Applications of Genomic Selection in Animal Breeding; Challenges and Opportunities
The world's population demand and animal output are significantly out of balance. Although traditional breeding techniques have been successful in selecting animal populations for a variety of traits with economic significance, the reliability of breeding value has always been in doubt. According to simulation and experiment data, genomic selection for young animals without own performance can predict breeding values with good accuracy. Genetic markers that cover the entire genome are employed in genomic selection, a sort of marker-assisted selection, to ensure that all loci for quantitative traits are in linkage disequilibrium with at least one marker. Early animal selection enables the development of innovative breeding techniques that increase genetic advancement while decreasing costs. The future of animal breeding companies lies in genomic selection, which increases genetic gain by reducing genetic interval and enhancing reliability. To regulate long-term genetic gain and increase the precision of genomic estimated value, more study is needed. An overview of the developments in genomic selection and its use in animal breeding was the goal of this paper.
Introduction
With a growing global population, there is a greater need for food production. Food production needs to double in the upcoming years in order to meet the demands of the world’s population, according to the Food and Agriculture Organization of the United Nations (FAO) [1, 2]. By enhancing genetic diversity, animal health, and animal husbandry practice, the predicted productivity gap could be closed. Intentional human selection is based on animal performance records and is related to animal breeding; selection is typically based on more than one trait [3]. Breeders have successfully produced improved animals using traditional animal breeding techniques (TAB) in the lack of molecular understanding [4]. Since ancient times, domestic species have used artificial selection based on the phenotypes of their own animals. The use of data on the phenotypes of relatives to forecast the “breeding values” of candidates for selection was made possible throughout the 20th century by selection index theory, followed by Best Linear Unbiased Prediction (BLUP a more complex approach relying on mixed linear models). This resulted in the effective selection of phenotypic variables with moderate to high heritability that is simple to measure [5]. However, efficient phenotyping required expensive phenotyping investments for traits that were hard to measure or had poor heritability. Numerous Quantitative Trait Loci (i.e., sections of the genome that account for a portion of the genetic variance of a trait) have been identified using genetic markers over the past 25 years. Quantitative genetics-based animal breeding has significantly enhanced animal productivity.
However, due to technological limitations, the use of genetic markers in breeding programmes has been very restricted [6]. Genomic selection, also known as dense panels of single nucleotide polymorphism (SNP) markers, has recently opened up new possibilities for choosing the animals that are most suitable for breeding. Utilizing the quantitative trait locus (QTL) associated with a specific phenotypic trait and using it for selection purposes is the fundamental idea behind this methodology. The extensive knowledge of dense genetic markers has improved the accuracy with which breeding values for young candidates may be estimated. In order to create prediction equations of an individual’s genetic merit, the principle of genomic selection calls for utilizing both genotypic and phenotypic data from the reference population [7]. Greater reference male or female populations are anticipated to increase the reliability of genomic selection [8]. On the other hand, progeny tested bulls frequently create the training set of genomic selection due to the trustworthy phenotypic information obtained from a large group of daughters. It has been suggested that an effective way to increase the precision of genomic selection is to use a combined reference population by gathering data from many populations [9].
The technical and financial effectiveness of animal breeding programs has greatly risen thanks to genomic selection [10], whose advantages were initially noted for dairy cattle. These anticipated benefits are often brought about by a shorter generation interval, increased EBV accuracy, and lower progeny testing expenditures [11]. These theoretical discoveries led to the fast implementation of genomic selection in dairy cattle breeding programs. The initial goal of this research was to describe the evolution of animal breeding practices from conventional approaches to genomic selection. The second goal was to provide an overview of how genomic selection has been used in various farm animals, such as ruminants, swine, and chickens.
