Retarded Swarming Motility in Bacillus Subtilis NRS-762 and Pseudomonas Aeruginosa PRD-10
Coping with nutritional stress is essential for cell survival, of which many strategies at the cellular level lend support for ensuring the survival of the population at a particular habitat. One postulated mechanism is swarming motility in bacterial cells, where, upon depletion of nutrients at a locale, cells would coordinate their movement, synthesize more flagella, and secrete lubricants for moving rapidly across surfaces in search for food. Known to engage in swarming motility, Bacillus subtilis and Pseudomonas aeruginosa are two common bacterial species with versatile metabolism that use the motility mode to colonize new habitats with more favourable environmental and nutritional conditions. However, experimental observations of bacteria growth on a variety of agar media revealed that B. subtilis NRS-762 (ATCC 8473) and P. aeruginosa PRD-10 (ATCC 15442) exhibited retarded swarming motility upon entry into stationary phase on solid media. Specifically, B. subtilis NRS-762 colonies exhibited round, wrinkled morphologies compared to complex filamented swarming patterns common in strains able to engage in swarming motility. On the other hand, P. aeruginosa PRD-10 colonies were round, mucoid, and expanded outwards from the colony centre without extending filaments from the centre; thereby, indicating retarded swarming motility. Thus, impaired cellular machinery for swarming motility or mutated and deleted genes likely account for observed retarded swarming motility in B. subtilis NRS-762 and P. aeruginosa PRD-10. More importantly, observations of small filaments extending radially from an expanded colony of P. aeruginosa PRD-10 grown on minimal salts medium supplemented with yeast extract highlighted possible loss of function of effector molecules that transmit cellular decision at swarming motility into movement, while sensory mechanisms feeding into the motility mechanism remained intact. More broadly, observations of impaired swarming motility in B. subtilis NRS-762 and P. aeruginosa PRD10 in two species otherwise endowed with the motility mode highlighted the presence of additional triggers for swarming motility are likely present, and the motility mode may have been evolutionary selected for other functions in addition to foraging for food in times of nutritional stress Graphical Abstract.
Short Description
Microbes activate sensory and effector mechanisms in search for food during nutritional stress. One such mechanism is swarming motility. Known as collective coordinated movement of microbes over solid surfaces, detailed mechanisms underlying the activation and coordination of movement between cells remain poorly understood. Observations of retarded swarming motility in Bacillus subtilis NRS-762 and Pseudomonas aeruginosa PRD-10, on a variety of agar media, however, highlighted that the motility mode is not universally conserved in all strains of a species endowed with the capability. Hence, swarming motility may not be essential as a cellular differentiation pathway for mediating food foraging during nutritional stress, and other as-yet unknown triggers and functions may be associated with the motility mechanism.
Highlights
1. Retarded Swarming motility was observed in growth of Bacillus subtilis NRS-762 (ATCC 8473) and Pseudomonas aeruginosa PRD-10 (ATCC 15442) on a variety of agar media. 2. Colonies of the two strains were round and did not exhibit characteristic Swarming filaments typical of Swarming motility, even after prolonged incubation.
3. Thus, absence of swarming motility in strains of bacterial species known to be endowed with the motility mode, thought to be important for food foraging in times of nutritional stress, highlighted that the motility mode may not be essential to survival of the species. 4. Additional as-yet unknown triggers and functions could be associated with Swarming motility as an evolutionary selected motility mode. 5. Presence of retarded filaments radiating outward from round colonies of P.aeruginosa PRD-10 during growth on minimal salts medium supplemented with yeast extract revealed that effector mechanism of swarming motility in the strain could be impaired, while sensory and decision making mechanisms of the motility mode remained intact.
Introduction
Lack of nutrients in a habitat could usually prompt cellular differentiation for motility in search for food. Such motility could manifest in a number of ways, one of which is swarming motility [1]. Specifically, swarming motility is the collective coordinated motion of cells in a directional manner over solid surfaces for colonizing new habitats, for example, in search of nutrients such as during nutrient starvation in stationary phase [2, 3], Graphical Abstract.
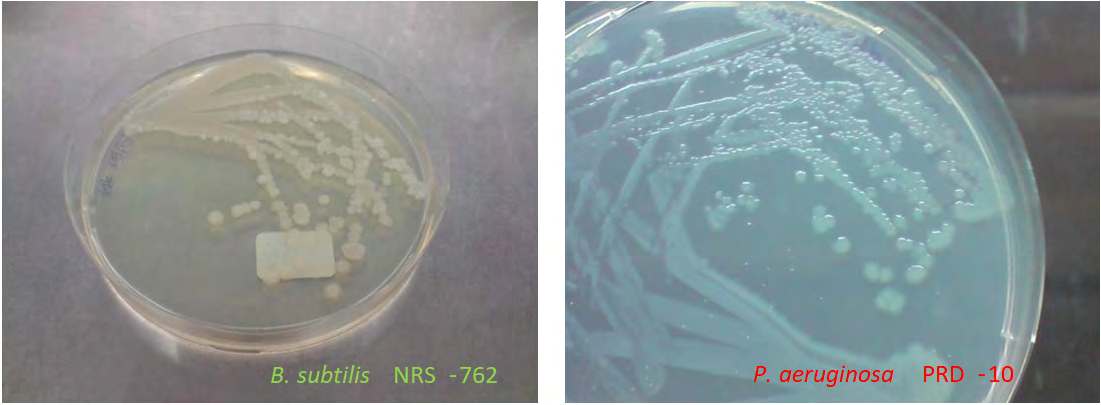
Graph 1: Graphical Abstract.
Bacillus subtilis, a common soil bacterium with a versatile metabolism and ability to inhabit diverse habitats is known to engage in swarming motility as a cellular differentiation programme in search of nutrients during nutritional stress. Upon activation of swarming motility, multiple flagella would be produced in a single cell.1 Bundled together, these flagella propel the bacterial cell forward on a surface lubricated by surfactant secreted by the swarming cells [1, 4]. Controlled by a sensory system that likely communicates with neighbouring cells, swarming motility typically result in complex patterns on solid medium upon entry of the culture into stationary phase. Many unknowns remain in the area of swarming motility research, such as the biophysical and biochemical underpinnings of the complex patterns on agar and their relation to cell-cell communications. Similarly, another common bacterium, Pseudomonas aeruginosa, also engages in swarming motility when grown on a variety of rich complex media agar. Typically, transition to swarming motility happens upon nutrient depletion in the growth environment, where multiple flagella are synthesized and bundled together for motility in a directional manner guided by the search for nutrients [1, 5]. Lubricants for motility on agar surface were also secreted by the bacterium for facilitating rapid movement typical in swarming motility, where a collection of cells engage in coordinated movement [1, 5, 6, 7, 8, 9]. However, multiple aspects of the sensory and cellular decision making effectors in swarming motility remain poorly understood. While the species, Bacillus subtilis, is known to engage in swarming motility as a form of cellular differentiation, experimental results with the culture of B. subtilis NRS-762 (ATCC 8473) suggested that the strain could not engage in swarming motility during growth on various agar media. Specifically, no complex motility patterns were observed in culture of B. subtilis NRS-762 on agar, where such patterns are known to exist in culture of other Bacillus subtilis strains. Hence, B. subtilis NRS-762 exhibited retarded swarming motility that manifest as round, wrinkled colonies, which could be due to impaired genetics controlling the cellular differentiation programme such as the presence of mutated genes that led to ineffective gene products unable to partake in the sensory and control of the motility mode. Other studies have highlighted that most laboratory strains of B. subtilis do not exhibit swarming motility [4, 10]. Similarly, retarded swarming motility was also observed in Pseudomonas aeruginosa PRD-10 (ATCC 15442) given the lack of complex motility patterns during growth on LB Lennox and other solid media. Specifically, round, wrinkled colonies of P. aeruginosa PRD-10 were observed during growth on various agar media after the culture reached stationary phase. Collectively, observations of retarded swarming motility in two strains of common bacteria, B. subtilis NRS-762, and P. aeruginosa PRD-10 suggested that the motility mode may not be absolutely conserved across all strains of a species; thereby, suggesting that the motility mode may not be evolutionary critical for rapid search of nutrients upon nutrient starvation. Hence, swarming motility as a complex motility mechanism might harbour other evolutionary underpinnings critical for as-yet unknown functions important for coping with environmental stressors such as nutrient depletion. Instead of using a rapid motility mode for searching more favourable habitats, observations of gradual outward growth of B. subtilis NRS-762 and P. aeruginosa PRD- 10 colonies that led to colony expansion and colonization of more nutritional sources, suggested that swarming motility may have triggers other than nutritional stress [2].
Materials and Methods
Materials
Composition of media in [g/L], LB Lennox: Tryptone, 10.0; Yeast extract, 5.0; NaCl, 5.0; LB Lennox agar: Tryptone, 10.0; Yeast extract, 5.0; NaCl, 5.0; Agar, 15.0; MM1: D-Glucose, 4.0; NH4Cl, 1.0; K2HPO4, 0.5; KH2PO4, 0.1; NaCl, 0.5; Agar, 15.0; MM2: D-Glucose, 2.0; NH4Cl, 0.5; K2HPO4, 0.5; KH2PO4, 0.1; NaCl, 0.5; MgSO4.7H2O, 1.0, Yeast extract, 1.0; Agar, 15.0; MM3: D-Glucose, 2.0; NH4Cl, 0.5; K2HPO4, 0.5; KH2PO4, 0.1; NaCl, 0.5; MgSO4.7H2O, 1.0, Yeast extract, 0.1; Agar, 15.0; MM4: D-Glucose, 2.0; NH4Cl, 0.5; K2HPO4, 0.5; KH2PO4, 0.1; NaCl, 0.5; MgSO4.7H2O, 1.0, Yeast extract, 0.5; Agar, 15.0; Culture of Bacteria on Solid Media Glycerol stock cultures of Bacillus subtilis NRS-762 and Pseudomonas aeruginosa PRD-10 were prepared in glycerol and stored at -70 oC until use. Glycerol stock cultures of both bacteria were used in inoculating 100 mL of LB Lennox medium in 250 mL glass shake flasks as starter cultures, where B. subtilis NRS-762 was incubated at 30 oC and 230 rpm rotational shaking, and P. aeruginosa PRD-10 was incubated at 37 oC and 230 rpm rotational shaking. After 24 hours of incubation, the two respective cultures were used as seed cultures for inoculating different agar media via the streak plate method. Inoculated agar plates were incubated at 30 oC for B. subtilis NRS-762 and 37 oC for P. aeruginosa PRD-10.
Results and Discussion
Growth of B. subtilis NRS-762 on various agar media did not manifest in growth patterns suggestive of swarming motility (Figure 1). Specifically, round, wrinkled colonies were observed, even after multiple days of incubation on agar media; thus, swarming motility might be impaired in the strain. More importantly, the colonies grew by expanding outwards from the centre of the colony, which was not a rapid approach for nutrient search in times of nutritional stress such as stationary phase (Figure 2). Hence, swarming motility might not be important for helping the clonal population forage for nutrients in a nutrient depleted environment as conventional outward growth of colony could provide new sources of nutrients adjacent to the colony [11]. Therefore, rapid search for new sources of nutrients as hypothesized for swarming motility [1] might not be important for overall clonal population maintenance, which suggested that swarming motility might have as yet unknown triggers and functions.
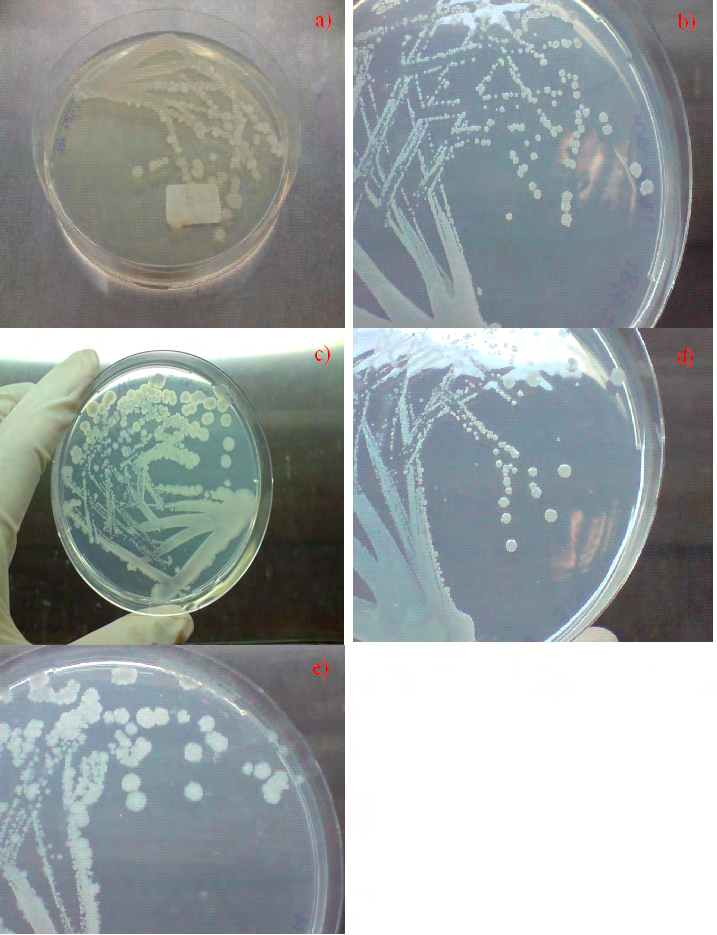
Figure 1: Growth of Bacillus subtilis NRS-762 on various agar at 30 oC. a) LB Lennox for 1 day of incubation, b) MM1 medium for 6 days of incubation, c) MM2 medium for 1 day of incubation, d) MM3 medium for 5 days of incubation, and e) MM4 medium for 1 day of incubation. Note the round, wrinkled colonies that did not exhibit complex swarming motility patterns upon entry into stationary phase.
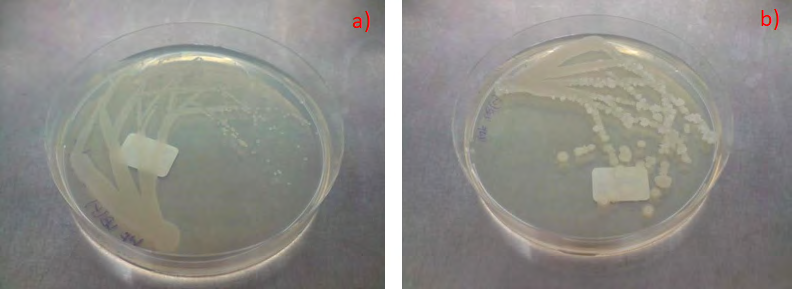
Similar to the case for B. subtilis NRS-762, Pseudomonas aeruginosa PRD-10 (ATCC 15442) was cultivated on a variety of agar media at 37 oC for different periods of time (Figure 3). All cultures reached stationary phase where there was depletion of nutrients, but no swarming motility was observed. Specifically, colonies were round, mucoid, and expanded in size from the colony centre with culture time (Figure 4). Such colony expansion did not require enhanced motility of the bacterium similar to that in swarming motility. On the other hand, evidence of retarded swarming motility could be seen in growth of P. aeruginosa PRD-10 on MM4 medium, where after 7 days of incubation, filaments extended radially from the colony centre (Figure 3e). Such growth pattern resembled that of initial phases of swarming motility but was not in sync with the cellular differentiation pathway that typically manifest in complex motility patterns [1]. Thus, retarded swarming motility was exhibited by P. aeruginosa PRD-10 that suggested that effector molecules that translate cellular decisions of swarming motility into movement did not function normally, while cellular apparatus that translates sensory information about nutritional stress into cellular decision for swarming motility likely remained intact.
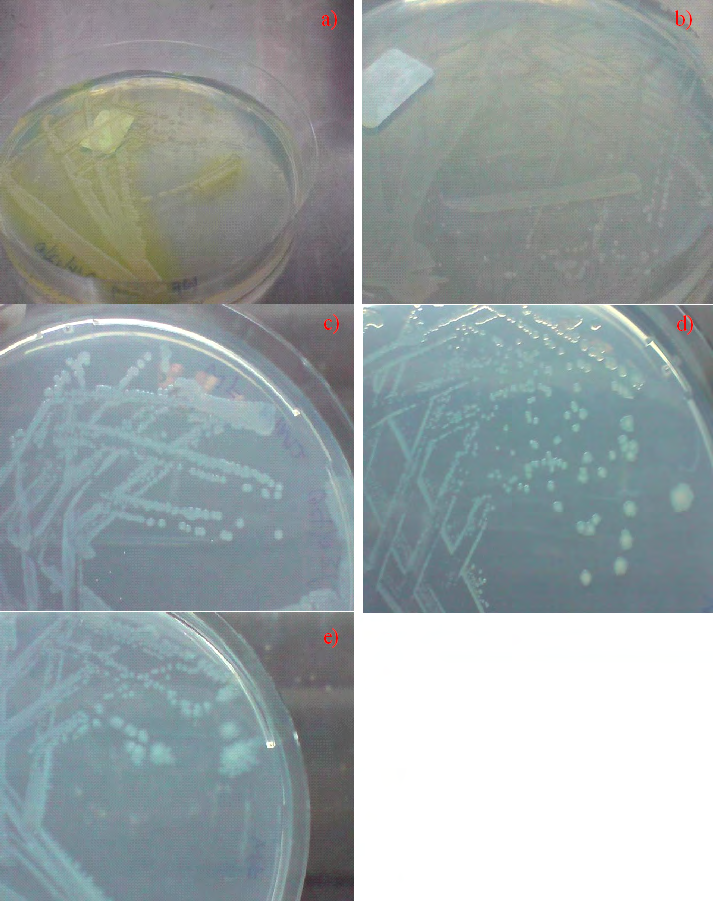

Taken together, both B. subtilis NRS-762 and P. aeruginosa PRD-10 exhibited retarded swarming motility during growth on a variety of agar media, where observations revealed round, wrinkled colonies for B. subtilis NRS-762, and round, mucoid colonies for P. aeruginosa PRD-10 even after several days of incubation. B. subtilis NRS-762 and P. aeruginosa PRD- 10 colonies expanded from the centre of the colony outwards; thereby, suggesting that swarming motility present in many strains of the two species were arrested in the two specific strains used in this study due possibly to genetic impairment of the motility pathway [12, 13]. Observations of the growth behaviour of B. subtilis NRS-762 and P. aeruginosa PRD-10 also suggested that swarming motility’s role may not be solely for rapid motility in search for nutritional sources during nutrient depletion [14]. More importantly, as-yet unknown triggers or stressors may evoke swarming motility in bacterial species endowed with the cellular differentiation pathway. P. aeruginosa PRD-10, however, offers an interesting look into possible loss of function in effector molecules of swarming motility that translates the cellular decision to enter the differentiation pathway into cell movement, while the sensory and control system supporting swarming motility decision-making remained intact.
Conclusion
Swarming motility is known as a bacterial motility mode supporting the acquisition of nutrients in new habitats when nutritional stress sets in. However, observations of retarded swarming motility in two strains of bacteria belonging to species known to engage in the motility mode upon entry into stationary phase highlighted the possibility that evolutionary forces may have selected swarming motility as a survival motility mechanism may have chosen other triggers that, together, feeds into the cellular decision matrix for the differentiation pathway. In essence, the primary role of swarming motility may not be in searching for new habitats with more favourable environmental and nutritional conditions. Rather, the motility mode could be a form of self-defence against other species that manifested in the rapid colonization of more habitats upon reaching a threshold population such as that during stationary phase. Both B. subtilis NRS-762 and P. aeruginosa PRD10 exhibited retarded swarming motility that suggested that different aspects of the cellular machinery for swarming motility may be impaired. In the case of P. aeruginosa PRD-10, effector mechanisms that turn cellular decisions for swarming motility into action may be significantly affected, which resulted in retarded swarming motility. On the other hand, the sensory and control mechanisms for swarming motility likely remained intact in P. aeruginosa PRD-10.
References
-
Kearns DB (2010) A field guide to bacterial swarming motility. Nat Rev Microbiol 8: 634-644.
-
Patrick JE, Kearns DB (2011) Swarming motility and the control of master regulators of flagellar biosynthesis. Mol Microbiol 83(1): 14-23.
-
Ang S, Horng YT, Shu JC, Soo PC, Liu JH, et al. (2001) The role of RsmA in the regulation of swarming motility in Serratia marcescens. J Biomed Sci 8(2): 160-169.
-
Kearns DB, Richard L (2004) Swarming motility in undomesticated Bacillus subtilis. Mol Microbiol 49(3): 581-590.
-
Ha DG, Kuchma SL, Toole OGA (2014) Plate-Based Assay for Swarming Motility in Pseudomonas aeruginosa. Methods Mol Biol 1149: 67-72.
-
Tremblay J, Déziel E (2010) Gene expression in Pseudomonas aeruginosa swarming motility. BMC Genomics 11(587): 1-15.
-
Oura H, Tashiro Y, Toyofuku M, Ueda K, Kiyokawa T, et al. (2015) Inhibition of Pseudomonas aeruginosa Swarming Motility by 1-Naphthol and Other Bicyclic Compounds Bearing Hydroxyl Groups. Appl Environ Microbiol 81(8): 2808-2818.
-
Kadosh BI, Zilka A, