A Study on the Lateral Channel of Scytalidium Catalase
Catalases are antioxidant enzymes. Their main function is to convert hydrogen peroxide, one of the reactive oxygen species, into water and oxygen. A bifunctional catalase enzyme from Scytalidium thermophilum, which belongs to the monofunctional catalase family, exhibits oxidase activity in the absence of hydrogen peroxide without the need for a cofactor. It is able to oxidize a variety of ortho- and para-diphenolic compounds, but the oxidase activity is very low (~ 10,000-fold) compared to its main activity in peroxide degradation. In this study, an attempt was made to improve the oxidase activity of catalase for its industrial utilization. For this purpose, the residues located in the lateral channel (H214, D224, and K248), which are thought to play an important role in the access of oxidase substances to the active site, were selected and converted into an amino acid with a smaller side chain -Ala- by site-directed mutagenesis. The changes in the catalytic efficiency of the variants were evaluated by spectrophotometric analysis. The results showed that oxidase activity was increased two- to threefold in all variants, while some exhibited decreased catalase activity. The D224A variant gave the best results as it showed approximately 2-fold increase in oxidase activity without loss of catalase activity (about 92% of the wild-type catalase activity was retained). Thus, a protein variant with improved properties was successfully produced.
Introduction
Thermophilic fungi can grow at temperatures between 20-60°C. As they are heat-resistant, they are preferred as thermostable enzyme sources in various industrial applications. Because of this property, thermophilic fungi are being studied by many researchers [1]. Catalases (EC 1.11.1.1.6) are metalloenzymes with a generally homotetrameric structure. Their main task is to catalyze the conversion of the hydrogen peroxide molecule (H2O2) into water and molecular oxygen. At low hydrogen peroxide levels, the enzyme can also exhibit peroxidase activity if a suitable electron donor is present in the environment [2]. In a study with mammalian catalase, the enzyme was reported to exhibit oxidase activity in the absence of H2O2, and the enzyme was named catalase oxidase [3]. This suggests that catalase-oxidase type enzymes with dual activity are much more common, including in humans.
Scytalidium thermophilum is a thermophilic fungus with an optimum temperature of 45-50°C, which considerably promotes the growth of Agaricus bisporus [4].
After sporulation, S. thermophilum turns dark brown and sometimes even charcoal black due to melanin and produces an extracellular catalase enzyme whose production is triggered in the presence of certain phenolic substances such as gallic acid and tannic acid, which occur in parallel with growth [5]. This enzyme, which has 77% homology with Penicillium vitale catalase, belongs to the group of ‘monofunctional large subunit catalase” [6]. Although it belongs to the family of monofunctional catalases, in the absence of H2O2 it can oxidize catechin, caffeic acid, catechol, chlorogenic acid and phenolic substances containing two hydroxyl groups in ortho-position, such as L-DOPA and para-diphenols like hydroquinone [7]. For this reason, the enzyme catalase-phenol oxidase (CATPO: Catalase-Phenol Oxidase) was named. The enzymatic activity, which is independent of hydrogen peroxide, requires the presence of oxygen and is inhibited by classical catalase inhibitors such as 3-amino-1,2,4-triazole (3TR) [8], albeit at a very low (~1 x 104-fold) level compared to catalase activity [2]. In view of this information, the CATPO enzyme of S. thermophilum can be catalytically improved by increasing the phenol oxidase activity of the CATPO enzyme of S. thermophilum by site- directed mutagenesis. The production of an efficient enzyme should contribute to the literature and lead to a better understanding of the dual activity and thus to new projects and the use of the enzyme in various industrial fields such as the textile industry, the food industry, and the development of biosensors.
Materials and Methods
Plasmids, Microorganisms, and Culture Preparation
A recombinant plasmid containing an N-terminal 6xHistag sequence adjacent to the catpo gene region, designated pET28a-CATPO [2] was used as the source of the catpo gene. Two E. coli bacterial strains, XL1-Blue and BL21 star (DE3), were used to produce different CATPO variants. During the cloning steps, the E. coli cells were grown aerobically at 37°C in LB medium supplemented with 50 µg/ mL kanamycin.
Site-directed Mutagenesis
Single point mutations were introduced into the coding region of catpo by QuikChange mutagenesis with Hot Start KOD DNA polymerase (Sigma), for 30 cycles prior to _Dpn_I digestion. PCR primers containing the desired mutations were purchased from BO&GA, Turkey and are listed in Table 1. Mutations were confirmed by DNA sequencing (Macrogen, Korea).
| Mutant | Sequence Change | Oligonucleotide* |
|---|---|---|
| H214A | CAC→GCC | GGGCTATGTCTGGTGCCGGTATCCCGCG |
| D224A | GAC→GCC | CGCGTTCTTACAGGCACATGGCCGGCTTCGG |
| K248A | AAA→GCC | GGCACTTCGCCTCTCGTCAGGGTAAAGCTAGTCTG |
Table 1: Mutations were confirmed by DNA sequencing (Macrogen, Korea).
Overexpression and Purification of CATPO Variants
After the pET28a-CATPO plasmid carrying the target mutation was transferred by transformation into strain E. coli BL21 star (DE3), a single colony carrying the mutation of interest was grown in LB medium (10 mL) containing 50 µg/ml kanamycin, then transferred to 1 liter of LB medium containing the same amount of kanamycin and allowed to grow at 600 nm until its OD was between 0.6 and 0.8. When the desired range was reached, 0.1 mM IPTG (=isopropylthio- β-galactoside) was added to increase enzyme production and incubated at 120 rpm for 24 hours. Subsequently, the cells were precipitated by centrifugation (10 min, 6000 rpm) [2].
For the purification of recombinant wild-type CATPO and 3 different lateral channel variants, the cells were stored at -80°C and kept on ice for a certain period. 3 hours in 100 mL lysis buffer (50 mM Na-HEPES, 25% (w/v) sucrose, 1% (v/v) Triton X 100, 5 mM MgCl2.6H2O, 1 mg/mL lysozyme, 5 μL benzonase nuclease (5KU, Sigma), 1:4 ratio of EDTA-free protease inhibitor tablets, pH 8.0). After lysis, the cells were precipitated at 10000 rpm (+4 °C) and the supernatant was placed on a dialysis membrane overnight and dialyzed in binding buffer (0.5 M NaCl with 20 mM sodium phosphate, pH 7.4) and prepared for purification. Affinity chromatography was used for one-step purification of His-tagged recombinant proteins as previously described [2, 8, 9].
Sodium Dodecyl Sulfate-polyacrylamide Gel Electrophoresis (SDS-PAGE)
To check the purity of the purified variants, an SDS-PAGE was performed. Electrophoresis was performed with the Bio-Rad Mini-Protean system. A 5% stacking gel and a 15% separating gel were prepared [10].
Enzyme Assays
Catalase and phenol oxidase activities were determined as previously described [2, 8, 9]. One unit of catalase was defined as the amount of enzyme catalyzing the degradation of 1 µmol H2O2 per minute in a 10 mM H2O2 solution at pH 7.0 and 60°C. One unit of phenol oxidase is defined as the amount that catalyzes the formation of 1 nmol of product per minute at 60 °C. The determination of phenol oxidase activity was performed using catechol as substrate [11].
Protein Concentration
Protein concentration analysis was determined by the Bradford method using bovine serum albumin (BSA) as the protein standard to generate the standard protein curve [12].
Results and Discussion
Site-Directed Mutagenesis on catpo Gene
To increase phenol oxidase activity of the CATPO enzyme without affecting catalase activity, site-directed mutagenesis was used to make mutations in the amino acids H214, D224, and K248, which are in the lateral channel of the enzyme and are presumably important for phenol oxidase activity. Different temperatures between 55°C and 63°C were used to bind the mutagenic trimers used in the Quick Change mutagenesis technique to the target DNA. The DNA bands of WT-CATPO and the variants with a length of about 8 kbp were observed by agarose gel electrophoresis (Figure 1). The optimal binding temperature for mutations H214A and K248A was 55.9°C; the optimal binding temperature for mutation D224A was 55-60°C. The optimal binding temperatures used to generate different CATPO variants, including H82N, V123F, E316F, H246W, and V228A, by site- directed mutagenesis studies [2, 8, 9] were similar to the binding temperatures in this study.