Traditional Animal Breeding
Breeding entails choosing domestic animals with the intention of enhancing quantitative or qualitative qualities in the offspring. Animals have been improved via a variety of breeding techniques, but the basic goal has always been to increase the production of superior animals with desired features [12]. Small holder farmers in low- to middle- income countries (LMICS) typically cannot participate in traditional genetic improvement programs, which are based on measuring large numbers of pedigree-recorded animals in clearly defined cohort groups for the full range of economically significant productive and adaptive traits [13, 14]. By characterizing native and hybrid animals for use in conservation, crossbreeding, and within-breed selection programs to improve economically significant traits, the use of genomic data in conjunction with information and communication technologies now offers significant new opportunities to increase the rates of genetic gain. As causal mutations are discovered, additional technologies, such as genome editing, in combination with cutting-edge reproductive techniques that enable quick multiplication and less reliance on cold chains for the delivery of improved genetics, could potentially transform livestock breeding even further [14].
Challenges
Due to numerous significant obstacles, there has only been a limited amount of utilization of genetic technology in grazing livestock in low-middle income countries. The issues are examined in the sections that follow, along with options to eliminate them for the ruminant livestock species that are most common in those areas beef and dairy cattle, sheep, and goats. Despite the fact that the paper’s main focus is on the use of genomic selection. The BLUP technique enhances the genetic response to selection by increasing the EBV’s dependability. This is true because the technique can take into account all systematic effects (such as batch, sex, production environment, and age variation) that are frequently linked to features in farm animals that are significant from an economic standpoint. Additionally, BLUP analysis uses all pedigree information through a numerator relationship matrix to account for increases in additive genetic variance brought on by inbreeding or assortative mating, which strengthens the reliability of EBV [15].
Despite the fact that animal selection using EBV inferred from phenotype has been quite successful, there are still a few drawbacks. These largely concern the ability to frequently and promptly capture phenotypic data of chosen candidates and/or their near relatives, which aids in accelerating selection processes at a young age to shorten generational gaps. Costly phenotypic recording for features that are hard to measure or have low heritability is also crucial in this case. Another drawback of TAB is that the feature of interest, which can only be detected late in life and on one sex, necessitates animal sacrifice (for meat quality) or exposure to circumstances that would make it difficult to sell or export the animals’ germplasm (for example, disease resistance). Additionally, these breeding procedures continue to be relatively slow because it takes a long time to gather enough daughter phenotype data to accurately calculate genetic assessments [2].
Opportunities
For vast or pastoral production systems in tropical and subtropical climes, including those in Africa and Asia, there are the highest potential gains in cattle output. These systems make use of land resources that have limited other potential applications, such as urbanization. Additionally, they take advantage of ruminants’ advantages over monogastric livestock species, which include their ability to graze on pastures of inferior quality. As has happened recently in the intensive livestock industries, the pastoral livestock industries are likewise much less likely than the intensive livestock industries to encounter unequal demands on their production techniques from urban populations. By 2050, livestock farmers in the tropics and subtropics will need to adopt new, cost-effective, and transformative technologies for use with animals that are already well acclimated to their production conditions in order to double yields from the same resource base. The same way they have in the past, traditional technologies that produce incremental adjustments will help to increase production [14, 16]. For instance, one study showed that US dairy farmers now need 21% fewer cows, 23% less feed, 65% less water, and 90% less land to produce 1 billion kg of milk than they did in 1944, with a 57% reduction in methane emissions and concurrently significant waste reductions. This is possible thanks to the use of long-established technologies, such as animal breeding and animal nutrition.
Marker-assisted Selection
Marker-Assisted Selection (MAS) refers to the use of genetic information in addition to phenotypic information to improve dependability (MAS). As early as 1900, the idea of MAS for the selection of superior animals was initially proposed [17]. In MAS, marker genes are used to denote the presence of desirable genes. Trait-controlling genes are dispersed across the genome, but relatively few genes have big effects, and many more genes have steadily smaller effects [18]. A Marker assisted selection relies on determining the relationship between a genetic marker and associated Quantitative Trait Loci (QTL) [19], where the relationship between the markers is dependent on the distance between the marker and the target traits [20]. The selection response benefits from the combination of conventional breeding approaches and molecular genetic techniques. With the use of MAS, it is now possible to identify the trait of interest that, regardless of environmental circumstance, will be passed on to the future. This approach aids in identifying traits like disease resistance that have a low phenotypic expression. Because an individual’s phenotype may be anticipated at a very early stage, selection is possible for recessive genes and mutants in addition to MAS selection.
The majority of desirable qualities are controlled by a large number of loci, making the MAS approach ineffective in animal breeding programs even though it boosts animal production by boosting reliability. As a result, there may not be much genetic gain because only some of the genetic diversity can be captured by a small number of loci [21]. Additionally, the use of MAS is hampered by the computations’ complexity, which entails estimating breeding values and using information from molecular markers [22] (Figure 1).
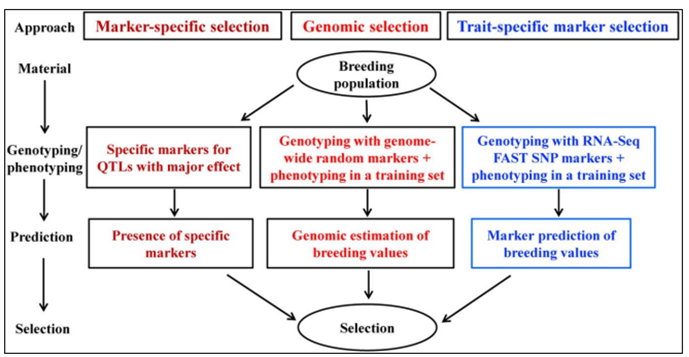
Development of Genomic Selection
For qualities with a straightforward genetic determinism, the MAS technique is successful, but it produces disappointing results in many more complex situations. The restricted genetic diversity and the weak correlation between markers and quantitative trait loci (QTL) at the population level were the two main factors contributing to this low productivity [5]. Due to the benefits of MAS in commercial breeding operations being obviously less than envisaged, another restriction of MAS is the expensive genotyping of selecting candidates. The entire genomes of numerous animals, including chicken, horse, sheep, cow, dog, cat, and rabbit, have been sequenced to date [23].
Principle of Genomic Selection
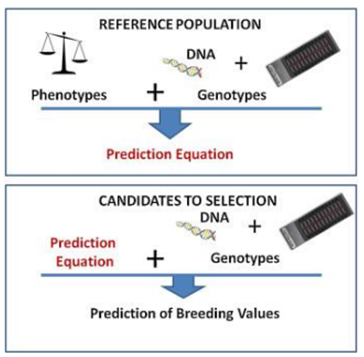
The main principle of genomic selection is the assessment of breeding value using marker data without knowledge of gene location. The gathering of phenotypic and genotypic data from the reference population is the initial step in the genomic selection process. All animals in the reference population are genotyped for SNPs across the complete genome to obtain genotypic information [5]. Having access to a sizable population of animals, such as a reference or training population, with correct phenotypes for the trait is therefore necessary for the initial phase in the genomic selection process (s). A whole-genome SNP array should also be used to genotype this population. The collected data will next be used to build a statistical model that estimates how each SNP affects the trait(s) of interest. A predictive equation to determine a genomic estimated breeding value is the end outcome (GEBV). Following a validation step, the prediction equation can be used to calculate the genomic breeding value of new animals based on their genotypes from the SNP array and in the lack of any reliable phenotypes for these animals. The population size and heritability of the characteristic under consideration affect the GEBV’s accuracy. González- Recio, et al. [24] cited a 4-fold improvement in GEBV accuracy over the parent average for feed conversion efficiency in the case of the chicken.
Three variables trait heritability (h2), the number of animals in the reference population (N), and the q parameter affect the GEBV’s accuracy [25]. When compared to conventional breeding techniques, genomic selection accelerates the pace of genetic gain, which boosts productivity (Table 1, Figure 2).
| Species | Genome Size (Gb) | Year |
|---|---|---|
| Chicken (Gallus gallus) | 1.05 | 2004 |
| Dog (Canis familiaris) | 2.4 | 2003 |
| Cattle (Bos Taurus) | 2.91 | 2009 |
| Horse (Equus caballus) | 2.47 | 2009 |
| Pig (Sus scrofa) | 2.2 | 2009 |
| Sheep (Ovis aries) | 2.78 | 2008 |
| Camel (Camelus dromedaries) | 2.2 | 2011 |
Table 1: Summary of first sequenced genomes for animal species.
Advantages of Genomic Selection
- Lowering generation interval
- Can be expanded to include any qualities identified in a reference population.
- Improved selection precision
- Intensifying the selecting process.
- Genomic selection Explains a substantially higher share of the genetic variance than MAS
Genomic Selection in Dairy Cattle
To identify the markers and enhance the qualities connected to milk production, cow health, udder health, and cow conformation, dairy cattle have undergone genomic selection. In regard to genomic selection in dairy cows, Australia is the industry leader, with the United States, Canada, and China all working on related issues. In dairy cattle, genomic prediction is more accurate than 0.8 for production traits and 0.7 for features including fertility, longevity, somatic cell count, and others [26]. Most dairy cattle are developed using a straightforward within-breed selection concept. On rare occasions, crossbred calves are born, and if no replacement heifers are required, they can be used to produce beef. Because bulls’ semen is transmitted by AI, they contribute far more to the domestic and international breeding pool than sires do. Despite the fact that breeding programs have been successful in enhancing qualities like milk output, there may be some restrictions with classical selection. Dairy cattle made a good model for the application of GS due to their increased health and fertility issues, relative importance of dairy cattle production globally, long generation interval due to the long reproductive cycle, and indirect measurement of the main traits of interest via the daughters of a bull [27].
Genomic Selection in Beef Cattle
The beef cattle sector can benefit more from genomic selection than the dairy cattle industry since beef cattle have longer generation intervals. Genomic selection is currently widely used in various breeds of beef. For instance, more than 52,000 Angus animals have been genotyped for GEBV evaluation in the United States [28]. However compared to dairy cattle, the accuracy of genetic forecasts in beef cattle has been lower. Because dairy cattle reference populations are of higher quality, the accuracy is lower. Less breed- specific animals from the reference population of beef cattle have undergone progeny testing. Also, compared to dairy cattle, the target population and validation animals may not be as closely connected to the reference population in beef cattle. The accuracy is quite poor if the target breed is not represented in the reference population. Genomic selection hasn’t been used as frequently in beef cattle breeding as it has in dairy cattle breeding, though. This is due in part to the lesser accuracy as well as the diminished economic benefits [29].
Genomic Selection in Small Ruminants
Small ruminants do not undergo extensive genomic selection, and they have lower genetic gains than dairy cattle. A very limited number of studies on genomic selection in small ruminants have been published. Recent research has examined the viability of genomic selection in small ruminants in meat sheep in Australia and New Zealand, dairy sheep in France, and dairy goats in France and the UK [30]. A reference population must be established, and animals that are genetically related to the larger population must undergo extensive phenotyping in order to connect genotypic data with phenotype. This is one of the most crucial conditions for genomic selection. In the French and UK dairy goat populations, the improvement in GEBV accuracy was 0.06 for milk yield and 0.14 for fat and protein content. The increase in accuracy was closely connected with the size of the reference population and the genomic heritability of the trait, indicating that accuracy and anticipated genetic gain may rise as reference populations grow [31] (Figure 3).
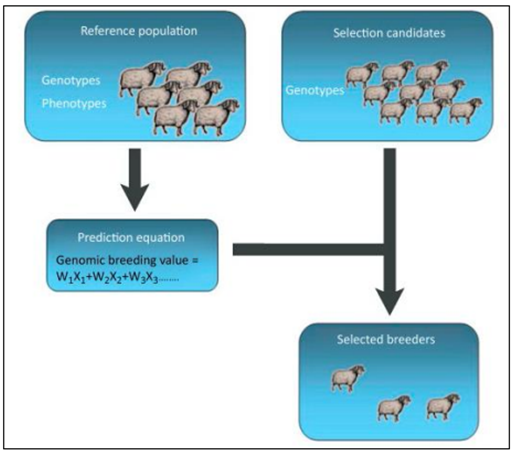
Genomic Selection in Pig
Pig’s genome became publicly released in 2009. In pigs, genomic selection can decrease generation intervals by up to 25%, increasing genetic gain and improving EBV accuracy by up to 50%, especially for traits that cannot be enhanced using conventional breeding techniques. Pigs with genomic selection had larger litters but have higher pre-weaning mortality rates [7].
Genomic Selection in Poultry
The first livestock species to be sequenced and millions of SNPs to be discovered was the chicken. Despite the fact that chickens have more than double the genetic gain compared to cattle and pigs due to the greater number of offspring they generate [32]. Yet, the genetic benefit can be further enhanced by reducing the generation gap with the use of genomic selection. Because the majority of features may be observed in both sexes at a young age, the application of genomic selection in broilers is less extensive than in layers. Furthermore, the increase in accuracy for broiler chicken features like fertility and egg production can be anywhere between 20 and 45, while it can be over 50% for traits that are highly heritable.
Conclusion
There is a need for a more effective and quick approach for the genetic modification of livestock, for the enhanced production, in order to fulfill the increasing demands of the growing population. A Nobel method in that field is genomic selection. It has an advantage over traditional breeding techniques since it may be used from an early age, shortening the generation gap and enhancing genetic gain. The full genetic data of livestock species may now be accessible extremely readily and inexpensively for breeding purposes because to advances in science and technology. Even if the application of genomic selection is less developed in other species than it is in dairy cattle, with the advancement of research, genomic selection will become an effective technique for the creation of elite animals in all livestock species. It is undoubtedly advantageous to conduct more research on how genomic selection operates in breeding programs, both for the benefit of the general population and for private breeding businesses.
References
-
FAO (2012) World Agriculture: towards 2030/2050 Interim Report. Food and Agriculture Organization of the United Nations, Rome, Italy.
-
Ibtisham F, Zhang L, Xiao M, An L, Ramzan MB, et al. (2017) Genomic selection and its application in animal breeding. Thai J Vet Med 47(3): 301-310.
-
Ventura RV, Miller SP, Dodds KG, Auvray B, Lee M, et al. (2016) Assessing accuracy of imputation using different SNP panel densities in a multi-breed sheep population. Genetics Selection Evolution 48: 1-20.
-
Garner JB, Douglas ML, Williams SRO, Wales WJ, Marett LC, et al. (2016) Genomic selection improves heat tolerance in dairy cattle. Sci Rep 6(1): 34114.
-
Boichard D, Ducrocq V, Croiseau P, Fritz S (2016) Genomic selection in domestic animals: principles, applications and perspectives. C R Biol 339(7-8): 274-277.
-
Deng T, Pang C, Ma X, Lu X, Duan A, et al. (2016) Four novel polymorphisms of buffalo INSIG2 gene are associated with milk production traits in Chinese buffaloes. Mol Cell Probes 30(5): 294-299.
-
Koning DJD (2016) Meuwissen et al. On Genomic Selection. Genetics 203(1): 5-7.
-
Azizian S, Shadparvar AA, Ghavi HZN, Shekalgorabi SJ (2016) Effect of increasing accuracy of genomic evaluations on economic efficiency of dairy cattle breeding programmes. Italian Journal of Animal Science 15(3): 379-385.
-
Zhou L, Ding X, Zhang Q, Wang Y, Lund MS, et al. (2013) Consistency of linkage disequilibrium between Chinese and Nordic Holsteins and genomic prediction for Chinese Holsteins using a joint reference population. Genet Sel Evol 45(1): 7.
-
König S, Simianer H, Willam A (2009) Economic evaluation of genomic breeding programs. J Dairy Sci 92(1): 382-391.
-
Shumbusho F, Raoul J, Astruc JM, Palhiere I, Lemarié S, et al. (2016) Economic evaluation of genomic selection in small ruminants: a sheep meat breeding program. Animal 10(6): 1033-1041.
-
Plieschke L, Edel C, Pimentel EC, Emmerling R, Bennewitz J, et al. (2016) Systematic genotyping of groups of cows to improve genomic estimated breeding values of selection candidates. Genet Sel Evol 48(1): 73.
-
Hickey JM, Bruce C, Whitelaw A, Gorjanc G (2016) Promotion of alleles by genome editing in livestock breeding programmes. J Anim Breed Genet 133(2): 83- 84.
-
Burrow HM, Mrode R, Mwai AO, Coffey MP, Hayes BJ, et al. (2021) Challenges and opportunities in applying genomic selection to ruminants owned by smallholder farmers. Agriculture 11(11): 1172.
-
Cowling WA, Stefanova KT, Beeck CP, Nelson MN, Hargreaves BL, et al. (2015) Using the animal model to accelerate response to selection in a self-pollinating crop. G3 (Bethesda) 5(7): 1419-1428.
-
Capper JL, Cady RA, Bauman DE (2009) The environmental impact of dairy production: 1944 compared with 2007. J Anim Sci 87(6): 2160-2167.
-
Sax K (1923) The association of size differences with seed-coat pattern and pigmentation in Phaseolus vulgaris. Genetics 8(6): 552-560.
-
Ribaut JM, Ragot M (2007) Marker-assisted selection to improve drought adaptation in maize: the backcross approach, perspectives, limitations, and alternatives. J Exp Bot 58(2): 351-360.
-
Goddard ME (1996) The use of marker haplotypes in animal breeding schemes. Genetics Selection Evolution 28(2): 161-176.
-
Ron M, Weller JI (2007) From QTL to QTN identification in livestock–winning by points rather than knock‐out: a review. Anim Genet 38(5): 429-439.
-
VanRaden PM, Van Tassell CP, Wiggans GR, Sonstegard TS, Schnabel RD, et al. (2009) Invited review: Reliability of genomic predictions for North American Holstein bulls. J Dairy Sci 92(1): 16-24.
-
Baruch E, Weller JI (2009) Incorporation of genotype effects into animal model evaluations when only a small fraction of the population has been genotyped. Animal 3(1): 16-23.
-
Fan B, Du ZQ, Gorbach DM, Rothschild MF (2010) Development and application of high-density SNP arrays in genomic studies of domestic animals. Asian- Australasian Journal of Animal Sciences 23(7): 833-847.
-
Recio OG, Gianola D, Rosa GJ, Weigel KA, Kranis A, et al. (2009) Genome-assisted prediction of a quantitative trait measured in parents and progeny: application to food conversion rate in chickens. Genetics Selection Evolution 41: 1-3.
-
Neves HH, Carvalheiro R, O’Brien AM, Utsunomiya YT, Carmo ASD, et al. (2014) Accuracy of genomic predictions in Bos indicus (Nellore) cattle. Genet Sel Evol 46(1): 17.
-
Wiggans GR, Vanraden PM, Cooper TA (2011) The genomic evaluation system in the United States: Past, present, future. J Dairy Sci 94(6): 3202-3211.
-
Jonas E, Koning DJD (2015) Genomic selection needs to be carefully assessed to meet specific requirements in livestock breeding programs. Front Genet 6: 49.
-
Lund MS, Roos AP, Vries AG, Druet T, Ducrocq V, et al. (2011) A common reference population from four European Holstein populations increases reliability of genomic predictions. Genet Sel Evol 43(1): 43.
-
Akanno EC, Plastow G, Woodward BW, Bauck S, Okut H, et al. (2014) Reliability of molecular breeding values for Warner-Bratzler shear force and carcass traits of beef cattle–An independent validation study. J Anim Sci 92(7): 2896-2904.
-
Mucha S, Mrode R, Lee IM, Coffey M, Conington J, et al. (2015) Estimation of genomic breeding values for milk yield in UK dairy goats. Journal of dairy science 98(11): 8201-8208.
-
Baloche G, Legarra A, Sallé G, Larroque H, Astruc JM, et al. (2014) Assessment of accuracy of genomic prediction for French Lacaune dairy sheep. J Dairy Sci 97(2): 1107- 1116.
-
Knol EF, Nielsen B, Knap PW (2016) Genomic selection in commercial pig breeding. Animal Frontiers 6(1): 15- 22.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania