Hashimoto’s Thyroiditis Autoimmune Disease: Background and Current Status, Update Overview of Biotechnological and Biomedical Fields and Future Trends for 3D Models
Hashimoto thyroiditis, also known as chronic autoimmune thyroiditis or chronic lymphocytic thyroiditis, is an autoimmune illness in which thyroid cells are damaged by immunological mechanisms involving cells and antibodies. Thyroid peroxidase and/or thyroglobulin autoantibodies in the serum are biochemical indicators of the condition, with females having a higher incidence than males and increasing with age. It’s the leading cause of hypothyroidism in affluent countries. Inadequate dietary iodine intake, on the other hand, is the most common cause of hypothyroidism worldwide. The development of antithyroid antibodies that target the thyroid tissue, causing gradual fibrosis, is the pathogenesis of Hashimoto thyroiditis. The diagnosis can be difficult, and as a result, the problem is frequently not detected until late in the disease process. The most prevalent laboratory findings are raised TSH and low thyroxine (T4) levels, as well as enhanced antithyroid peroxidase (anti-TPO) antibodies. The pathogenesis, diagnosis, and management of Hashimoto thyroiditis are discussed in this article.
Introduction
Autoimmune thyroid diseases (AITDs) are the most frequent autoimmune diseases, affecting roughly 5% of the general population and predominating in women. AITDs are the most common cause of acquired thyroid disease during infancy and adolescence, despite reaching a peak in maturity. AITDs include Hashimoto’s thyroiditis (HT) and Graves’ disease (GD), both of which are characterized by thyroid infiltration by T and B cells that respond against thyroid antigens and the production of thyroid autoantibodies [1]. Hashimoto thyroiditis is an autoimmune illness that damages thyroid cells through immunological mechanisms mediated by cells and antibodies. It’s the leading cause of hypothyroidism in affluent countries. In contrast, the most prevalent cause of hypothyroidism worldwide is a lack of iodine in the diet. This condition is also known as chronic lymphocytic thyroiditis and chronic autoimmune thyroiditis [2]. The pathogenesis of the disease is characterized by the production of antithyroid antibodies, which target thyroid tissue and cause progressive fibrosis. Diagnosis is frequently difficult and may not occur until late in the disease phase. The most prevalent laboratory results include elevated thyroid- stimulating hormone (TSH) levels and low levels of free thyroxine (fT4), as well as enhanced antithyroid peroxidase (TPO) antibodies [3]. Patients may show signs, symptoms, and laboratory findings of hyperthyroidism or normal levels earlier in the course of the disease. This is due to the fact that thyroid gland cell death can be intermittent. Women are disproportionately affected. Women outnumber men by a factor of at least ten. Although some sources suggest that diagnosis occurs more frequently in the fifth decade of life, the majority of women are diagnosed between the ages of 30 and 50 [3]. Levothyroxine, at a recommended dose of 1.6 to 1.8 mcg/kg/day, is the standard treatment. In the human body, T4 is converted to T3, which is the active form of thyroid hormone. Excessive supplementing can have negative and morbid consequences, including arrhythmias (the most prevalent of which is atrial fibrillation) and osteoporosis. The pathogenesis, diagnosis, and management of Hashimoto thyroiditis are discussed in this article [2].
Background of Hashimoto’s Thyroiditis
Hashimoto’s thyroiditis is named after Japanese physician Hakaru Hashimoto of Kyushu University’s medical school, who was the first to describe the symptoms of people with Struma lymphomatosa, a severe lymphocyte infiltration of the thyroid gland. In addition, all of the data in his first study came from four women. These findings explained the clinical features seen in these women, including lymphoid and plasma cell infiltration, lymphoid follicle development with germinal centers, fibrosis, degraded thyroid epithelial cells, and leukocytes in the lumen [4]. Despite Dr. Hashimoto’s discovery and publishing, the disease was not recognized as separate from Reidel’s Thyroiditis, a widespread disease in Europe at the time. Despite the fact that many earlier investigations had been documented and published, Hashimoto’s strumalymphomatosa was only recognized as an early phase of Reidel’s Thyroiditis in the early 1900s. It wasn’t until 1931 that the disease was recognized as a separate entity, when Cleveland researchers Allen Graham characterized the symptoms and presentation in the same way as Hakuru did [5]. Drs. Rose and Witebsky were able to show in 1956 that immunizing specific mice with thyroid extracts from other rodents mimicked the condition Hakuru and other researchers were seeking to describe. Anti-thyroglobulin antibodies were also identified in blood serum samples from the same animals by these specialists Cappa, et al. [6]. Researchers from the Middlesex Hospital in London were able to conduct human studies on individuals with comparable symptoms later that year. They were able to determine that these unwell patients have an immunological reaction to human thyroglobulin after purifying anti- thyroglobulin antibodies from their serum. Based on these information, it was hypothesized that Hashimoto’s struma is an autoimmune thyroid disease [3].
Risk Factors
These factors may increase your chances of getting Hashimoto’s disease:
a) Sex: Hashimoto’s disease is significantly more common in women. b) Age: Hashimoto’s disease can strike at any age, but it is most frequent in middle life. c) Heredity: If you have a family history of thyroid or other autoimmune diseases, you’re more likely to get Hashimoto’s disease. d) Other autoimmune disease: You’re more likely to acquire Hashimoto’s disease if you have another autoimmune condition like rheumatoid arthritis, type 1 diabetes, or lupus. e) Radiation exposure: Hashimoto’s disease is more common in people who are exposed to high levels of environmental radiation.
Hashimoto’s Stage
Hashimoto’s thyroiditis is a type of autoimmune disease that affects the thyroid gland. This means that our immune system sees the thyroid gland as a foreign intruder and begins to assault it, resulting in thyroid tissue loss.
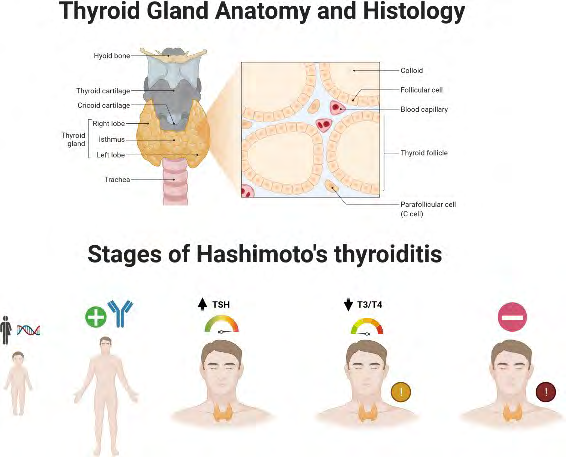
Figure 1: Thyroid Gland Anatomy and Histology and 5 Stages of Hashimoto autoimmune thyroid disease: 1) A person has a genetic predisposition to develop Hashimoto’s and thyroid function is normal, without attack on the thyroid. 2) A person may test positive for thyroid antibodies and may have changes consistent with Hashimoto’s but have normal TSH level. 3) Thyroid glands start to lose its ability to make enough hormone and there will be a slightly elevated TSH with normal T3/T4. More symptoms will be seen at this stage. 4) Thyroid glands have fully lost its ability to compensate, elevate TSH and low T3/T4 levels. With overt hypothyroidism, Hashimoto’s are often diagnosed. 5) Fully progression of the autoimmune response and other types of autoimmune conditions may develop. Created with BioRender.com
The person’s ability to create thyroid hormone will be lost if this degradation continues for a long time. Hashimoto’s disease is rarely diagnosed until it has progressed to the advanced stages, when the thyroid gland has been severely damaged. Wiersinga and colleagues identified five stages of Hashimoto’s disease in 2014 Yamada, et al. [7].
Clinical Criteria
Doctors may test for Hashimoto’s disease if you’re feeling increasingly tired or sluggish, have dry skin, constipation, or a hoarse voice, or if you’ve had past thyroid problems or goiter. Signs and symptoms, as well as the results of blood tests that measure thyroid hormone and thyroid- stimulating hormone (TSH) levels produced in the pituitary gland, are used to diagnose Hashimoto’s disease. A hormone test, done through a blood test, can reveal the amount of hormone produced by the thyroid and pituitary gland. Thyroid hormone levels are low when your thyroid is underactive. TSH is raised at the same time because your pituitary gland is attempting to stimulate your thyroid gland to create more thyroid hormone [1]. An antibody test is useful as the creation of aberrant antibodies is the cause of Hashimoto’s disease, which is an autoimmune condition. Antibodies against thyroid peroxidase (TPO antibodies), an enzyme ordinarily found in the thyroid gland and involved in the generation of thyroid hormones, can be detected using a blood test. However, not everyone with Hashimoto’s thyroiditis has a positive TPO antibody test. Many patients have TPO antibodies but no goiter, hypothyroidism, or other thyroid issues [1]. Previously, doctors couldn’t discover an underactive thyroid (hypothyroidism), the primary symptom of Hashimoto’s disease, until the disease had progressed. The sensitive TSH test allows doctors to discover thyroid issues early, and in certain cases, before you have any symptoms. Because the TSH test is the most accurate screening tool, your doctor would most likely order a TSH test first and then a thyroid hormone test if necessary. TSH testing are also important in the treatment of hypothyroidism. These tests also assist your doctor in determining the appropriate medicine dosage, both at the outset and over time [1].
Treatment and Management of Hashimoto’s Disease
Replacement thyroid hormone is the most common treatment for hypothyroidism. Orally administered titrated levothyroxine sodium is the medicine of choice. It has a 7-day half-life and can be used on a daily basis. To avoid poor absorption, it should not be taken with iron or calcium supplements, aluminum hydroxide, or proton pump inhibitors. For optimal absorption, take it first thing in the morning on an empty stomach [8]. The typical dose is 1.6-
1.8 mcg/kg per day, but this varies from patient to patient. Patients under the age of 50 should be started on the full dose; however, patients with cardiovascular disorders and the elderly should be given lesser doses. The recommended beginning dose for patients over 50 years old is 25 mcg/ day, with reevaluation after six to eight weeks. In contrast, the dose of thyroxine must be increased by 30% during pregnancy, and larger doses of levothyroxine are required to maintain euthyroid status in patients with short-bowel syndrome [8].
Genetic Susceptibility
Human Leukocyte Antigen (HLA) Genes
The major histocompatibility complex (MHC) region on chromosome 6p21, which encodes human leukocyte antigens (HLAs), was the first gene locus linked to autoimmune thyroid disease. Several immune response genes are found in the HLA area, which is highly polymorphic. The HLA molecule, which is found on antigen presenting cells (APCs), binds and delivers an antigenic peptide, allowing T cells to recognize and respond to it Chen, et al. [9]. HLA alleles with a higher affinity for auto antigenic thyroidal peptides are thought to play a role in the development of autoimmune thyroid disease. However, autoantigen occurrence within the thyroid or thyroid draining lymph nodes is required to induce thyroid autoimmunity, which is followed by HLA presentation. On thyrocytes, abnormal expression of HLA class II molecules has been seen in HT [10].
Cytotoxic T lymphocyte Antigen-4 (CTLA-4) Gene
On chromosome 2q33, the CTLA-4 gene, which is the second key immune-regulatory gene linked to autoimmune thyroid disease, is located. T-cell activation is suppressed by the production of CTLA-4 on the surface of T cells, which is produced by T-cell receptor activation. CTLA-4 gene polymorphisms may lower CTLA-4 antigen expression or function, resulting in less suppression of T-cell proliferation and, as a result, increased susceptibility to autoimmune responses [11]. Several variants of the CTLA-4 gene have been examined in HT patients in the past. Among them, the (AT)n microsatellite CTLA-4 polymorphism in the untranslated region (UTR) was linked to HT in Caucasian and Japanese patients, but not in Italians. A 49A/G single nucleotide polymorphism (SNP) in exon 1 that results in threonine to alanine substitution has been linked to HT, but other investigations have not verified this association. A major meta-analysis of 866 HT patients found a substantial link with 49A/G, using both published and unpublished data [11].
Thyroglobulin Gene
Thyroglobulin is a significant thyroid-specific antigen that is also found in the bloodstream, making it an ideal target for the autoimmune response. The thyroglobulin gene is found on chromosome 8q24, and whole genome studies in Japan and the United States were the first to correlate this area to HT and autoimmune thyroid disease. Thyroglobulin gene was discovered to be one of the primary thyroid specific susceptibility genes, related and associated with autoimmune thyroid disease, after precise mapping of this region [10]. Later, different alleles of several microsatellite markers and SNPs in the thyroglobulin gene were linked to HT, potentially changing its expression, antigenicity, iodination, or HLA binding. In both Japanese and Caucasian populations, the relationship of the Tgms2 microsatellite marker in intron 27 with HT was verified. Human thyroglobulin sequencing showed 14 SNPs, four of which were linked to HT, including the exon 10-12 SNP cluster and exon 33 SNP [9].
Spheroid Models
A 3D bioprinter was used to produce thyroid and allantoic spheroids in a collagen hydrogel in 2017. The construct was placed under the renal capsule of hypothyroid mice. After grafting, they noticed vascularization of the construct, as well as restored thyroxine levels and body temperature, indicating functional thyroid regeneration [19]. Bioprinting is the additive biofabrication of three-dimensional (3D) tissues and organ structures using self-assembling tissue spheroids as the building blocks. The thyroid gland is a simple endocrine organ that can be used to test the suggested bioprinting method. A functioning vascularized mouse thyroid gland construct was bioprinted using embryonic tissue spheroids [20]. Thyroid tissue was created using the self-assembly principle, using thyroid spheroids (TS) and allantoic spheroids (AS) as sources of thyrocytes and endothelial cells, respectively. Closely placed embryonic tissue spheroids merged into a single integral construct during the culture, EC from AS invaded and vascularized TS, and epithelial cells from the TS gradually produced follicles. Observed the establishment of a capillary network surrounding follicular cells in this experimental scenario, similar to what we see during in utero thyroid development when the thyroid epithelium controls the recruitment, invasion, and extension of EC around follicles [21]. Reduced endogenous EC from TS before bioprinting to prove that EC from AS are responsible for vascularization of the thyroid gland construct. Thyroid tissue that had been decreased was totally revascularized with EC from AS. After being grafted under the kidney capsule of hypothyroid mice, the cultured bioprinted construct was able to normalize blood thyroxine levels and body temperature. Bioprinting of a functioning vascularized mouse thyroid gland construct is a step forward in bioprinting technology, allowing researchers to investigate the self-assembling capabilities of tissue spheroids [20].
Thyroid Organoids
During embryonic development, the thyroid is formed from the same progenitor cells as the lungs. According to the initial report, temporary overexpression of Pax8 and Nkx2-1 was sufficient to create thyroid organoids from mouse ESCs. Longmire’s methodology also produced thyroid organoids by sequentially exposing progenitors to BMP/FGF4, then plating the cells in 3D architecture in Matrigel with thyroid stimulating hormone (TSH) to stimulate thyroid organoids formation [15]. Kurmann, et al. [15] developed a methodology for manufacturing thyroid organoids using mouse PSCs based on this protocol. Thyroid organoids were generated in this fashion, and when transplanted into hypothyroid mice, they secreted thyroid hormones and rescued the mice. The same procedure could be used to produce human thyroid organoids from iPSCs. Kurmann described a method for long term growth of thyroid organoids capable of iodide uptake and thyroglobulin synthesis. Normal thyroid-like tissues emerged after these organoids were implanted into a mouse model of hypothyroidism [15]. InVitro Human Thyroid Model Hundreds of potential TDCs have been identified due to significant advances in the development and implementation of HTS tests for thyroid-related MIEs. An in vitro screening model that more accurately reflects the thyroid gland intrinsic biology could be useful for orthogonal confirmation of tissue level effects on the TH production pathway, lowering the uncertainty associated with HTS experiments [22]. The physical and functional properties of 2D and 3D thyroid culture models were characterized using primary human thyrocytes obtained from euthyroid donors. Some features of native thyroid function, such as TSH-dependent gene expression and TG synthesis, were able to be replicated by thyrocytes in a standard 2D monolayer. The 2D culture format, on the other hand, was definitely lacking in all essential components for thyroid hormonogenesis. In a stable, long-term in vitro culture model, adaptation to a 3D hydrogel culture format using medium optimized for human thyrocytes restored synthesis of iodothyronine hormones for up to 20 days [23]. Furthermore, by developing the 3D microtissue model in a 96-well culture format, inhibition of TH production could be reproducibly investigated using a known TDC reference set to directly test the efficacy and potency of TH disruption in a concentration-dependent manner. Primary cells, unlike many immortalized cell lines, have stringent criteria for successful cell culture development. The composition of one of the most regularly used primary thyroid cell culture medium formulations was originally designed for FRTL rat thyrocytes [23]. Insulin, hydrocortisone, transferrin, glycol-L-histidyl- L-lysine acetate, somatostatin, and TSH are included in the formulation, which is made up of Ham’s F-12 base medium supplemented with calf serum and six additives. TSH levels greater than the specified concentration of 0.04 mU/mL were required for maximum performance, particularly in the 3D culture format, to maximize cellular responses. When cells were maintained at significantly higher levels of 1 mU/ml TSH, the effects on gene expression, TG protein production, and TH output were most consistent, hence this concentration in h7H media was judged best for optimizing the dynamic range in the 3D microtissue test [22].
Impact of Thyroid Autoantibodies Positivity on InVitro Fertilization
The female reproductive system’s functionality could be significantly impacted by thyroid disease through a variety of methods. Menstrual abnormalities, infertility, anovulation, and poor obstetric outcomes are all linked to overt thyroid illnesses such Hashimoto thyroiditis. Thyroid autoimmunity is the most common pathologic causing thyroid dysfunction in women of reproductive age. When compared to the general population, infertile women have a higher prevalence of thyroid autoimmune dysfunction [11]. However, there is a lack of evidence on the direct influence of TAA positivity on IVF success, particularly in the community of infertile, euthyroid TAA positive women. When overt thyroid disease or subclinical hypothyroidism coexist, maternal TAA positive may affect IVF/ICSI results. It’s likely that TAA doesn’t directly influence oocyte development, embryo implantation, or embryo quality, but rather compromises the physiological processes of implantation through Th1- mediated cytotoxicity [9].
Protection against Thyroid Autoimmunity
Self-tolerance is maintained through two ways. The thymic elimination of auto reactive T cells during fetal life is referred to as central tolerance. Those cells that escape central tolerance are prevented from causing autoimmunity by peripheral tolerance mechanisms, in which Tregs play a critical role. They suppress effector T cells, APCs, and B cells, maintaining immunological unresponsiveness to self- antigens and reducing an overactive immune response. They can suppress target cells directly or through suppressor cytokines that are produced. Tregs are T cells that come from the thymus or naive T cells in the periphery, and they express CD25 (the IL-2 receptor chain) and FoxP3. As a result, CD4 CD25 Foxp3 Tregs play a crucial function in the immune system [12]. Regulatory suppressive effects are caused by cells with the highest CD25 expression (CD4+CD25high). The significance of Tregs in self-tolerance has been widely explored in animal models, particularly in mouse experimental autoimmune thyroiditis, where thymically- derived CD4 CD25 Foxp3 Tregs provided protection against autoimmunity. CD25 expression was higher in HT patients than in healthy persons in humans. The proportion of Tregs was lower intrathyroidal than in peripheral blood in patients with autoimmune thyroid disease [13]. Three-color flow cytometry revealed a larger proportion of special Tregs with high levels of FoxP3 in HT patients when compared to controls. This points to a compensatory increase in the Treg subgroup in order to reduce the immunological response. TGF-β induces the expression of Foxp3 and the production of Treg cells in Tregs, fibroblasts, macrophages, endothelial cells, and thyrocytes in inflammatory thyroid tissue [13]. TGF-β promotes suppressive Tregs and inhibits T cell development, making it a major regulator of immunological tolerance. As a result, serum levels of TGF-β were lower in HT patients than in controls, and did not change after therapy with levothyroxine. As a result, TGF-β levels appear to be linked to HT rather than thyroid dysfunction. TGF-β has been shown to have a suppressive impact in the development of experimental autoimmune thyroiditis. TGF-β may trigger the development of fibrosis in the latter stages of thyroiditis [12].
Conclusion
HT is one of the most common autoimmune disorders, which can be triggered by a variety of factors in genetically predisposed people. Multiple susceptibility genes, some of which are also found in other autoimmune disorders but are unique to thyroid autoimmunity, may be implicated in disease development. Immune-regulatory genes including HLA, CTLA-4, and PTPN22 are now known to play a role in the etiology of HT. Furthermore, recent research has linked cytokine genes like IFN-, IL-4, and TGF-β to the development and severity of HT, implying that they influence the balance between Th1 and Th2 processes. According to current knowledge, a complicated combination between hereditary and non-genetic variables leads to increased thyroid antigen presentation and decreased immune tolerance, resulting in Th1-type autoimmunity, thyroid destruction, and clinical illness. Despite significant progress in our understanding of HT during the last decade, the exact processes of its growth remain unknown. Hopefully, new information will provide a better understanding of illness causation in the near future, allowing us to detect at- risk individuals and potentially prevent the onset of clinical disease.
References
-
Malkowski B, Serafin Z, Glonek R, Suwała S, Lopatto R, et al. (2019) The role of 18F-FDG PET/CT in the management of the autoimmune thyroid diseases. Frontiers in endocrinology 10: 208.
-
Aurizio F, Villalta D, Metus P, Doretto P, Tozzoli R (2015) Is vitamin D a player or not in the pathophysiology of autoimmune thyroid diseases. Autoimmunity reviews 14(5): 363-369.
-
Brown J, Solomon DH, Beall GN, Terasaki PI, Chopra IJ, et al. (2012) Autoimmune thyroid diseases—Graves’ and Hashimoto’s. Annals of Internal Medicine 88(3): 379- 391.
-
Pastuszak D, Sewerynek E, Domanska D, Gładys A, Skrzypczak R, et al. (2012) CTLA-4 gene polymorphisms and their influence on predisposition to autoimmune thyroid diseases (Graves’ disease and Hashimoto’s thyroiditis). Arch Medical Sci 8(3): 415-421.
-
Iddah MA, Macharia BN (2013) Autoimmune thyroid disorders. International Scholarly Research Notices.
-
Cappa M, Bizzarri C, Crea F (2011) Autoimmune thyroid diseases in children. Journal of thyroid research.
-
Yamada H, Itoh M, Hiratsuka I, Hashimoto S (2014) Circulating micro RNA s in autoimmune thyroid diseases. Clin Endocrinol 81(2): 276-281.
-
Kawicka A, Regulska B (2015) Metabolic disorders and nutritional status in autoimmune thyroid diseases. Postepy Hig Med Dosw 69: 80-90.
-
Chen H, Hou Y, He J, Cheng Z, Wang R (2014) A study on the clinical effect and immunological mechanism in the treatment of Hashimoto’s thyroiditis by moxibustion. J Tradi Chin Med 13(1): 14-18.
-
Eschler DC, Hasham A, Tomer Y (2014) Cutting edge: the etiology of autoimmune thyroid diseases. Clin Rev Allergy Immunol 41(2): 190-197.
-
Toulis KA, Anastasilakis AD, Tzellos TG, Goulis DG, Kouvelas D (2010) Selenium supplementation in the treatment of Hashimoto’s thyroiditis: a systematic review and a meta-analysis. Thyroid 20(10): 1163-1173.
-
Pearce EN, Farwell AP, Braverman LE (2003) Thyroiditis. New England Journal of Medicine 348(26): 2646-2655.
-
Taneri F, Yuksel O, Aydin A, Tezel E, Onuk E (2019) Total thyroidectomy for the treatment of Hashimoto’s thyroiditis coexisting with papillary thyroid carcinoma. Adv Ther 24(3): 510-516.
-
Antonica F, Kasprzyk DF, Opitz R, Iacovino M, Liao XH, et al. (2012) Generation of functional thyroid from embryonic stem cells. Nature 491(7422): 66-71.
-
Kurmann AA, Serra M, Hawkins F, Rankin SA, Mori M, et al. (2015). Regeneration of thyroid function by transplantation of differentiated pluripotent stem cells. Cell Stem Cell 17(5): 527-542.
-
Hollenberg AN, Choi J, Serra M, Kotton DN (2017) Regenerative therapy for hypothyroidism: mechanisms and possibilities. Mol Cell Endocrinol 445: 35-41.
-
Weeber F, Ooft SN, Dijkstra KK, Voest EE (2017) Tumor organoids as a pre-clinical cancer model for drug discovery. Cell Chem Biol 24(9): 1092-1100.
-
Ma Risheng, Morshed SA, Latif R, Davies TF (2015) Thyroid cell differentiation from murine induced pluripotent stem cells. Front Endocrinol 6: 56.
-
Bulanova EA, Koudan EV, Degosserie J, Heymans C, Pereira FD, et al. (2017) Bioprinting of a functional vascularized mouse thyroid gland construct. Biofabrication 9(3): 034105.
-
Hornung MW, Korte JJ, Olker JH, Denny JS, Knutsen C, et al. (2018) Screening the ToxCast phase 1 chemical library for inhibition of deiodinase type 1 activity. Toxicol Sci 162(2): 570-581.
-
Kohling HL, Plummer SF, Marchesi JR, Davidge KS, Ludgate M (2017) The microbiota and autoimmunity: Their role in thyroid autoimmune diseases. Clin Immunol 183: 63-74.
-
Kosian PA, Haselman JT, Korte JJ, Challis K, Macherla C, et al. (2015) In vitro, Ex vivo, and in vivo determination of thyroid hormone modulating activity of benzothiazoles. Toxicol Sci 146(2): 254-264.
-
Degitz SJ, Korte LM, Olson JM, Kosian PA, Linnum AL, et al. (2010) Inhibition of thyroid hormone release from cultured amphibian thyroid glands by methimazole, 6-propylthiouracil, and perchlorate. Toxicolo Sci 118(1): 42-51.
- Origin, Evolution, and Functional Impact of Short Insertion- Deletion Variants in Human Genomes: A Review
- Harnessing Molecular Glues for Next-Generation Vaccine, Cancer and Cardiovascular Disease Drug Development: A Comprehensive Review
- Lateral Cervical Epidermal Inclusion Cyst in a Paediatric Patient: A Rare Case Report
- Malarial Plasmodium Falciparum with Hepatitis B and C Virus Infections among Blood Donors in Ife Central Local Government Area, Ile Ife, Osun State, Nigeria
- Withanolides and Withaferin A- What’s next in Ashwagandha Research
- Designing of Dual Pulse Photoacoustic Tomography for Imaging of Drug-Response and Tumor Growth