Development of Techniques about Production of Cells and Nucleotide Sequences with Immunogenic Properties
In vitro-cultures of normal mouse embryonic fibroblasts from 3T3 cell line, but also of mouse malignant myeloma cells, as well as mixed cultures of both cellular types were prepared and proceeded in different conditions. In the mouse malignant myeloma cells, containing inserted Murine Leukemia Virus (MuLV) (with RNA-genome, Retroviridae family), signs of increased malignant in vitro-potential were noted in comparison to non-containing mouse myeloma cells as decreased sizes and number of the myeloma cells, but also their decreased capability for 3D-growth. In pre-incubation of normal mouse embryonic fibroblasts in cultural fluid, supplemented by previous cultivation of mouse malignant myeloma cells in it, followed by freezing in the presence of cryo-protector Dimethylsulfoxide (DMSO), subsequent thawing and re-incubation, appearance of hybrid cells of myeloid-like and phagocyte-like cells, as well as lymphoid-like and plasmatic cells with malignant myeloma cells, were noted. In co-cultivation of normal mouse embryonic fibroblasts with mouse malignant myeloma cells, containing inserted MuLV, followed by freezing in the presence of DMSO, subsequent thawing and re-incubation, exchange of nucleotide sequences between viral particles and normal mouse cells was proposed on the influence of the used organic detergent, as well as of the drastic temperature changes. Possibilities about production of cellular and recombinant viral vaccines by exchange of nucleotide sequences between viral particles and cells could be suggested.
Introduction
The techniques for in vitro-_cultivation of viruses in cell cultures allow the design of various viral recombinants, including of recombinant viral vaccines. As appropriate tools can be used for DNA- and RNA-viruses [1, 2, 3, 4, 5], but also bacteria plasmids and yeasts [4, 6]. For this aim, an intact _tk gene, coding the enzyme timidinkinase (TK), has been found to be necessary, but, on the other hand – the integration of the searched gene(s) out of tk locus of the viral genome, as well as virus promoter, which could provide the expression of the inserted gene(s). Changes in the promoter and/or in the insertion site, as well as in the target vector repeats in fragments, expressing proteins with immunomodulator functions, have been proven to be possible. In this way have been inserted genes, coding cell receptors, cytokines, enzymes, complement activators, apoptosis activators and/ or inhibitors, surface antigens, tumor markers. Besides the respective inserted gene(s), a marker gene has also been found to be necessary, but both gene types are controlled by appropriate promoter sequences. Most often, as marker genes could be used such, which code color proteins (for example, fluorescent protein, as GFP, YFP, RFP) or products, participating in a color reaction (enzymes luciferase or timidinkinase), or is connected in any way resistance to any antibiotic preparation [3]. The described gene sequences should be connected by treatment with specific ligases, but the used initial vector constructs should contain specific restriction sites, which is necessary the withed DNA-fragments of interest be obtained by treatment with respective restriction enzymes (most often bacterial endo- nucleases). Other important components are the reverted end DNA-repeats.In this connection, the main goal of the current study is directed to development of in vitro-methods of protection and immunogenicity.
Materials and Methods
Normal mouse embryonic fibroblasts from line 3T3 were incubated Dulbecco’s Modified Minimal Essential Medium (DMEM) (Sigma-Aldrich), supplemented with 10% Fetal Calf Serum (FCS) (Sigma-Aldrich), 100 U/ml penicillin (Sigma-Aldrich) and 100 μg/ml streptomycin (Sigma- Aldrich). Also, suspension cultures of mouse malignant myeloma cells, containing inserted Murine Leukemia Virus (MuLV) (Retroviridae family) were incubated in analogically supplemented medium RPMI 1640. Mixed cultures were also prepared and analogically proceeded, by pre-incubation of sub-populations of normal mouse embryonic fibroblasts in cultural fluid, supplemented by previous incubation of mouse malignant myeloma cells in it, subsequently centrifuged and filtered, but also by addition of cultural fluids plus suspensions of cells from both types. All cultures were cultivated at 370C in incubator with 5% СО2 and 95% air humidification. All cells were observed by inverted light microscope (Leica). Sub-population of 3T3 mouse embryonic fibroblasts from line were incubated in supplemented cultural fluids after previous co-cultivation with mouse malignant myeloma cells, and another sub-group was co- cultivated with sub-population of mouse malignant myeloma cells (cultural fluids plus suspensions of both cellular types). Sub-populations of both types of mixed cultures, described above, were then freezed in the presence of cryo-protector Dimethylsulfoxide (DMSO), subsequently thawed and re- cultivated in fresh medium in the standard conditions, described above.
Results
In the mouse malignant myeloma cells, containing inserted MuLV, were noted signs of increased malignant in vitro-potential (Figure 1A), compared to non-containing mouse myeloma cells (Figure 1B). As signs were accepted the observed decreased sizes and number of the myeloma cells, but also their decreased capability for 3D-growth (Figure 1B). In pre-incubation of normal mouse embryonic fibroblasts in cultural fluid, supplemented by previous cultivation of mouse malignant myeloma cells in it, subsequent freezing in the presence of cryo-protector DMSO, followed by thawing and re-incubation, appearance of hybrid cells of myeloid- like and phagocyte-like cells, as well as lymphoid-like and plasmatic cells with malignant myeloma cells, were noted (Figure 1C). In co-cultivation of normal mouse embryonic fibroblasts with mouse malignant myeloma cells, containing inserted MuLV, subsequent freezing in the presence of DMSO, followed by thawing and re-incubation, exchange of nucleotide sequences between viral particles and normal mouse cells was proposed on the influence of the used organic detergent, as well as of the drastic temperature changes (Figure 1D). On this principle, possibilities about production of cellular and recombinant viral vaccines by exchange of nucleotide sequences between viral particles and cells could be suggested.
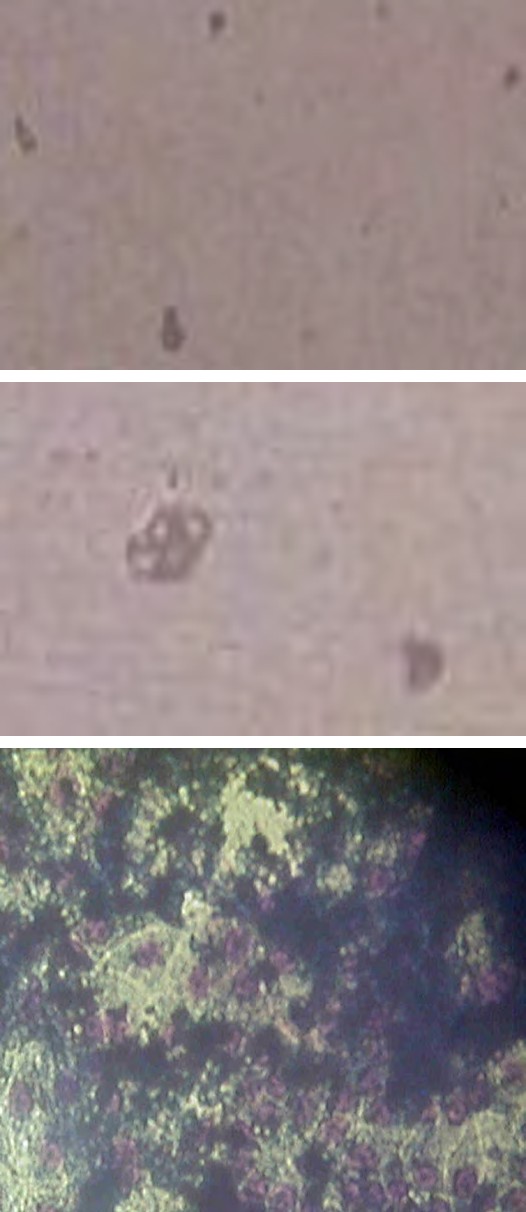
Figure 1A: mouse malignant myeloma cells, non- containing inserted viral genome sequences.
Figure 1B: mouse malignant myeloma cells, transfected previously with MuLV.
Figure 1C: incubated in cultural fluid from mouse malignant myeloma cells (eventually containing nucleotide sequences from the transforming vector), freezed in the presence of DMSO, thawed and re-incubated. Signs of initial myeloid and phagocyte, as well as of lymphoid and plasmatic cells phenotype, respectively, could be noted.
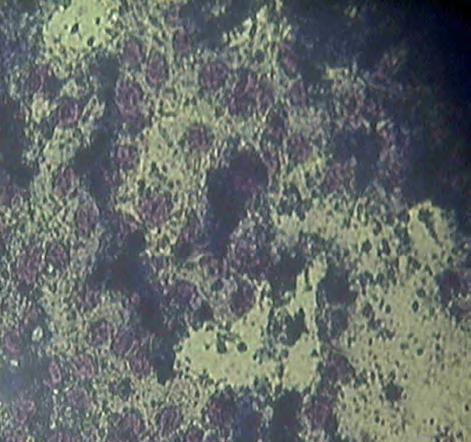
Figure 1D: co-cultivated with mouse malignant myeloma cells, freezed in the presence of DMSO, thawed and re- incubated. Appearance of hybrid cells between myeloid- like progenitors and myeloma cells and between lymphoid- like progenitors and malignant myeloma cells, respectively, could be seen, Fixed light-microscopy preparations, stained by Giemsa-dye, magnification: 100X.
Discussion
The results obtained could be explained with activated fusion between separate cells on the influence of DMSO, as well as of the drastic temperature changes. They were in confirmation of literature data about the activated fusion on the influence of DMSO. The observed changes could be explained with eventual existence of sub-populations of stem-like cells in the general normal embryonic 3T3 cell line, which are able to differentiate in various directions, depending of the respective incubation conditions. Messages about similarly derived hybrid cells have been received in fusion of malignant cells with myeloid and lymphoid cells [7], but also with normal fibroblasts [8]. The current data also suggested a possibility for appearance of membrane glycoprotein receptors on the surface cells in initial phases of differentiation in appropriate conditions as for instance viral and/or malignant antigens, as well as different types of appropriate immunomodulators. These results were in agreement with the literature findings in this direction [9, 10, 11, 12, 13]. On the other hand, both myeloid and lymphoid cells have shown capabilities for differentiation in respective directions in transfer in them of appropriate nucleotides, coding respective antigens, on one hand, but also as usable novel safe vaccines, carrying genes for respective malignant antigens or cytokines, on the other. Furthermore, the existence of “intrinsic antiviral immunity” changes, as internal protection of the cell in response to its infection has been proved [14, 15]. Another explanation could be connected with the proved action of some cell-produced proteins as glycoprotein receptors to specific bio-molecules [16]. In this way, the current data supported the importance of various intra- and extra-cellular inter-molecular interactions in cascade regulatory pathways, underlining functions of the different types of cells [17, 18].
References
-
Barrette S, Douglas JL, Seidel NE, Bodine DM (2000) Lentivirus-based vectors transduce mouse hematopoietic stem cells with similar efficiency to moloney murine leukemia virus-based vectors. Blood 96(10): 3385-3391
-
Borysiewicz LK, Flander A, Numako M, Man S, Wilkinson GW, et al. (1996) A recombinant vaccinia virus encoding human papillomavirus types 16 and 18, E6 and E7 proteins as immunotherapy for cervical cancer. Lancet 347(9014):1523-1527.
-
Boyle D, Coupar B, Gibbs A, Seigman L, Both G (1987) Fowlpox virus thymidine kinase: nucleotide sequence and relationships to other thymidine kinases. Virology 156(2): 355-365
-
Brachmann CB, Davies A, Cost GJ, Caputo E, Li J, et al. (1998) Designer deletion strains derived from _Saccharomyces_ _cerevisiae S288C_: a useful set of strains and plasmids for PCR mediated gene disruption and other applications. Yeast 14(2): 115-132.
-
Chen S, Agarwal A, Glushakova OY, Jorgensen MS, Salgar SK, et al. (2003) Gene delivery in renal tubular epithelial cells using recombinant adeno-associated viral vectors. J Am Soc Nephrol 14(4): 947-958.
-
Domi A, Moss B (2005) Engineering of a _vaccinia virus_ bacterial artificial chromosome in _Escherichia coli_ by bacteriophage λ−based recombination. Nat Meth 2(2): 95-97.
-
Freiberg S, Klein G, Wiener F, Harris H (1975) Hybrid cells derived from fusion of Ta2-Ha ascites carcinoma with normal fibroblasts. II. Characterization of isoantigenic variants sublines. J Natl Cancer Inst 50(5): 1269-1286.
-
Gong J, Nikrui N, Chen D, Koido S, Wu Z, et al. (2000) Fusions of human ovarian carcinoma cells with autologous or allogeneic dendritic cells induce antitumor immunity. J Immunol 165(3): 1705-1711.
-
Keller GM (2005) Embryonic stem cell differentiation: emergence of a new era in biology and medicine. Genes & Dev 19(10): 1129-1155.
-
Kyba M, Perlingeiro RCR, Hoover RR, Lu C-W, Pierce J, et al. (2003) Enhanced hematopoietic differentiation of embryonic stem cells conditionally expressing Stat5. Proc Natl Acad Sci USA 100(1): 11904-11910.
-
McDonnald PP, Bald A, Cassatella MA (1997) Activation of NF-kappaB pathway by inflammatory stimuli in human. Blood 89(9): 3421-3433.
-
Norwood TH, Zeigler CJ, Martin GM (1976) Dimethyl sulfoxide enhances polyethylene glycol-mediated cell fusion. Somat Cell Genet 2(3): 263-270.
-
Okubo T, Yanai N, Watanabe S, Arai K-I, Obinata M (2000) Effect of human granulocyte-macrophage colony stimulating factor (hGM-CSF) on lymphoid and myeloid differentiation of sorted hematopoietic stem cells from hGM-CSF receptor gene transgenic mice. J Biochem 127(4): 591-596.
-
Cheng L, Du C, Lavau C, Chen S, Tong J, et al. (1998) Sustained gene expression in _retrovirally_ transduced, engrafting human hematopoietic stem cells and their lympho-myeloid progeny. Blood 92(1): 83-92.
-
Murray MJ, Peters NE, Reeves MB (2018) Navigating the host cell response during entry into sites of latent cytomegalovirus infection. Pathogens 7(1): 30.
-
Yan N, Chen ZJ (2012) Intrinsic antiviral immunity. Nat Immunol 13(3): 214-222.
-
Popi AF, Motta FLT, Mortara RA, Schenkman S, Lopes JD, et al. (2009) Co-ordinated expression of lymphoid and myeloid specific transcription factors during B-1b cell differentiation into mononuclear phagocytes _in vitro_. Immunology 126(1): 114-122.
-
Schöler HR, Hatzapoulis AK, Balling R, Suzuki N, Gruss P (1989) A family of octamer-specific proteins present during mouse embryogenesis: evidence for germline- specific expression of an Oct factor. EMBO J 8(9): 2543- 2550.
- Update on Malariology and Malaria Vaccines
- Addressing Vaccine Hesitancy in the Age of Measles Resurgence: A Mini-Review
- Exploring Barriers and Facilitators of Group Antenatal Care Implementation in Kaduna State, Nigeria: A Qualitative Evaluation
- The Role of IL-11 in Regenerative Medicine and Tissue Engineering
- New Prediction of Mortality rate of Covid -19 According to WHO Estimation
- Measles Vaccine in Kano, Northern Nigeria: Past, Present and Future