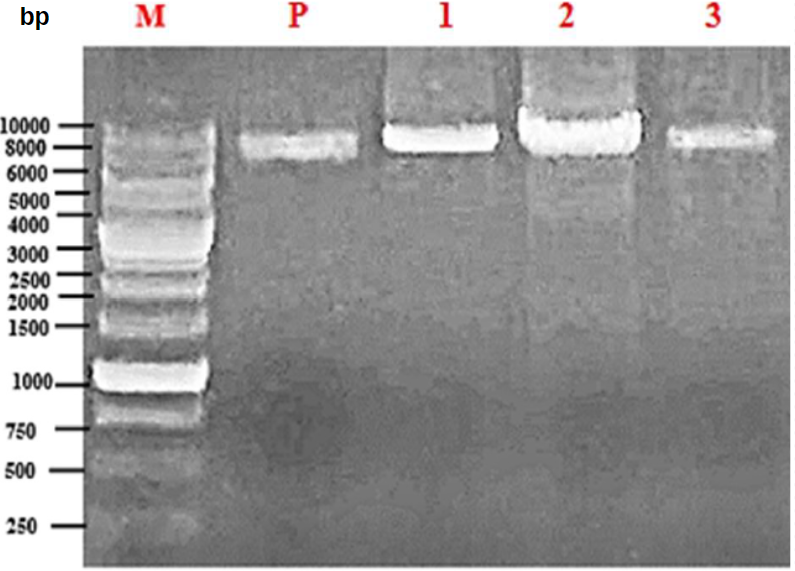
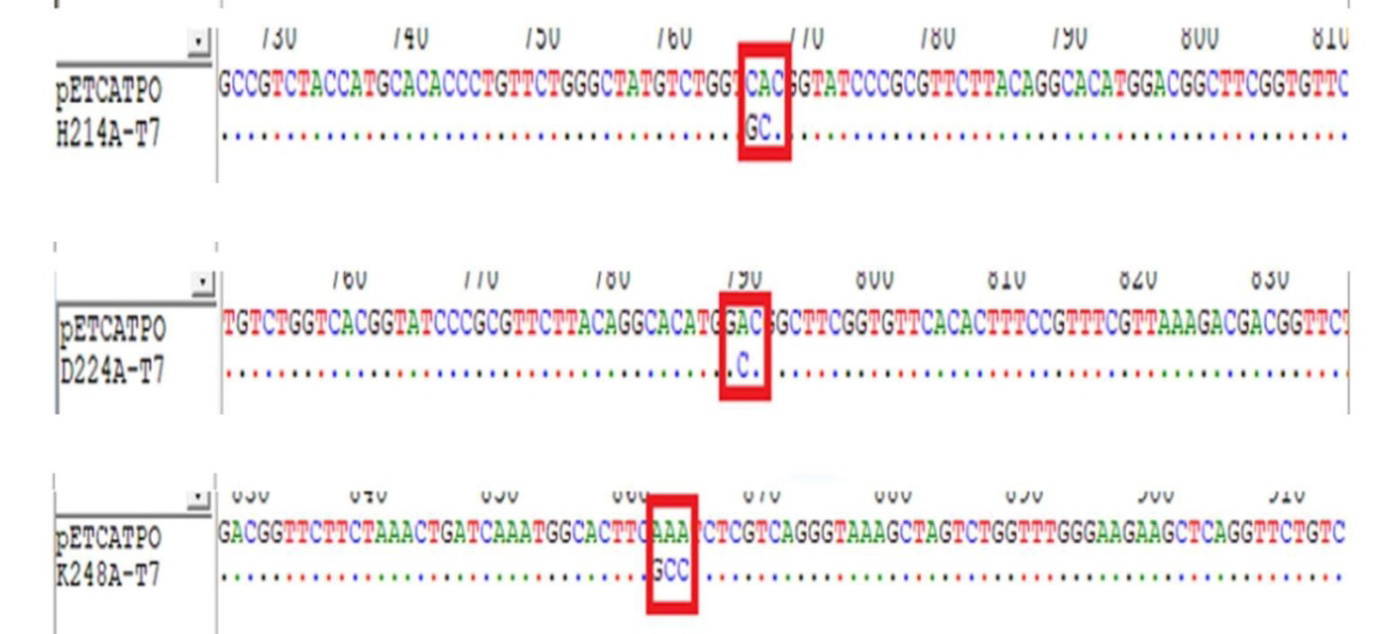
The DNA sequence of the WT-CATPO plasmid and the DNA sequences of the five mutant variants were compared using the Bioedit program, and the presence of the desired mutations was confirmed (Figure 2). It was found that the five variants whose DNA sequences were compared with pET28a-CATPO contained mutations with the desired trait in the designed gene regions and a clean sequence was obtained without any contamination in the sequence. The results of DNA sequence analysis of H214A, D224A and K248A, respectively, are shown in Figure 2.
Purification of CATPO Variants by Affinity Chromatography
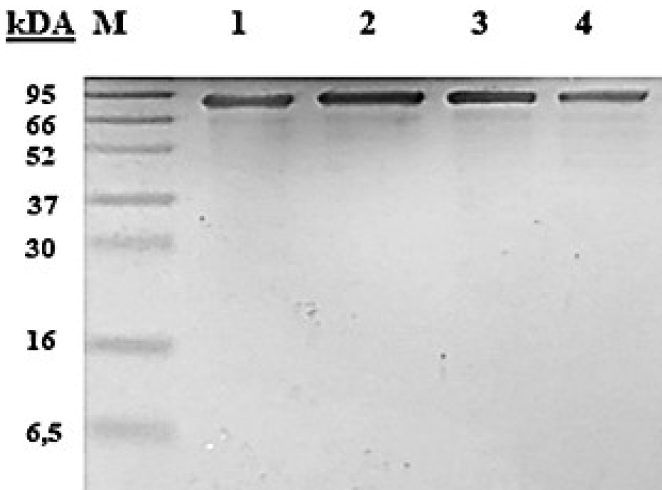
Purification of histidine-labelled wt-CATPO and its three variants was performed by affinity chromatography in one step as described in the Materials and Methods section. The purity of the proteins was checked by SDS-PAGE, and in all cases a single band of about 80 kDa was detected (Figure 3).
Comparison of Specific Activities of wt-CATPO and Its Variants
The CATPO mutants listed in Table 2 were generated by specifically converting the amino acids H214, D224 and K248, which are located at or near the substrate/inhibitor binding site defined for phenolic compounds at the lateral channel [8], to the small-chain amino acid Ala. The aim was to expand the lateral channel to make it more suitable for phenolic compounds and consequently increase oxidase activity.
| Specific Activity | %WT Activity | |||
|---|---|---|---|---|
| Mutant | Catalase Activity (μmol/mg/min) | Phenol Oxidase Activity (nmol/mg/min) | Catalase | Phenol Oxidase |
| wt- CATPO | 15700 ± 281 | 211 ± 2 | 100 | 100 |
| H214A | 2340 ± 3 | 360 ± 1 | 15 | 171 |
| K248A | 5028 ± 27 | 524± 1 | 32 | 248 |
| D224A | 14500 ± 175 | 351 ± 1 | 92 | 166 |
Table 2: Comparison of specific activities of wt-CATPO and its variants.
As expected, phenol oxidase activity was significantly increased in all mutant proteins compared to wt-CATPO (Table 2). However, the catalase activities of H214A and K248A were reduced by 85 % and 68 %, respectively. H214, which carries a positively charged imidazole ring, is in the lateral channel upstream of the inhibitor binding site, approximately 7.8 angstroms away from the catalase inhibitor 3TR identified in previous studies [8]. It forms a polar bond with N156, which is partially located near the active site. The 85% loss of catalase activity observed in the A214 variant created by the mutation suggests that the presence of the amino acid histidine at this position is important. Considering that the amino acid histidine is involved in the stabilization of protein structure [13], its conversion to a hydrophobic amino acid with a smaller side chain (Ala) may have affected the protein structure, which would explain the decrease in catalase activity.
The positively charged amino acid K248 is located on the surface of the substrate/inhibitor binding site [8] in the lateral channel. It interacts ionically with D224 in this region. It also tends to form H-bonds with the water molecule inside the protein together with H246 and E316. In other words, K248 constricts the lateral channel due to its long side chain but interacts with the polar amino acids in its vicinity. Therefore, the conversion of K248 to A248 may have accelerated the passage of phenols here by widening the lateral channel but may also have led to a decrease in catalytic activity because it prevents the formation of weak bonds that stabilize the tertiary structure of the protein.
Consistent with these results, the phenol oxidase activities of all mutants were observed to increase compared to the wt-CATPO enzyme, presumably due to the increase in lateral channel diameter because of the mutations. The results of the specific activity analysis show that the phenol oxidase activities of all variants were increased 2- to 3-fold compared to the wt-CATPO enzyme. However, only the D224A variant showed an increase in phenol oxidase activity (2-fold increase) without a decrease in catalase activity compared to wt-CATPO.
A similar study with the lateral channel [8] showed the presence of a substrate/inhibitor binding site for various phenolic compounds in this channel. As a result of the mutations performed in this study, the increase in phenol oxidase activity of all mutants compared to the wt-CATPO enzyme supports the presence of a substrate binding site for various phenolic compounds.
Conclusion
In this study, site-directed mutation studies were performed to increase the oxidase activity of the CATPO enzyme. For this purpose, different amino acids (H214, D224, and K248) were targeted in the lateral channel and the desired mutations were generated by site-directed mutagenesis. When comparing the catalase and phenol oxidase activities of wt-CATPO and the three variants, a 2-3- fold increase in the phenol oxidase activities of the variants was observed. Compared to wt-CATPO, the phenol oxidase activities of mutants H214A (171%), D224A (166%), and K248A (248%) were higher. On the other hand, a decrease in catalase activity was observed with all variants except D224A. Thus, D224A proved to be the variant that fulfilled the desired conditions for the development of an effective enzyme and was more active in terms of phenol oxidase activity compared to wt-CATPO without decreasing catalase activity. It is expected that this study will be groundbreaking for future enzyme and protein engineering studies.
Acknowledgement
This work was funded by Kocaeli University (Grant No. FYL-2019-1568 and FMP-2023-3425).
References
-
Ögel ZB, Yarangümeli K, Dü H, Ifrij I (2001) Submerged cultivation of Scytalidium thermophilum on complex lignocellulosic biomass for endoglucanase production. Enzyme Microb Technol 28(7-8): 689-695.
-
Yuzugullu Y, Trinh CH, Fairhurst L, Ogel ZB, Mcpherson MJ, et al. (2013) Investigating the active centre of the _Scytalidium thermophilum_ catalase. Acta Crystallogr Sect F Struct Biol Cryst Commun 69(pt 4): 369-375.
-
Vetrano AM, Heck DE, Mariano TM, Mishin V, Laskin DL, et al. (2005) Characterization of the oxidase activity in mammalian catalase. J Biol Chem 280(42): 35372- 35381.
-
Wiegant WM (1992) Growth characteristics of the thermophilic fungus _Scytalidium thermophilum_ in relation to production of mushroom compost. Appl Environ Microbiol 58(4): 1301-1307.
-
Yuzugullu Y, Ögel ZB, Bölükbaşi U, Coruh N, Karakaş G (2011) Production of a novel bifunctional catalase- phenol oxidase of _Scytalidium thermophilum_ in the presence of phenolic compounds. Turk J Biol 35(6): 697- 704.
-
Kocabas DS, Bakir U, Phillips SEV, Mcpherson MJ, Ogel ZB (2008) Purification, characterization, and identification of a novel bifunctional catalase phenol oxidase from _Scytalidium thermophilum_. Appl Microbiol Biotechnol 79: 407-415.
-
Avci GK, Coruh N, Bolukbasi U, Ogel ZB (2013) Oxidation of phenolic compounds by the bifunctional catalase- phenol oxidase (CATPO) from _Scytalidium thermophilum_. Appl Microbiol Biotechnol 97(2): 661-672.
-
Karakus YY, Goc G, Balci S, Yorke, BA, Trinh CH, et al. (2018) Identification of the site of oxidase substrate binding in _Scytalidium thermophilum_ catalase. Acta Crystallogr D Struct Biol 74(10): 979-985.
-
Yuzugullu Y, Trinh CH, Smith MA, Pearson AR, Phillips SEV, et al. (2013) Structure, recombinant expression and mutagenesis studies of the catalase with oxidase activity from _Scytalidium thermophilum_. Acta Crystallogr D Biol Crystallogr 69(3): 398-408.
-
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227(5259): 680-685.
-
Ögel ZB, Yüzügüllü Y, Mete S, Bakir U, Kaptan Y, et al. (2006) Production, properties and application to biocatalysis of a novel extracellular alkaline phenol oxidase from the thermophilic fungus _Scytalidium thermophilum_. Appl Microbiol Biotechnol 71(6): 853-862.
-
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. A n a l Biochem 72: 248-254.
-
Chelikani P, Fita I, Loewen PC (2004) Diversity of structures and properties among catalases. Cell Mol Life Sci 61(2): 192-208.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania