation processes.
Plastics are one of the major contaminants in the environment and their recalcitrant nature makes their biodegradation an extremely slow process. The marine bacteria produce PHB depolymerase, lipase, peroxidase, PEG dehydrogenase, polyurethanases, monooxygenase MHETase, PETase, and various other enzymes that attack and degrade plastics [103]. The actinomycete Streptomyces sp. SM14 associated with the marine sponge was isolated and a PETase-like enzyme SM14est was identified that was capable of degrading polycaprolactone [104]. Delacuvellerie et al. through their research analysis concluded that marine microbial biofilms with bacterial component caused the degradation of plastic polymers, indicated by a percentage decrease in molecular weight, through colonization and biofilm formation followed by secretion of various enzymes [105]. Another microbial consortium of Vibrio alginolyticus and Vibrio parahaemolyticus isolated from marine sediments was very effective in degrading PVA-LLDPE [106].
| Microorganism | Source | Bioremediation | Reference |
|---|---|---|---|
| Lactobacillus plantarum MF042018 | Alexandrian Mediterranean Seacoast | Biosorption of nickel & chromium | [91] |
| Vibrio T-1.3, Altermonas BP-4.3 & Cobetia S-237 | Dichlorination of Polyvinyl chloride | [92] | |
| L. boronitolerans MH2 | Johor Strait seawater | Trichloroacetic acid (TCA) degradation | [93] |
| Alteromonas macleodii, Pseudovibrio denitrificans, Pseudovibrio ascidisceicola, | Red Sea sponge Theonella swinhoei | Arsenic reduction | [94] |
| Pseudomonas chengduensis PPSS-4 | marine sediment of Paradip Port, Odisha | Sequestration of Pb(II), Cr(VI), & Cd(II) | [95] |
| Alcaligenes faecalis LNDR-1 | Sea water bank | Polyethylene degradation | [96] |
| Bacillus algicola (003-Phe1), Rhodococcus soli (102-Na5), Isoptericola chiayiensis (103-Na4), & Pseudoalteromonas agarivorans (SDRB-Py1) | Taean beach sediments | Biosurfactants production to desorbed crude oil from marine water & use it as carbon source | [97] |
| Nocardioides OK12 | Beach plastisphere | Biofilm based degradation of poly(3-hydroxybutyrate) | [98] |
| Bacillus sp. JY14 | Marine soil | Degradation of polyhydroxyalkanoates | [99] |
| Nocardiopsis sp. GRG 3 (KT235642) | Marine brown algae | Bioflocculant production & sorption of Cd, Cr, Pb, & Hg | [100] |
| Streptomyces sp. CuOff24 | Coastal sediments | Biosorption of radionuclide of Sr2+ | [101] |
| Nocardiopsis sp. 13H | Coast sediment around nuclear power plant site | extracellular polymeric substance associated adsorption of cesium | [102] |
Table 2: Marine bacteria in bioremediation.
The polyaromatic hydrocarbons (PAH) in the marine environments are highly insoluble and marine microorganisms cause their degradation via aerobic or anaerobic processes with reduction of oxygen, nitrate, or sulfates acting as electron acceptors. In rare cases, Fe (II) or Mn (III) may also act as a terminal electron acceptor. The production of biosurfactants by marine bacteria also helps in the degradation of insoluble materials by increasing the accessibility of microorganisms [90, 107]. Petroleum refining and transportation are the common reason for the pollution of marine environment by oil spills. Marine microorganisms interact and cause its degradation by producing lipase enzymes and use it as a carbon source. Three marine thermophilic bacterial species Geobacillus sp. D7, Anoxybacillus geothermalis D9 and Geobacillus sp. D4, were found to produce lipase enzyme which had an oil degradation efficiency of 17.3%, 13.1%, and 12.1% respectively [108].
Alcanivorax borkumensis produces a lipase enzyme which along with other hydrocarbon degrading enzymes in a crude preparation showed efficient degradation of petroleum contaminants (6000 ppm) at 88.52%, thus showing potential for bioremediation of hydrocarbon contamination [109]. These organisms also produce biosurfactants that help in the emulsification of oil and have been directly linked to crude oil degradation [107]. Further Bacillus cereus UCP 1615 isolated from petroleum contaminated marine sample was found to produce anionic biosurfactant which showed to effectively remove the toxicity caused by the oil spills with a survival rate of 90% for Poecilia vivipara [110].
Heavy metals such as cobalt, cadmium, lead, etc. are highly toxic and their accumulation and entry into the food chain can have serious effects [111]. Since all sewage is dumped into the marine environment, they have high levels of these heavy metals and act as their reservoir. The bacterial species in these environments have adapted and cause the removal of these heavy metals through biomineralization, biotransformation, biosorption, bioaccumulation, etc [90]. It has been reported that bacterial species of Enterobacter, Flavobacterium, Bacillus, Pseudomonas, and Micrococcus sp. can absorb these heavy metals as they have a high surface to volume ratio and teichoic acid is present as a cellular component that attracts the positively charged heavy metals [112]. Biofilms can also remove heavy metals, either by sorption or by releasing exopolymers that act as biosurfactants [113]. A comparative study by Priyadarshanee and Das using Pseudomonas chengduensis showed that a biofilm was more efficient in metal removal compared to planktonic cells [95]. Studies on biosurfactants have shown high efficiency in removing heavy metals [114] and finally, biotransformation is used to reduce toxicity by oxidation of these heavy metals as an approach of bioremediation by marine bacteria [115].
Biotechnological Advances in Marine Bioprospecting
Marine bioprospecting has been a research focus for many years, investigating the potential of marine-derived substances for commercial use in a variety of areas, including food and pharmaceuticals. Various culture- independent methods, omics technologies and genetic engineering approaches forms the backbone of such studies, and recent improvements in these methods have created better opportunities to cement marine biotechnology in an industrial setting.
Culture-Independent Gene-Targeted Methods of Marine Microbial Bioprospecting
The higher ground of revolutions and innovations in marine microbial bioprospection has been provided with culture independent methods. Conventional methods of bioprospection provide set of limitations for unculturable marine microbial communities [116]. Only 30 bacterial divisions out of more than 100 proposed bacterial divisions include the cultivable representatives [117]. Marine microbial community stands at the top for unculturable microbes applying conventional methods [118]. Hence strategies to gain in depth knowledge of unculturable microbial communities are essential in the discovery of novel function and bioprospecting [119]. Among various culture independent methods, molecular phylogeny of 16S rRNA continues to be the most widely applied in studies. This phylogenetic approach is often considered important in research framework because of two critical properties of 16S rRNA gene. The two critical properties includes: (i) presence in all life forms, and (ii) presence of domain structure with variable rates of evolution, thus allowing phylogenetic reconstruction at different levels [120]. A better efficiency to classical phylogenetic methods was brought by 16S rRNA gene hypervariable region large-scale sequencing [121]. 16S rRNA gene sequencing can be utilized in phylogenetically driven marine bio-discoveries [122]. Species chemical diversity and taxonomic diversity are coupled due to importance of secondary metabolism in specialization hence community analysis by 16S rRNA sequencing can be applied for the selection of most diverse sampling site improving recovery of novel taxa in a specific culture [123].
A complementary view on phylogenetic driven marine bio-discoveries can be achieved through functional gene- based approaches. Such approaches focus centrally on the microbial communities holding potential to carry out desired function. The marine environment has been studied extensively to demonstrate number of microbial genes encoding enzymes central in environmental processes. Functional gene-based approaches has been an integral part of such marine research framework and has contributed towards demonstration of several marine microbial enzymes participating in nitrogen fixation [124], sulphate reduction [125], hydrocarbon biodegradation [126], denitrification [127], ammonia oxidation [128] etc.This approach is highly focused nature and applies primers with variable specificities thus making this approach powerful. Functional gene-based biological tools were applied in a study to demonstrate bacterial hydrocarbon degradation in marine environment. Diversity of alkane degrading bacterial population in sediments of chronically polluted subantarctic region was studied using alkane monooxygenase (alkB) gene library analysis. Genes described in Actinobateria and Proteobacteria were affiliated to sequences obtained from sediment clone libraries with amino acid identity of 67% to sample sequences. The alkB gene sequence distribution among samples from different sites helped in identification of ecologically significant alkB genes in Subantarctic region and their further application as biomarkers in alkane degradation studies. 16S rRNA amplicon pyrosequencing helped in indicating several microbial genera in which alkB was previously not described such as Thalassospira, Oleisspira etc. This study demonstrated abundance of genera associated with oil biodegradation that was not previously studies such as Maribius, Gillisia, Bizionia, Robiginitomaculum and Spongiibacter [126]. However functional gene-based approaches bear certain limitations such as insufficient database sequence information for different functional genes with respect to 16S rRNA genes, inaccuracy in taxonomic assignment due to lateral gene transfer etc [129].
The goal of linking unculturable microbe phylogenetic identity to its function can be possibly fuelled further by another culture independent gene-targeted method called labelled isotopic-based methods. Results from such studies can provide unprecedented opportunities to develop insights into ecosystems and microbial communities and help in identification of biotechnologically relevant microorganisms [130]. The method involves radioactive labelling of substrates which allows easy identification of microbial communities that incorporate the labelled substrates. Labelled isotopic-based methods when combined with identification tools, they can be applied for discovery of unknown microbes in both functional and phylogenetic regards [131]. Stable isotope probing (SIP) is one of the most widely applied approaches, in which substrates labelled with stable isotope are utilized and assimilated by microbial populations and heavier components are integrated into cell components. The lighter and heavier components are physically separated and then analyzed [132]. DNA-SIP approach proceeds by DNA separation in caesium chloride and then separation and analysis by sequencing and cloning [133]. The RNA-SIP focuses on RNA molecule itself maintaining sequence-based DNA-SIP phylogenetic resolution. RNA-SIP provides with high turnover number independent of cell replication and high copy number [134]. Webster, et al. [135] conducted a SIP comparison DNA (16S rRNA and dsrA genes) and phospholipid fatty acids (PLFA) for studying functional diversity of prokaryotes in sulphate reducing marine sediment enrichment slurries. Different substrates such as acetate, glucose and pyruvate were labelled with 13C at low concentration of 100μM. Over the incubation period of 7 days, DGGE profiles of PFLA and 16S rRNA showed limited changes indicating presence of stable bacterial community. PFLA were rapidly labelled in 13C labelled glucose slurry indicating presence of active glucose-utilizing population in the sample. The study further concluded the presence of Desulfosarcina and Desulfococcus sulphate-reducing bacteria (SRB) and SRB related sequences in 13C-acetate- DNA dsrA gene library.
Hence study demonstrated application of DNA and PFLA-SIP to develop in-depth knowledge on carbon flow as well as active microbial population in marine environments. SIP provides certain limitations such as lack of radiolabeled substrate availability and associated health concerns. Besides SIP, fluorescence in-situ hybridization also allows for identification and detection of microbial cells using rRNA gene probes [136]. When combined with microautoradiography (MAR), FISH (FISH-MAR) provides the possibility of direct observation of labelled substrate incorporation into microbial cells [137]. Hence culture-independent gene-targeted methods can provide larger opportunities for marine microbial bioprospecting especially for previously unknown and possibly novel representatives.
Omics Approaches in Marine Bioprospecting
Advent of “omics” technology has revolutionized biotechnological fields and contributed to numerous innovations towards a sustainable scientific foundation. The application of omics approaches in marine biotechnology has produced a wealth of insights into the genetic and cellular processes in marine organisms. Such approaches represent a breakthrough in the study of marine diversity and its potential for bioprospecting [138]. The introduction of multi- omics techniques such as metagenomics, metaproteomics, metatranscriptomics and metabolomics strengthens marine biotechnology-based research, expanding its scope for efficient marine applications [139] (Table 3).
Evaluation of biotechnological potential of marine microorganisms can be successfully carried out using genetic sequencing and annotation [153]. Genome-guided marine bioprospecting methods have shown promising results in developing insights in biosynthesis of metabolite from marine microbe, thus discovery of bioactive substances [154]. Development of advanced software such as Antibiotic & Secondary Metabolite Analysis Shell (antiSMASH) has allowed efficient genome guided detection of antibiotics and other metabolites produced from different microbes [155]. This software has been applied in discovery of marine derived phosphoglycolipids, type I, II and III polyketides (PKs), bacteriocins, aminoglycosides, lantibiotics, aminocumarins, oligosaccharide antibiotics, ectoines, homoserine lactones etc. [156]. Genome-guided methods were applied in identification of a secondary metabolite produced by marine actinomycete, Salinispora tropica. Production of large number of natural PKs were discovered, particularly PK-Non-ribosomal peptide (NRP) hybrid (salinosporamide A), an anticancer drug [157]. Genome sequencing has been applied in characterization of marine Streptomyces cryptic genes, which has allowed identification of several important bioactive compounds [158]. For instance, enediyne antibiotic identification from marine S. coelicolor and S. avermitilis [158], biosynthesis of 4-Z-annimycin, polyenoic acid and 4-E-annimycin from S. calvus [159]. DNA sequencing of marine microorganisms provides new information about marine species and ecological niche [160].
Genome sequencing of three different strains of the marine cyanobacteria Prochlorococcus (SS120, MED4, and MIT9313) was performed by Rocap, et al. The study concluded that the strains belonged to same species, with a slight difference in genome size: MIT9313 had a genome size of 2.4 Mb, while SS120 and MED4 had a genome size of 1.7 Mb. Such a genomic study provided sufficient insight into the stability and environmental adaptation of marine microbial strains [161]. Such studies establish a better understanding of marine microbial growth and adaptation which are necessary for their commercial application in biotechnological functions. Sequence-based screening analysis requires specially designed primers for polymerase chain reaction (PCR) that bind to conserved sequences on DNA. The method also involves cloning the metagenomic library into a DNA vector and transforming it into a production cell. The resulting clones can then be screened for the desired sequences [162]. Metagenomic and single- cell approaches were used to discover Entotheonella, a fellow inhabitant of the marine sponge Theonella swinhoei. PCR primers specifically designed to bind genes expressing the desired cellular signaling pathways were used to detect Entotheonella DNA. This study revealed that the Entotheonella microbiome contributes very significantly to the production of peptides and polyketides in the chemically and microbially rich T. swinhoei sponge [163]. Metagenomic studies have been successfully used in novel gene discovery [164]. In such studies, the environmental DNA sample is fragmented and then cloned into conventional vectors. The DNA fragments can be sequenced, and over thousands of genes can be analyzed without screening. Such analysis may reveal several new genes with unknown and potential functions [165]. Two strategies are used in such research studies: Binning and Assembly. Binning compares reference data to assign metagenomic sequence to a taxonomic group, whereas assembly generates longer sequences for assembling sequence reads.
| Omics technology | Marine organisms studied | Techniques employed | Results concluded | Applications for human benefit | Reference |
|---|---|---|---|---|---|
| Meta-genomics | Candidatus Endobugula sertula | End sequencing and sequencing by primer walking | Identification of putative gene bryostatin polyketide synthase (PKS) gene cluster encoding active bryostatin. | Anti-cancer activity | [140] |
| Thermoproteaceae and Acidilobaceae | Function based and sequence-based techniques | Identification of heat-active β-glucosidase (Bgl 1) of archaeal origin with higher activity towards cellobiose, lactose and cellotriose, | Applicable in harsh industrial processes. | [141] | |
| Halobacillus salinus | 16S rRNA gene sequencing | Identification of secondary metabolites such as phenethylamide inhibiting quorum-sensing mediated functions in gram-negative bacteria | Antimicrobial potential | [142] | |
| Actinobacteria | Terminal-restriction fragment length polymorphism (T-RFLP), Denaturing gradient gel electrophoresis (DGGE), Thermal-GGE (TGGE) | Discovery and Identification of marine actinomycetes derived antileishmanial drugs | Prevention of leishmania | [143] | |
| Meta- transcriptomics | Mucor racemosus CBMAI 847, Marasmiellus sp. CBMAI 1062, Bacillus subtilis CBMAI 707, and Dietzia maris CBMAI 705 | Illumina HiSeq 2500 | Analysis of marine microbial consortium for Remozol Brilliant Blue R (RBBR) detoxification and decolourization | potential for bioremediation processes of textile effluents. | [144] |
| Thermotoga sp. | 454 GS-FLX sequencing | Bioprospecting of thermostable glycoside hydrolase (GH) having higher lignocelluloses degradation efficiency. | Lignocellulosic biomass degradation | [145] | |
| Meta-proteomics | Nematostella vectensis | Classical Chromatography | Identification of marine invertebrate equivalent of Fibroblast growth factor (FGF) in humans | Medical treatment | [146] |
| Haliotis laevigata | Gas chromatography (GC) coupled to Mass spectrometry (MS) | Analysis of marine invertebrate equivalent of Insulin like growth factor (IGF) 1 and 2 in humans. | Medical treatment | [147] | |
| Amphimedon queenslandica | Chromatography techniques | Marine invertebrate equivalent of Transforming growth factor β (TGF-β) | Medical treatment | [148] | |
| Gracilaria gracilis | Gas chromatography (GC) coupled with Mass spectrometry (MS) | Identification and extraction of R-phycoerythrin together with other valuable products such as arachidonic acid (omega-PUFA). | Relevant in nutraceticals, pharmaceuticals and biotechnological applications. | [149] | |
| Metabolomics | Aspergillus terreus | Multivariate analysis of Liquid chromatography/Mass spectrometry (MS) data | Isolation and identification of 7-desmethylcitreoviridin | Antifungal potential | [150] |
| Integrated omics approaches | |||||
| Metagenomics, Metaproteomics, Metabolomics | Candidatus Endoecteinascidia frumentensis | Shotgun sequencing, BLASTX and TBLASTN approaches, LC-FTIR- MS, LC-MS/MS | Identification and characterization of ET-743 or trabectedin biosynthetic pathway having tetrahydroisoquinoline functions. | Strong anticancer activities | [151] |
| Metagenomics, Metaproteomics, Metabolomics | Haliclona simulans (Bacillus subtilis MMA7) | PCR, Reverse Phase High Performance Liquid Chromatography (RP-HPLC), MALDI- TOF MS, BLASTP and BLASTN, Tandem mass spectrometry analysis (MS2). | Discovery of subtilomycin, a novel lantibiotic | Antimicrobial and medical treatment potential | [152] |
Table 3: Application of omics technology for marine bioprospecting.
Assembly allows reads to be easily subjected to bioinformatics analysis [166]. Genomic sequence markers such as k-mer can be used to identify species or strains [167]. The sequences are then subjected to clustering and bioinformatics approaches for gene prediction and functional annotation [168]. The complexity of metagenomic approaches can be reduced by metatranscriptomics, which focuses only on metabolically active genes and functions [169]. Metatranscriptomics involves sequencing the entire transcriptome and analyzing it using bioinformatics tools. It provides an excellent alternative to building a marine reference database that goes beyond genetic modeling [170]. Marine Microbial Eukaryotic Transcriptome Sequencing Project (MMETSP) is dedicated towards creation of over
650 functionally annotated, assembled as well as easily accessible transcriptomes, providing an important database for the integration of eukaryotic microbes into marine ecology [171]. The complexity in culturing marine lineages and poorly characterized molecular data make it difficult for MMETSP to cover all eukaryotic groups [172]. Several research-based studies have been conducted to analyse metatranscriptome in marine prokaryotic groups thus providing further opportunities of marine bioprospecting at broader range [173]. Metatranscriptomics has been very successful in studies analyzing marine microbial symbiosis and the utilization of certain complex compounds such as methyl phosphonate (mPn) by marine microorganisms [174]. 16S rRNA amplicon sequencing and metatranscriptomics have been used to analyze mutualism among seagrasses: Zostera marina, Z. japonica, and associated microbiomes. Metatranscriptome sequencing helped in identifying microbial genes involved in denitrification, indole-3- acetate synthesis, sulfur oxidation, agarase production, methanol consumption, etc. The study thus provided the first metatranscriptome-based evidence that the seagrass microbiome mediates multiple functions for the benefit of its host [175].
However, greater insight into the biological processes of the marine microbiome can be gained when the results of metagenomics and metatranscriptomics are complemented with data from metaproteomics analysis [176]. Marine metaproteomics provides a better understanding of how marine microbes play significant role in biogeochemical cycles [177]. Metaproteomics strategies include: sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS- PAGE) combined with MALDI-TOF mass spectrometry (MS) or SDS -PAGE combined with electrospray ionization source tandem MS (ESI-MS/ MS), another approach is liquid chromatography (LC) combined with electrospray ionization source tandem MS (LC -ESI- MS /MS) [178]. Metaproteomics has been successfully used in studies analyzing marine organic matter (OM) and microbial communities from specific marine habitats. One study examined the response of bacterioplankton to diatom blooms in North Sea. Taxonomically differentiated expression of carbohydrate- active enzymes and mechanisms of phosphorus acquisition were observed, indicating dynamic population succession at the genus level among Gammaproteobacteria, Bacteroidetes, and Alphaproteobacteria. These microbial groups were found to be highly specialized in successive degradation of algae OM [179]. Similarly, a metaproteomic study of Olavius algarvensis (marine gutless worm) and associated symbiotic microbial community was conducted by Kleiner, et al. which revealed several unusual pathways of energy and carbon utilization: (a) microbial assimilation of host waste products such as malate, acetate, succinate and propionate, (b) utilization of carbon monoxide and hydrogen as energy sources, (c) expression of high-affinity transporters, (d) energy-efficient pathways of sulfate reduction and carbon dioxide fixation [180].
To characterize metabolites and metabolic responses of marine microbes, another “omics” approach is being pursued, metabolomics. This unique approach can be successfully used to measure metabolites and identify biological markers [181]. Nuclear magnetic resonance (NMR) and mass spectroscopy (MS) are widely used methods to perform metabolomics studies. A metabolomics study was conducted to explore the chemical space of marine natural products. An untargeted LC-MS/ MS based metabolomic approach coupled with molecular networks was used to analyze the chemicals associated with 1000 marine microbes. In the study, 76 families of molecules were annotated including valinomycin, desferrioxamine E, actinomycin D etc. which are clinically very significant. In addition, a structural inventory of two new molecules, maridric acids A and B, has been performed [182].
Integrated omics approach is another widely used approach to establish solid grounds for marine bioprospecting at biotechnological levels [183]. Such approaches may combine different omic technologies together for better analysis and identification of marine organisms. Integrated omics approach was utilized by Kersten, et al. for investigation and characterization of glycosylated bioactive compounds from marine Streptomyces sp. SPB74 and S. arenicol CNB-527. Liquid chromatography (LC)-Mass spectrometry (MS); metaproteomics was applied for glycosylated natural product analysis. The researchers analysed N- and O-glycosyl groups in sugrar moieties by MSn analysis. Genome sequence identification ws conducted by antiSMASH software; metagenomics. BLAST was used for determining function of glycosylation genes. Such techniques helped to conclude characterization of cinerubin B (anticancer agent) gene cluster from Streptomyces sp. SBP74 and arenimycin B (antibiotic) gene cluster from S. arenicola CNB-527 [184]. Therefore, the omics approach is a very relevant field of technology that ensures a better future for marine biotechnology through new innovations and explorations in the still largely undiscovered world of the oceans
Genetic Engineering of Marine Microorganisms
Genetic manipulation is a powerful and extremely important technology in studies to evaluate the biotechnological potential of microorganisms. Genetic engineering (GE) offers a breakthrough in the development of microbial strains with improved industrial functions. Various studies have been carried out showing higher relevance of genetic engineering techniques in marine biotechnology fields. Major applications of genetic engineering in marine biotechnology include exploration of undiscovered sources of genetic information involved with the environmental sample after screening metagenomic libraries expressed in culture dependent marine hosts [185], furthermore, investigation of desired gene expression and function in culturable marine hosts and evaluation of their biotechnological potential [186]. Various methods have been used in studies, such as heterologous expression of the marine metagenome and recent CRISPR/Cas9-mediated genome manipulation (Figure 2).
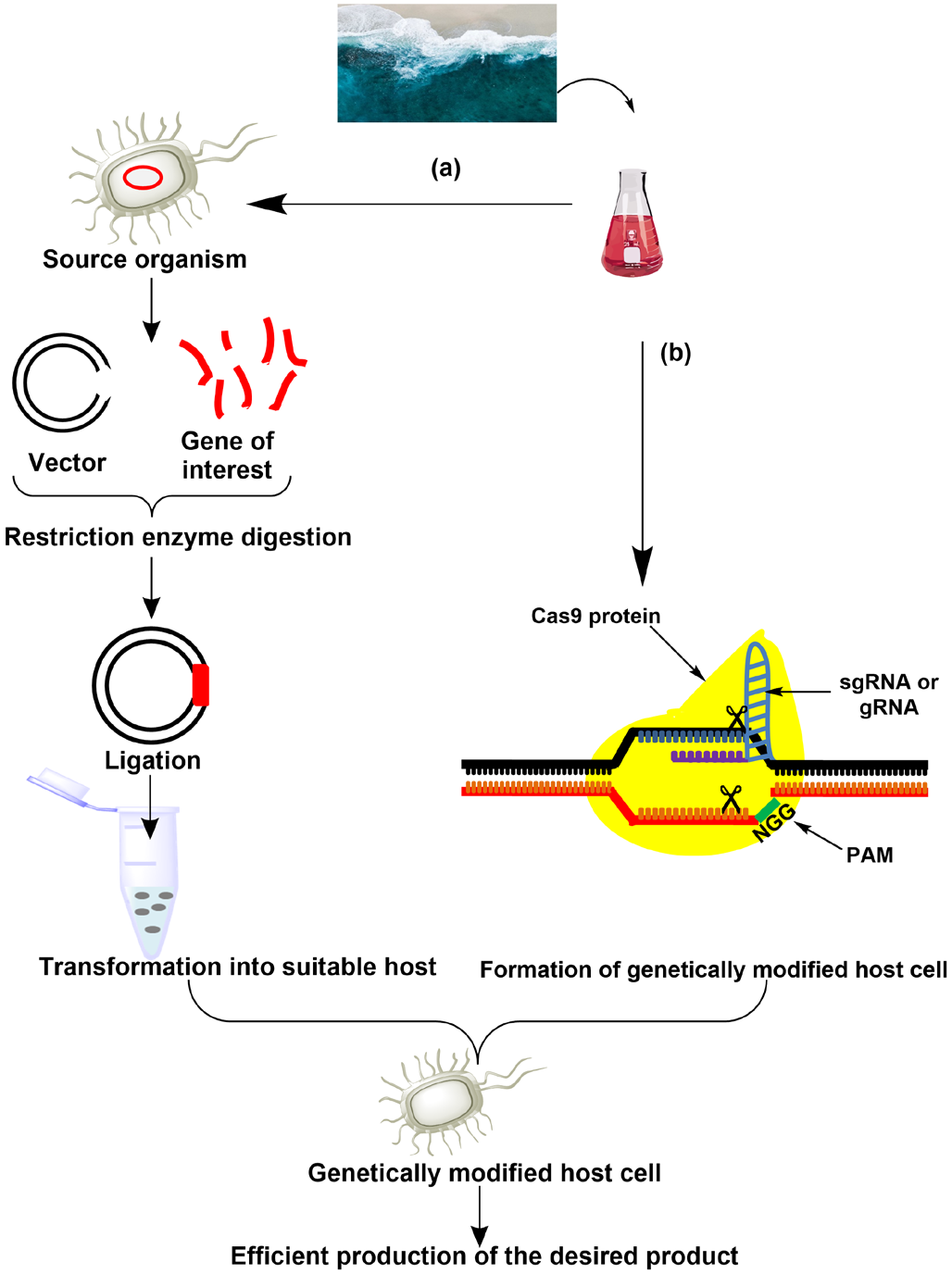
Figure 2: Genetic engineering approaches: (a) heterologous gene expression i.e., the expression of the desired gene into a non-native host using recombinant DNA technology. (b) CRISP/Cas9 mediated gene modification which employs the use of Cas9 nuclease enzyme complexed with gRNA to mediate double stranded DNA breaks at the desired site of genome allowing existing genes to be removed or new genes to be added.
There are several mechanisms to introduce external genetic material into a suitable host, such as conjugation, electroporation [187], chemical transformation [188], etc. Conjugation-mediated genetic manipulation was performed on a nonpathogenic marine bacterium Vibrio natriegens (previously classified as Pseudomonas natriegens and Beneckea natriegens) which was isolated from salt marsh mud. V. natriegens was conjugated to Escherichia coli S17- 1 λpir. Allelic exchange in V. natriegens was mediated by establishing pDM4, a sacB-based suicide vector system. This method was used to perform chromosomal deletion of several genes: dldh for D-lactate dehydrogenase, lldh for L-lactate dehydrogenase, pflfor pyruvate formate lyase, &mdh for malate dehydrogenase. The engineered strains showed higher anaerobic biosynthesis of alanine with a total volume productivity of 0.56+/-0.10 g alanine l-1 min-1. This study made V. natriegens a marine bacterium with high biotechnological potential [189]. A similar overproduction of the antimicrobial nucleoside antibiotic A201A from the marine actinomycete Marinaactinospora thermotolerance SCSIO 00652 was performed by Zhu, et al. using E. coli ET12567/pUZ8002 for intergeneric conjugation. Whole- genome sequencing and annotation methods were used to locate the gene sequences encoding A201A and the gene that inhibits its synthesis, i.e. mtdF. The inhibitory gene was inhibited using the λ-red-mediated PCR targeting method. The resulting mutant strains showed 25-fold higher A201A synthesis compared with wild-type strains [190].
Heterologous gene expression has always been an integral part of microbial genetic engineering-based approaches. The GE approach provides an excellent way to mediate expression of desired gene in a suitable and efficient host cell that does not naturally possess that gene. In this method, the desired gene fragment is inserted into a vector using recombinant DNA technology (RDT), which is then transformed into a suitable host cell [191]. This method has been used in studies with marine microorganisms and their bioprospecting. Heterologous expression of cryptomaldamide-synthesizing biosynthetic gene cluster (BGC) from the marine cyanobacterium Moorena producensstrain JHB in Synechococcus elongatus PCC7942 and Anabaena (Nostoc) PCC7120 was performed. Large BGCs are involved in the synthesis of enzymes such as non-ribosomal peptide synthetases, polyketide synthetases and other hybrid pathways which helps in the production of various bioactive compounds including cryptomaldamide with pharmaceutical potential. An Ananbaena strain containing homologous sequences to S. elongatus for chromosomal recombination at neutral site was developed. After CRISPR-based manipulation, segregated recombinant Anabaena double clones were obtained. Using genetic engineering, 28.7 kb of BGC from the marine cyanobateria Moorena (Moorea) producens JHB was expressed into host cells. High titers of cryptomaldamide were obtained from the engineered Anabaena strains compared with S. elongatus [192].
One of the recent breakthroughs in the field of genetic engineering is genetic manipulation mediated by Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)/ CRISPR-associated protein (Cas9). Originally derived from endogenous adaptive immunity in bacteria and archaea, CRISPR/Cas9 is a highly orthogonal and very versatile technology that promotes efficient research studies in the fields of systems biology, synthetic biology, and metabolic engineering. The CRISPR system contains a CRISPR precursor RNA (pre-crRNA) that is in complex with a transactivating CRISPR RNA (tracrRNA). The RNA hybrid is recognized by the enzyme RNase III, which generates the mature crRNA-tracrRNA hybrid. The mature crRNA combines with Cas9 and the dual RNA: Cas9 complex is directed to the DNA site. Cas9 mediates RNA-directed DNA double-strand endonuclease cleavage to generate DNA double-strand breaks (DSBs) [193]. In genetic manipulation studies, crRNA is replaced by a specially designed short guide RNA (sgRNA) that mediates site-specific DSB formation. DSBs are repaired by homologous DNA repair or non-homologous end-joining repair pathways that mediate gene knock-in or gene knock-out (indels) [194]. CRISPR/ Cas9 technology is a new and unique genetic engineering approach in marine biotechnology. Several studies have shown the great potential of this technology in the emerging fields of marine biotechnology and bioprospecting. In a study investigating genes involved in fatty acid biosynthesis, Xia et al. successfully used the CRISPR/Cas9 approach in conjunction with the lamda red recombinase system to copy genes involved in fatty acid metabolism from the marine bacterium Shewanella frigidimarina into E. coli [195]. Thus, various studies demonstrate that genetic engineering provides a solid foundation for better and more efficient marine biotechnological fields.
Climate Change and Impacts on Marine Biodiversity
The marine environment and its associated bacterial and other microbial species are not only a source of potential bioactive compounds and bioremediation, but also play an important role in global climate change. Climate change is having a huge impact on marine life forms, causing a shift and loss of biodiversity which has an impact also on the bacterial communities of this environment. Numerous studies already show the impact of climate change on marine biodiversity in terms of larger life forms such as fish, corals, mammals and plankton. However, studies to understand the impact and use of marine microorganisms to mitigate climate change are now gaining attention [196]. Global warming has led to melting of ice, which affects salinity. Increased ocean water temperature affects the water and thus indirectly affects the survival of marine life forms, favoring only those that can adapt to these changes [197]. In addition, high temperature also favors algal and cyanobacterial blooms, directly affecting nutrient distribution. Increased CO2 in the atmosphere is absorbed by the ocean, which turns into acid, decreasing the pH of seawater. All this affects mineral and nutrient distribution and increases the reproductive rate of microbes, leading to nutrient scarcity and consequent extinction of microbial communities, which in turn affects the marine food web and marine biodiversity [198].
Conversely, the marine microbes including bacteria can help mitigate the problem of global warming and climate change. The marine microbes, which account for nearly 98% of the biomass of the ocean, are the main source of oxygen and at the same time can trap the greenhouse gasses from the atmosphere to be used in their metabolic processes. Therefore, marine microbes have the potential to mitigate the negative effects of global warming and climate change. The ocean ecosystem contains nearly 93% of the world’s CO2 cycled by marine bacteria and archaea, i.e., about 90 billion tons of CO2 per year, of which 60 billion tons are released by anthropogenic activities. The large population of photosynthetic microorganisms, especially in the Polar Regions, plays a key role in maintaining the fraction of anthropogenic CO2 in the atmosphere [199]. In addition, the metabolic activity of marine bacteria such as respiration of organic matter contributes to the CO2 released in the atmosphere. The methane gas released from the seabed and the marine methanogenic microbes have the potential to increase the concentration of greenhouse gasses in the environment. However, the methanotrophic bacteria and archaea found in these deep-sea systems scavenge and use them to neutralize their effect [199]. Thus, the marine environment has an impact on the geochemical cycle of greenhouse gasses and thus on global climate change.
Conclusion and Future Perspective
The marine environment covers a vast area with diverse habitats, which makes it a diversity hotspot for isolating novel organisms, both eukaryotic and prokaryotic. Therefore, the marine environment has a high bio-perspective for obtaining novel enzymes and other biomolecules especially from the bacterial species considering their dynamic ability to adapt to various extremities. As much of the marine environment remains unexplored due to various technological challenges, the advancement in science and technology can help to reach these areas of the environment while the accessible marine habitats are being studied using the classical and modern omic technologies for the development and improvement of marine-derived bacteria and derived metabolites for various and commercial, environmental and health support.
Statements and Declarations
Competing Interests
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This work was supported by “University Research Fund Award 2018” by Sikkim University to AKV.
Author Contributions Statement
AKV conceived of the presented idea. DC and AKV discussed every topic and wrote the review. AKV provided critical feedback, supervised the study, and helped in shape the review.
Acknowledgement
The authors would like to thank the Department of Microbiology, Sikkim University, for providing the computational infrastructure and central library facilities for procuring references and plagiarism analysis (URKUND: Plagiarism Detection Software).
References
-
Lozupone CA, Knight R (2007) Global patterns in bacterial diversity. Proc Natl Acad Sci USA 104(27): 11436-11440.
-
Rotter A, Barbier M, Bertoni F, Bones AM, Cancela ML, et al. (2021) The Essentials of marine biotechnology. Front in Mar Sci 8: 629629.
-
Bryson M, Ferrari R, Figueira W, Pizarro O, Madin J, et al. (2017) Characterization of measurement errors using structure‐from‐motion and photogrammetry to measure marine habitat structural complexity.Ecol Evol 7(15): 5669-5681.
-
Baharum SN, Beng EK, Mokhtar MAA (2010) Marine microorganisms: potential application and challenges. Journal of Biological Sciences 10(6): 555-564.
-
Munn CB (2020) Microbial aspects of marine biofouling, biodeterioration, and pollution. In: Munn CB (Ed.), Marine Microbiology. 3rd(Edn.), CRC Press, pp: 31.
-
Barzkar N, Jahromi ST, Poorsaheli HB, Vianello F (2019) Metabolites from marine microorganisms, micro, and macroalgae: immense scope for pharmacology. Mar Drugs 17(8): 464.
-
Pettit RK (2011) Culturability and secondary metabolite diversity of extreme microbes: expanding contribution of deep sea and deep-sea vent microbes to natural product discovery. Mar Biotechnol (NY) 13(1): 1-11.
-
Javed F, Qadir MI, Janbaz KH, Ali M (2011) Novel drugs from marine microorganisms. Crit Rev Microbiol 37(3): 245-249.
-
Aryal S, Karki G, Pandey S (2015) Microbial diversity in freshwater and marine environment. Nepal J Biotech 3(1): 68-70.
-
Galéron J (2014) Deep-sea environment. In: Fouquet Y, Lacroix D (Eds.), Deep Marine Mineral Resources. Springer, pp: 41-54.
-
Anderson TR, Rice T (2006) Deserts on the sea floor: Edward Forbes and his azoic hypothesis for a lifeless deep ocean. Endeavour 30(4): 131-137.
-
Roberts CM (2002) Deep impact: the rising toll of fishing in the deep sea. Trends in ecology & evolution 17(5): 242-245.
-
Kaiser MJ, Attrill MJ, Jennings S, Thomas D, Barnes KA, et al. (2011) Marine ecology: processes, systems, and impacts. 2nd(Edn.), Oxford University Press.
-
Costello MJ, Breyer S (2017) Ocean depths: the mesopelagic and implications for global warming. Current Biology 27(1): R36-R38.
-
Smith CR, Leo FCD, Bernardino AF, Sweetman AK, Arbizu PM (2008) Abyssal food limitation, ecosystem structure and climate change. Trends in Ecology & Evolution 23(9): 518-528.
-
Durden JM, Bett BJ, Jones DOB, Huvenne VAI, Ruhl HA (2015) Abyssal hills–hidden source of increased habitat heterogeneity, benthic megafaunal biomass and diversity in the deep sea. Progress in Oceanography 137: 209-218.
-
Ebbe B, Billett DSM, Brandt A, Ellingsen K, Glover A, et al. (2010) Diversity of abyssal marine life. Life in the World’s Oceans: Diversity, Distribution, and Abundance, edited by: McIntyre A, pp: 139-160.
-
Hoffmann KC, Bienhold C, Buttigieg PL, Knittel K, Pérez RL, et al. (2020) Diversity and metabolism of Woeseiales bacteria, global members of marine sediment communities. The ISME journal 14(4): 1042-1056.
-
Aylward FO, Santoro AE (2020) Heterotrophic Thaumarchaea with small genomes are widespread in the dark ocean. mSystems 5(3): e00415-00420.
-
Glazer AN (2013) Natural reserves and preserves. 2nd(Edn.), Encyclopedia of Biodiversity, pp: 442-450.
-
Canganella F (2001) Hydrothermal vent ecosystems and representative hyperthermophilic microorganisms. Annals of microbiology 51(1): 11-27.
-
Dick GJ (2019) The microbiomes of deep-sea hydrothermal vents: distributed globally, shaped locally. Nat Rev Microbiol 17(5): 271-283.
-
González JM, Masuchi Y, Robb FT, Ammerman JW, Maeder DL, et al. (1998) Pyrococcus horikoshii sp. nov., a hyperthermophilic archaeon isolated from a hydrothermal vent at the Okinawa Trough. Extremophiles 2(2): 123-130.
-
Bae SS, Kim YJ, Yang SH, Lim JK, Jeon JH, et al. (2006) Thermococcus onnurineus sp. nov., a hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent area at the PACMANUS field. Journal of microbiology and biotechnology 16(11): 1826-1831.
-
Takai K, Nakamura K, Toki T, Tsunogai U, Miyazaki M, et al. (2008) Cell proliferation at 122 C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc Natl Acad Sci 105(31): 10949-10954.
-
Allioux M, Jebbar M, Slobodkina G, Slobodkin A, Moalic Y, et al. (2021) Complete genome sequence of Thermosulfurimonas marina SU872T, an anaerobic thermophilic chemolithoautotrophic bacterium isolated from a shallow marine hydrothermal vent. Marine Genomics 55: 100800.
-
Cheng JH, Wang Y, Zhang XY, Sun ML, Zhang X, et al. (2021) Characterization and Diversity Analysis of the Extracellular Proteases of Thermophilic Anoxybacillus caldiproteolyticus 1A02591 From Deep- Sea Hydrothermal Vent Sediment. Front Microbiol 12: 643508.
-
Baumann LMF, Taubner RS, Bauersachs T, Steiner M, Schleper C, et al. (2018) Intact polar lipid and core lipid inventory of the hydrothermal vent methanogens Methanocaldococcus villosus and Methanothermococcus okinawensis. Organic Geochemistry 126: 33-42.
-
Alain K, Marteinsson VT, Miroshnichenko ML, Osmolovskaya EAB, Prieur D, et al. (2002) Marinitoga piezophila sp. nov., a rod-shaped, thermo-piezophilic bacterium isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int J Syst Evol Microbiol 52(pt 4): 1331-1339.
-
Takai K, Miyazaki M, Hirayama H, Nakagawa S, Querellou J, et al. (2009) Isolation and physiological characterization of two novel, piezophilic, thermophilic chemolithoautotrophs from a deep‐sea hydrothermal vent chimney. Environ Microbiol 11(8): 1983-1997.
-
Wang S, Jiang L, Hu Q, Liu X, Yang S, et al. (2021) Elemental sulfur reduction by a deep‐sea hydrothermal vent Campylobacterium Sulfurimonas sp. NW10. Environ Microbiol 23(2): 965-979.
-
Jiang L, Lyu J, Shao Z (2017) Sulfur metabolism of Hydrogenovibrio thermophilus strain S5 and its adaptations to deep-sea hydrothermal vent environment. Front Microbiol 8: 2513.
-
Grassle JF (2013) Marine Ecosystems.
-
Rogers A (2019) Threats to seamount ecosystems and their management. 2nd(Edn.) World seas: an environmental evaluation, Elsevier 3: 427-451.
-
Wessel P, Sandwell DT, Kim SS (2015) The global seamount census. Oceanography 23(1): 24-33.
-
Busch K, Hanz U, Mienis F, Mueller B, Franke A, et al. (2020) On giant shoulders: How a seamount affects the microbial community composition of seawater and sponges. Biogeosciences 17(13): 3471-3486.
-
Roberts EM, Mienis F, Rapp HT, Hanz U, Meyer HK, et al. (2018) Oceanographic setting and short-timescale environmental variability at an Arctic seamount sponge ground. Deep Sea Research Part I: Oceanographic Research Papers 138: 98-113.
-
Emerson D, Moyer CL (2015) Microbiology of seamounts: common patterns observed in community structure. Oceanography 23(1): 148-163.
-
Emerson D, Rentz JA, Lilburn TG, Davis RE, Aldrich H, et al. (2007) A novel lineage of proteobacteria involved in formation of marine Fe-oxidizing microbial mat communities. PLoS One 2(7): e667.
-
Auman AJ, Breezee JL, Gosink JJ, Kämpfer P, Staley JT (2006) Psychromonas ingrahamii sp. nov., a novel gas vacuolate, psychrophilic bacterium isolated from Arctic polar sea ice. Int J Syst Evol Microbiol 56(pt 5): 1001- 1007.
-
Poli A, Finore I, Romano I, Gioiello A, Lama L, et al. (2017) Microbial diversity in extreme marine habitats and their biomolecules. Microorganisms 5(2): 25.
-
Zotchev SB (2012) Marine actinomycetes as an emerging resource for the drug development pipelines. J Biotechnol 158(4): 168-175.
-
Kim SK (2019) Marine Microorganism Resources and Biotechnology. Essentials of Marine Biotechnology, Springer, pp: 381-415.
-
May O (2019) Industrial enzyme applications–overview and historic perspective. Industrial enzyme applications, pp: 1-24.
-
Böttcher H, Soltmann U, Mertig M, Pompe W (2004) Biocers: ceramics with incorporated microorganisms for biocatalytic, biosorptive and functional materials development. J Mater Chem 14(14): 2176-2188.
-
Fasim A, More VS, More SS (2021) Large-scale production of enzymes for biotechnology uses. Current Opinion in Biotechnology 69: 68-76.
-
Zhang C, Kim SK (2010) Research and application of marine microbial enzymes: status and prospects. Marine drugs 8(6): 1920-1934.
-
Yuan D, Lan D, Xin R, Yang B, Wang Y (2014) Biochemical properties of a new cold-active mono-and diacylglycerol lipase from marine member Janibacter sp. strain HTCC2649. Int J Mol Sci 15(6): 10554-10566.
-
Lailaja V, Chandrasekaran M (2013) Detergent compatible alkaline lipase produced by marine Bacillus smithii BTMS 11. World J Microbiol Biotechnol 29(8): 1349-1360.
-
Navvabi A, Razzaghi M, Fernandes P, Karami L, Homaei A (2018) Novel lipases discovery specifically from marine organisms for industrial production and practical applications. Process biochemistry 70: 61-70.
-
Zhang JW, Zeng RY (2008) Purification and characterization of a cold-adapted α-amylase produced by Nocardiopsis sp. 7326 isolated from Prydz Bay, Antarctic. Mar Biotechnol 10(1): 75-82.
-
Dou S, Chi N, Zhou X, Zhang Q, Pang F, et al. (2018) Molecular cloning, expression, and biochemical characterization of a novel cold-active α-amylase from Bacillus sp. dsh19-1. Extremophiles 22(5): 739-749.
-
Liu J, Zhang Z, Dang H, Lu J, Cui Z (2011) Isolation and characterization of a cold-active amylase from marine Wangia sp. C52. African J Microbiol Res 5(10): 1156- 1162.
-
Sanchez AC, Ravanal MC, Andrews BA, Asenjo JA (2019) Heterologous expression and biochemical characterization of a novel cold-active α-amylase from the Antarctic bacteria Pseudoalteromonas sp. 2-3.” Protein expression and purification 155: 78-85.
-
Kalpana BJ, Pandian SK (2014) Halotolerant, acid‐alkali stable, chelator resistant and raw starch digesting α‐ amylase from a marine bacterium Bacillus subtilis S8–18. J Basic Microbiol 54(8): 802-811.
-
Liu S, Ahmed S, Fang Y (2019) Cloning, expression and characterization of a novel α-amylase from Salinispora arenicola CNP193. Protein J 38(6): 716-722.
-
Wu G, Qin Y, Cheng Q, Liu Z (2014) Characterization of a novel alkali-stable and salt-tolerant α-amylase from marine bacterium Zunongwangia profunda.” Journal of Molecular Catalysis B: Enzymatic 110: 8-15.
-
Ding N, Zhao B, Ban X, Li C, Prasad BVV, et al. (2021) Carbohydrate-Binding Module and Linker Allow Cold Adaptation and Salt Tolerance of Maltopentaose-Forming Amylase from Marine Bacterium Saccharophagus degradans 2-40T. Front Microbiol 12: 708480.
-
Barzkar N (2020) Marine microbial alkaline protease: an efficient and essential tool for various industrial applications. International Journal of Biological Macromolecules 161: 1216-1229.
-
Wu S, Liu G, Zhang D, Li C, Sun C (2015) Purification and biochemical characterization of an alkaline protease from marine bacteria Pseudoalteromonas sp. 129‐1. J Basic Microbiol 55(12): 1427-1434.
-
Maruthiah T, Esakkiraj P, Immanuel G, Palavesam A (2014) Alkaline serine protease from marine Bacillus flexus APCMST-RS2P: purification and characterization. Current Biotechnology 3(3): 238-243.
-
Makino K, Koshikawa T, Nishihara T, Ichikawa T, Kondo M (1981) Studies on protease from marine bacteria. 1 Isolation of marine Pseudomonas sp 145-2 and purification of protease. Microbios 31(124): 103-112.
-
Roseline TL, Sachindra NM (2018) Purification and Characterization of Agarase from Marine Bacteria Acinetobacter sp. PS12B and Its Use for Preparing Bioactive Hydrolysate from Agarophyte Red Seaweed Gracilaria verrucosa. Appl Biochem Biotechnol 186(1): 66-84.
-
Rao TE, Imchen M, Kumavath R (2017) Marine enzymes: production and applications for human health. Adv Food Nutr Res 80: 149-163.
-
Patil BS, Jayaprakasha GK, Murthy KNC, Vikram A (2009) Bioactive compounds: historical perspectives, opportunities, and challenges. J Agric Food Chem 57(18): 8142-8160.
-
Jensen PR, Fenical W (2000) Marine microorganisms and drug discovery: current status and future potential. Drugs from the Sea, pp: 6-29.
-
Krishnan K, Mani A, Jasmine S (2014) Cytotoxic activity of bioactive compound 1, 2-benzene dicarboxylic acid, mono 2-ethylhexyl ester extracted from a marine derived Streptomyces sp. VITSJK8. International journal of molecular and cellular medicine 3(4): 246-254.
-
Norouzi H, Khorasgani MR, Danesh A (2019) Anti-MRSA activity of a bioactive compound produced by a marine Streptomyces and its optimization using statistical experimental design. Iran J Basic Med Sci 22(9): 1073- 1084.
-
Paderog MJV, Suarez AFL, Sabido EM, Low ZJ, Saludes JP, et al. (2020) Anthracycline shunt metabolites from Philippine marine sediment-derived Streptomyces destroy cell membrane integrity of multidrug-resistant Staphylococcus aureus. Front Microbiol 11: 743.
-
Xu D, Nepal KK, Chen J, Harmody D, Zhu H, et al. (2018) Nocardiopsistins AC: New angucyclines with anti- MRSA activity isolated from a marine sponge-derived Nocardiopsis sp. HB-J378. Synth Syst Biotechnol 3(4): 246-251.
-
Bolorunduro OFD, Adeleye IA, Akinleye MO, Wang PG (2019) Anticancer potential of metabolic compounds from marine actinomycetes isolated from Lagos Lagoon sediment. Journal of Pharmaceutical Analysis 9(3): 201- 208.
-
Le TC, Katila N, Park S, Lee J, Yang I, et al. (2020) Two new secondary metabolites, saccharochlorines A and B, from a marine bacterium Saccharomonospora sp. KCTC- 19160. Bioorg Med Chem Lett 30(11): 127145.
-
Liu H, Chen Z, Zhu G, Wang L, Du Y, et al. (2017) Phenolic polyketides from the marine alga-derived Streptomyces sp. OUCMDZ-3434. Tetrahedron 73(36): 5451-5455.
-
Zhou X, Huang H, Chen Y, Tan J, Song Y, et al. (2012) Marthiapeptide A, an anti-infective and cytotoxic polythiazole cyclopeptide from a 60 L scale fermentation of the deep sea-derived Marinactinospora thermotolerans SCSIO 00652. J Nat Prod 75(12): 2251-2255.
-
Esposito FP, Ingham CJ, Ortiz RH, Bizet C, Tasdemir D, et al. (2018) Isolation by miniaturized culture chip of an Antarctic bacterium Aequorivita sp. with antimicrobial and anthelmintic activity. Biotechnology Reports 20: e00281.
-
Murniasih T, Cindhy CI, Sukara E, Yunovilsa MP (2020) Anti-mycobacterium compound derived from Erythrobacter sp. isolated from Callyspongia aurizusa. IOP Conf Ser: Earth Environ Sci IOP Publishing 429: 012009.
-
Wang C, Fan Q, Zhang X, Lu X, Xu Y, et al. (2018) Isolation, characterization, and pharmaceutical applications of an exopolysaccharide from Aerococcus Uriaeequi. Marine drugs 16(9): 337.
-
Anjum K, Sadiq I, Chen L, Kaleem S, Li XC, et al. (2018) Novel antifungal janthinopolyenemycins A and B from a co-culture of marine-associated Janthinobacterium spp. ZZ145 and ZZ148. Tetrahedron Letters 59(38): 3490- 3494.
-
Wang L, Groβe T, Stevens H, Brinkhoff T, Simon M, et al. (2006) Bioactive hydroxyphenylpyrrole-dicarboxylic acids from a new marine Halomonas sp.: Production and structure elucidation. Appl Microbiol Biotechnol 72(4): 816-822.
-
Ameen F, AlNadhari S, Al-Homaidan AA (2021) Marine microorganisms as an untapped source of bioactive compounds. Saudi J Biol Sci 28(1): 224-231.
-
Wiese J, ImhoffJF (2019) Marine bacteria and fungi as promising source for new antibiotics. Drug Dev Res 80(1): 24-27.
-
Silva LG, Villarreal JMI, Garza ZJM (2018) Bacillus and Virgibacillus strains isolated from three Mexican coasts antagonize Staphylococcus aureus and Vibrio parahaemolyticus. FEMS Microbiol Lett 365(19): fny202.
-
Siddharth S, VittalRR (2018) Evaluation of antimicrobial, enzyme inhibitory, antioxidant and cytotoxic activities of partially purified volatile metabolites of marine Streptomyces sp. S2A. Microorganisms 6(3): 72.
-
Zhang J, Li B, Qin Y, Karthik L, Zhu G, et al. (2020) A new abyssomicin polyketide with anti-influenza A virus activity from a marine-derived Verrucosispora sp. MS100137. Appl Microbiol Biotechnol 104(4): 1533- 1543.
-
Prachyawarakorn V, Mahidol C, Sureram S, Sangpetsiripan S, Wiyakrutta S, et al. (2008) Diketopiperazines and phthalides from a marine derived fungus of the order Pleosporales. Planta Med 74(1): 69-72.
-
Newman DJ, Cragg GM (2004) Marine natural products and related compounds in clinical and advanced preclinical trials. J Nat Prod 67(8): 1216-1238.
-
Shin D, Byun WS, Moon K, Kwon Y, Bae M, et al. (2018) Coculture of marine Streptomyces sp. with Bacillus sp. produces a new piperazic acid-bearing cyclic peptide. Front Chem 6: 498.
-
Mayer AMS, Rodríguez AD, Berlinck RGS, Fusetani N (2011) Marine pharmacology in 2007–8: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous system, and other miscellaneous mechanisms of action. Comp Biochem Physiol C Toxicol Pharmacol 153(2): 191-222.
-
Elaiyaraja V, Latha MH, Devi CS (2018) Optimization and production of anti-inflammatory and anti-diabetic metabolites from marine Streptomyces sp. VITJS8. Research Journal of Pharmacy and Technology 11(7): 2866-2868.
-
Mohanrasu K, Rao RGR, Raja R, Arun A (2020) Bioremediation Process by Marine Microorganisms. Encyclopedia of Marine Biotechnology 4: 2211-2228.
-
Ameen FA, Hamdan AM, El-Naggar MY (2020) Assessment of the heavy metal bioremediation efficiency of the novel marine lactic acid bacterium, Lactobacillus plantarum MF042018. Scientific reports 10(1): 314.
-
Khandare SD, Chaudhary DR, Jha B (2021) Bioremediation of polyvinyl chloride (PVC) films by marine bacteria. Marine Pollution Bulletin 169: 112566.
-
Heidarrezaei M, Shokravi H, Huyop F, Koloor SSR, Petrů M (2020) Isolation and characterization of a novel bacterium from the marine environment for trichloroacetic acid bioremediation. Appl Sci 10(13): 4593.
-
Shoham S, Weinberger A, Kaplan A, Avisar D, Ilan M (2021) Arsenate reducing bacteria isolated from the marine sponge Theonella swinhoei: Bioremediation potential. Ecotoxicology and Environmental Safety 222: 112522.
-
Priyadarshanee M, Das S (2021) Bioremediation potential of biofilm forming multi-metal resistant marine bacterium Pseudomonas chengduensis PPSS-4 isolated from contaminated site of Paradip Port, Odisha. Journal of Earth System Science 130(3): 1-17.
-
Nag M, Lahiri D, Dutta B, Jadav G, Ray RR (2021) Biodegradation of used polyethylene bags by a new marine strain of Alcaligenes faecalis LNDR-1. Environ Sci Pollut Res 28(30): 41365-41379.
-
Lee DW, Lee H, Kwon BO, Khim JS, Yim UH, et al. (2018) Biosurfactant-assisted bioremediation of crude oil by indigenous bacteria isolated from Taean beach sediment. Environ Pollut 241: 254-264.
-
Suzuki M, Tachibana Y, Takizawa R, Morikawa T, Takeno H, et al. (2021) A novel poly (3-hydroxybutyrate)- degrading actinobacterium that was isolated from plastisphere formed on marine plastic debris. Polymer Degradation and Stability 183: 109461.
-
Cho JY, Park SL, Lee HJ, Kim SH, Suh MJ, et al. (2021) Polyhydroxyalkanoates (PHAs) degradation by the newly isolated marine Bacillus sp. JY14. Chemosphere 283: 131172.
-
Rajivgandhi G, Vimala RTV, Maruthupandy M, Alharbi NS, Kadaikunnan S, et al. (2021) Enlightening the characteristics of bioflocculant of endophytic actinomycetes from marine algae and its biosorption of heavy metal removal. Environmental Research 200: 111708.
-
Kamala K, Sivaperumal P, Thilagaraj R, Natarajan E (2020) Bioremediation of Sr2+ ion radionuclide by using marine Streptomyces sp. CuOff24 extracellular polymeric substances. Journal of Chemical Technology & Biotechnology 95(4): 893-903.
-
Sivaperumal P, Kamala K, Rajaram R (2018) Adsorption of cesium ion by marine actinobacterium Nocardiopsis sp. 13H and their extracellular polymeric substances (EPS) role in bioremediation. Environ Sci Pollut Res Int 25(5): 4254-4267.
-
Anjana K, Hinduja M, Sujitha K, Dharani G, Ganesh KA (2020) Review on plastic wastes in marine environment– Biodegradation and biotechnological solutions. Mar Pollut Bull 150: 110733.
-
Almeida EL, Rincón AFC, Jackson SA, Dobson ADW (2019) In silico screening and heterologous expression of a polyethylene terephthalate hydrolase (PETase)- like enzyme (SM14est) with polycaprolactone (PCL)- degrading activity, from the marine sponge-derived strain Streptomyces sp. SM14. Front Microbiol 10: 2187.
-
Delacuvellerie A, Benali S, Cyriaque V, Moins S, Raquez JM, et al. (2021) Microbial biofilm composition and polymer degradation of compostable and non- compostable plastics immersed in the marine environment. Journal of Hazardous Materials 419: 126526.
-
Raghul SS, Bhat SG, Chandrasekaran M, Francis V, Thachil ET (2014) Biodegradation of polyvinyl alcohol- low linear density polyethylene-blended plastic film by consortium of marine benthic vibrios. International Journal of Environmental Science and Technology 11(7): 1827-1834.
-
Sachaniya BK, Gosai HB, Panseriya HZ, Vala AK, Dave BP (2019) Polycyclic aromatic hydrocarbons (PAHs): occurrence and bioremediation in the marine environment. Marine Ecology: Current and Future Developments 1(32): 435-466.
-
Yusoff DF, Rahaman RNZRA, Masomian M, Ali MSM, Leow TC (2020) Newly isolated alkane hydroxylase and lipase producing Geobacillus and Anoxybacillus species involved in crude oil degradation. Catalysts 10(8): 851.
-
Kadri T, Magdouli S, Rouissi T, Brar SK (2018) Ex-situ biodegradation of petroleum hydrocarbons using Alcanivorax borkumensis enzymes. Biochemical Engineering Journal 132: 279-287.
-
Durval IJB, Mendonça AHR, Rocha IV, Luna JM, Rufino RD, et al. (2020) Production, characterization, evaluation and toxicity assessment of a Bacillus cereus UCP 1615 biosurfactant for marine oil spills bioremediation. Mar Pollut Bullet 157: 111357.
-
Glorennec P (2006) Analysis and reduction of the uncertainty of the assessment of children’s lead exposure around an old mine. Environ Res 100(2): 150-158.
-
Iyer A, Mody K, Jha B (2004) Accumulation of hexavalent chromium by an exopolysaccharide producing marine Enterobacter cloaceae. Mar Pollut Bull 49(11-12): 974-977.
-
El-Masry MH, El-Bestawy E, Nawal IE (2004) Bioremediation of vegetable oil and grease from polluted wastewater using a sand biofilm system. World Journal of Microbiology and Biotechnology 20(6): 551-557.
-
Das P, Mukherjee S, Sen R (2009) Biosurfactant of marine origin exhibiting heavy metal remediation properties. Bioresour Technol 100(20): 4887-4890.
-
Abbas SZ, Rafatullah M, Ismail N, Shakoori FR (2018) Electrochemistry and microbiology of microbial fuel cells treating marine sediments polluted with heavy metals. Rsc Advances 8(34): 18800-18813.
-
Bernard L, Schäfer H, Joux F, Courties C, Muyzer G, et al. (2000) Genetic diversity of total, active and culturable marine bacteria in coastal seawater. Aquatic Microbial Ecology 23(1): 1-1.
-
Prakash O, Shouche Y, Jangid K, Kostka JE (2013) Microbial cultivation and the role of microbial resource centers in the omics era. Applied Microbiology and Biotechnology 97(1): 51-62.
-
Heidelberg KB, Gilbert JA, Joint I (2010) Marine genomics: at the interface of marine microbial ecology and biodiscovery. Microb Biotechnol 3(5): 531-543.
-
Mincer TJ, Fenical W, Jensen PR (2005) Culture- dependent and culture-independent diversity within the obligate marine actinomycete genus Salinispora. Appl Environ Microbiol. 71(11): 7019-7028.
-
Gray JP, Herwig RP (1996) Phylogenetic analysis of the bacterial communities in marine sediments. Appl Environ Microbiol 62(11): 4049-4059.
-
Yang B, Wang Y, Qian PY (2016) Sensitivity and correlation of hypervariable regions in 16S rRNA genes in phylogenetic analysis. BMC bioinformatics 17: 35.
-
Tringe SG, Hugenholtz P (2008) A renaissance for the pioneering 16S rRNA gene. Curr Opin Microbiol 11(5): 442-446.
-
Goodfellow M, Fiedler HP (2010) A guide to successful bioprospecting: informed by actinobacterial systematics. Antonie Van Leeuwenhoek 98(2): 119-142.
-
Zehr JP (2011) Nitrogen fixation by marine cyanobacteria. Trends in microbiology 19(4): 162-173.
-
Jimenez JRP, Kerkhof LJ (2005) Phylogeography of sulfate-reducing bacteria among disturbed sediments, disclosed by analysis of the dissimilatory sulfite reductase genes (dsrAB). Applied and Environmental Microbiology 71(2): 1004-1011.
-
Guibert LM, Loviso CL, Marcos MS, Commendatore MG, Dionisi HM, et al. (2012) Alkane biodegradation genes from chronically polluted subantarctic coastal sediments and their shifts in response to oil exposure. Microb Ecol 64(3): 605-616.
-
Braker G, Rı́o HLAD, Devol AH, Fesefeldt A, Tiedje AJM (2001) Community structure of denitrifiers, Bacteria, and Archaea along redox gradients in Pacific Northwest marine sediments by terminal restriction fragment length polymorphism analysis of amplified nitrite reductase (nirS) and 16S rRNA genes. Appl Environ Microbiol 67(4): 1893-1901.
-
Park SJ, Park BJ, Rhee SK (2008) Comparative analysis of archaeal 16S rRNA and amoA genes to estimate the abundance and diversity of ammonia-oxidizing archaea in marine sediments. Extremophiles 12(4): 605-615.
-
Hickey WJ, Chen S,Zhao J (2012) The phn island: a new genomic island encoding catabolism of polynuclear aromatic hydrocarbons. Front Microbiol 3: 125.
-
Neufeld JD, Wagner M, Murrell JC (2007) Who eats what, where and when? Isotope-labelling experiments are coming of age. ISME J 1(2): 103-110.
-
Wellington EMH, Berry A, Krsek M (2003) Resolving functional diversity in relation to microbial community structure in soil: exploiting genomics and stable isotope probing. Curr Opin Microbiol 6(3): 295-301.
-
Dumont MG, Murrell JC (2005) Stable isotope probing—linking microbial identity to function. Nat Rev Microbiol 3(6): 499-504.
-
Dunford EA, Neufeld JD (2010) DNA stable-isotope probing (DNA-SIP). J Vis Exp 42: 2027.
-
Whiteley AS, Ghori NUH, Grez BM, Vuong P, Waite I, et al. (2019) RNA stable-isotope probing. Methods in Molecular Biology.
-
Webster G, Watt LC, Rinna J, Fry JC, Evershed RP, et al. (2006) A comparison of stable‐isotope probing of DNA and phospholipid fatty acids to study prokaryotic functional diversity in sulfate‐reducing marine sediment enrichment slurries. Environ Microbiol 8(9): 1575-1589.
-
Amann RI, Ludwig W, Schleifer KH (1995) Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol Rev 59(1): 143-169.
-
Ishii K, Mußmann M, MacGregor BJ, Amann R (2004) An improved fluorescence in situ hybridization protocol for the identification of bacteria and archaea in marine sediments. FEMS Microbiol Ecol 50(3): 203-213.
-
Abid F, Zahid MA, Abedin ZU, Nizami SB, Abid MJ, et al. (2018) Omics 3: Omics approaches in marine biotechnology: The treasure of ocean for human betterments. Omics Technologies and Bio-Engineering, Elsevier, pp: 47-61.
-
Daniotti S, Re I (2021) Marine Biotechnology: Challenges and Development Market Trends for the Enhancement of Biotic Resources in Industrial Pharmaceutical and Food Applications. A Statistical Analysis of Scientific Literature and Business Models. Mar Drugs 19(2): 61.
-
Sudek S, Lopanik NB, Waggoner LE, Hildebrand M, Anderson C, et al. (2007) Identification of the putative bryostatin polyketide synthase gene cluster from “Candidatus Endobugula sertula”, the uncultivated microbial symbiont of the marine bryozoan Bugula neritina. J Nat Prod 70(1): 67-74.
-
Schröder C, Elleuche S, Blank S, Antranikian G (2014) Characterization of a heat-active archaeal β-glucosidase from a hydrothermal spring metagenome. Enzyme Microb Technol 57: 48-54.
-
Teasdale ME, Liu J, Wallace J, Akhlaghi F, Rowley DC (2009) Secondary metabolites produced by the marine bacterium Halobacillus salinus that inhibit quorum sensing-controlled phenotypes in gram-negative bacteria. Appl Environ Microbiol 75(3): 567-572.
-
Bolorunduro OFD, Osuolale O, Saibu S, Adeleye IA, Aminah NS (2021) Bioprospecting marine actinomycetes for antileishmanial drugs: current perspectives and future prospects. Heliyon 7(8): e07710.
-
Vieira GAL, Cabral L, Otero IVR, Ferro M, Faria AUD, et al. (2021) Marine associated microbial consortium applied to RBBR textile dye detoxification and decolorization: Combined approach and metatranscriptomic analysis. Chemosphere 267: 129190.
-
Lyford JR (2012) Bioprospecting for thermostable glycoside hydrolases: a metatranscriptomic approach, [Honolulu]:[University of Hawaii at Manoa],[December 2012].
-
Rentzsch F, Fritzenwanker JH, Scholz CB, Technau U (2008) FGF signalling controls formation of the apical sensory organ in the cnidarian Nematostella vectensis. Development 135(10): 1761-1769.
-
Weiss IM, Göhring W, Fritz M, Mann K (2001) Perlustrin, a Haliotis laevigata (abalone) nacre protein, is homologous to the insulin-like growth factor binding protein N-terminal module of vertebrates. Biochem Biophys Res Commun 285(2): 244-249.
-
Srivastava M, Simakov O, Chapman J, Fahey B, Gauthier ME, et al. (2010) The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 466(7307): 720-726.
-
Francavilla M, Franchi M, Monteleone M, Caroppo C (2013) The red seaweed Gracilaria gracilis as a multi products source. Mar Drugs 11(10): 3754-3776.
-
Adpressa DA, Loesgen S (2016) Bioprospecting chemical diversity and bioactivity in a marine derived Aspergillus terreus. Chem Biodivers 13(2): 253-259.
-
Rath CM, Janto B, Earl J, Ahmed A, Hu FZ, et al. (2011) Meta-omic characterization of the marine invertebrate microbial consortium that produces the chemotherapeutic natural product ET-743. ACS Chem Biol 6(11): 1244-1256.
-
Phelan RW, Barret M, Cotter PD, O’Connor PM, Chen R, et al. (2013) Subtilomycin: a new lantibiotic from Bacillus subtilis strain MMA7 isolated from the marine sponge Haliclona simulans. Mar Drugs 11(6): 1878-1898.
-
Metzker ML (2010) Sequencing technologies—the next generation. Nat Rev Genet 11(1): 31-46.
-
Liu X, Bolla K, Ashforth EJ, Zhuo Y, Gao H, et al. (2012) Systematics-guided bioprospecting for bioactive microbial natural products. Antonie Van Leeuwenhoek 101(1): 55-66.
-
Medema MH, Blin K, Cimermancic P, Jager VD, Zakrzewski P, et al. (2011) AntiSMASH: rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res 39(Web Server issue): W339-W346.
-
Blin K, Medema MH, Kazempour D, Fischbach MA, Breitling R, et al. (2013) AntiSMASH 2.0—a versatile platform for genome mining of secondary metabolite producers. Nucleic Acids Res 41(Web Server issue): W204-W212.
-
Udwary DW, Zeigler L, Asolkar RN, Singan V, Lapidus A, et al. (2007) Genome sequencing reveals complex secondary metabolome in the marine actinomycete Salinispora tropica. Proceedings of the National Academy of Sciences 104(25): 10376-10381.
-
Zazopoulos E, Huang K, Staffa A, Liu W, Bachmann BO, et al. (2003) A genomics-guided approach for discovering and expressing cryptic metabolic pathways. Nat Biotechnol 21(2): 187-190.
-
Kalan L, Gessner A, Thaker MN, Waglechner N, Zhu X, et al. (2013) A cryptic polyene biosynthetic gene cluster in Streptomyces calvus is expressed upon complementation with a functional bldA gene. Chem Biol 20(10): 1214-1224.
-
Alió CP, Simó R (2002) Studying marine microorganisms from space. Int Microbiol 5(4): 195- 200.
-
Rocap G, Larimer WF, Lamerdin J, Malfatti S, Chain P, et al. (2003) Genome divergence in two Prochlorococcus ecotypes reflects oceanic niche differentiation. Nature 424(6952): 1042-1047.
-
Schloss PD, Handelsman J (2003) Biotechnological prospects from metagenomics. Curr Opin Biotechnol 14(3): 303-310.
-
Wilson MC, Mori T, Rückert C, Uria AR, Helf MJ, et al. (2014) An environmental bacterial taxon with a large and distinct metabolic repertoire. Nature 506(7486): 58-62.
-
Culligan EP, Sleator RD, Marchesi JR, Hill C (2014) Metagenomics and novel gene discovery: promise and potential for novel therapeutics. Virulence 5(3): 399- 412.
-
Venter JC, Remington K, Heidelberg JF, Halpern AL, Rusch D, et al. (2004) Environmental genome shotgun sequencing of the Sargasso Sea. Science 304(5667): 66- 74.
-
Sharpton TJ (2014) An introduction to the analysis of shotgun metagenomic data. Front Plant Sci 5: 209.
-
Tu Q, He Z, Zhou J (2014) Strain/species identification in metagenomes using genome-specific markers. Nucleic Acids Research 42(8): e67.
-
Richardson EJ, Watson M (2013) The automatic annotation of bacterial genomes. Brief Bioinform 14(1): 1-12.
-
Bashiardes S, Schapira GZ, Elinav E (2016) Use of metatranscriptomics in microbiome research. Bioinform Biol Insights 10: 19-25.
-
Gilbert JA, Hughes M (2011) Gene expression profiling: metatranscriptomics. Springer Nature Experments, pp: 195-205.
-
Johnson L, Alexander H, Brown CT (2016) Marine Microbial Eukaryotic Transcriptome Sequencing Project, re-assemblies. figshare.
-
Keeling PJ, Burki F, Wilcox HM, Allam B, Allen EE, et al. (2014) The Marine Microbial Eukaryote Transcriptome Sequencing Project (MMETSP): illuminating the functional diversity of eukaryotic life in the oceans through transcriptome sequencing. PLoS Biol 12(6): e1001889.
-
Gifford SM, Sharma S, Kanto JMR. Moran MA (2011) Quantitative analysis of a deeply sequenced marine microbial metatranscriptome. ISME J 5(3): 461-472.
-
Martínez A, Ventouras LA, Wilson ST, Karl DM, DeLong EF (2013) Metatranscriptomic and functional metagenomic analysis of methylphosphonate utilization by marine bacteria. Front Microbiol 4: 340.
-
Crump BC, Wojahn JM, Tomas F, Mueller RS (2018) Metatranscriptomics and amplicon sequencing reveal mutualisms in seagrass microbiomes. Front Microbiol 9: 388.
-
Wang DZ, Xie ZX, Zhang SF (2014) Marine metaproteomics: current status and future directions. J Proteomics 97: 27-35.
-
Dong HP, Hong YG, Lu S, Xie LY (2014) Metaproteomics reveals the major microbial players and their biogeochemical functions in a productive coastal system in the northern South China Sea. Environ Microbiol Rep 6(6): 683-695.
-
Williams TJ, Cavicchioli R (2014) Marine metaproteomics: deciphering the microbial metabolic food web. Trends Microbiology 22(5): 248-260.
-
Teeling H, Fuchs BM, Becher D, Klockow C, Gardebrecht A, et al. (2012) Substrate-controlled succession of marine bacterioplankton populations induced by a phytoplankton bloom. Science 336(6081): 608-611.
-
Kleiner M, Wentrup C, Lott C, Teeling H, Wetzel S, et al. (2012) Metaproteomics of a gutless marine worm and its symbiotic microbial community reveal unusual pathways for carbon and energy use. Proceedings of the National Academy of Sciences 109(19): E1173-E1182.
-
Idle JR, Gonzalez FJ (2007) Metabolomics. Cell Metab 6(5): 348-351.
-
Floros DJ, Jensen PR, Dorrestein PC, Koyama N (2016) A metabolomics guided exploration of marine natural product chemical space. Metabolomics 12(9): 145.
-
Schofield MM, Sherman DH (2013) Meta-omic characterization of prokaryotic gene clusters for natural product biosynthesis. Curr Opin Biotechnol 24(6): 1151- 1158.
-
Kersten RD, Ziemert N, Gonzalez DJ, Duggan BM, Nizet V, et al. (2013) Glycogenomics as a mass spectrometry-guided genome-mining method for microbial glycosylated molecules. Proceedings of the National Academy of Sciences 110(47): E4407-E4416.
-
Aakvik T, Degnes KF, Dahlsrud R, Schmidt F, Dam R, et al. (2009) A plasmid RK2-based broad-host-range cloning vector useful for transfer of metagenomic libraries to a variety of bacterial species. FEMS Microbiol Lett 296(2): 149-158.
-
Zess EK, Begemann MB, Pfleger BF (2016) Construction of new synthetic biology tools for the control of gene expression in the cyanobacterium Synechococcus sp. strain PCC 7002. Biotechnol Bioeng 113(2): 424-432.
-
Bahi MM, Tsaloglou MN, Mowlem M, Morgan H (2011) Electroporation and lysis of marine microalga Karenia brevis for RNA extraction and amplification. J R Soc Interface 8(57): 601-608.
-
Weinstock MT, Hesek ED, Wilson CM, Gibson DG (2016) Vibrio natriegens as a fast-growing host for molecular biology. Nature Methods 13(10): 849-851.
-
Hoffart E, Grenz S, Lange J, Nitschel R, Müller F, et al. (2017) High substrate uptake rates empower Vibrio natriegens as production host for industrial biotechnology. App Environ Microbiol 83(22): e01614- 01617.
-
Zhu Q, Li J, Ma J, Luo M, Wang B, et al. (2012) Discovery and engineered overproduction of antimicrobial nucleoside antibiotic A201A from the deep-sea marine actinomycete Marinactinospora thermotolerans SCSIO 00652. Antimicrob Agents Chemother 56(1): 110-114.
-
Glick BR (1995) Metabolic load and heterologous gene expression. Biotechnol Adv 13(2): 247-261.
-
Taton A, Ecker A, Diaz B, Moss NA, Anderson B, et al. (2020) Heterologous expression of cryptomaldamide in a cyanobacterial host. ACS Synth Biol 9(12): 3364-3376.
-
Wang Y, Cheng H, Liu Y, Liu Y, Wen X, et al. (2021) In- situ generation of large numbers of genetic combinations for metabolic reprogramming via CRISPR-guided base editing. Nat Commun 12(1): 678.
-
Zhao J, Fang H, Zhang D (2020) Expanding application of CRISPR-Cas9 system in microorganisms. Synth Syst Biotechnol 5(4): 269-276.
-
Xia J, Wang L, Zhu JB, Sun CJ, Zheng MG, et al. (2016) Expression of Shewanella frigidimarina fatty acid metabolic genes in E. coli by CRISPR/cas9-coupled lambda Red recombineering. Biotechnol Lett 38(1): 117-122.
-
Worm B, Lotze HK (2021) Marine biodiversity and climate change. 3rd(Edn.),Climate change, Elsevier: 445- 464.
-
Glöckner FO, Gasol JM, Stal LJ, Sandaa RA, O’Gara F, et al. (2012) Marine microbial diversity and its role in ecosystem functioning and environmental change, European Science Foundation, pp: 1-84.
-
Dutta H, Dutta A (2016) The microbial aspect of climate change. Energy, Ecology and Environment 1(4): 209-232.
-
Suttle CA (2007) Marine viruses—major players in the global ecosystem. Nat Rev Microbiol 5(10): 801-812.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania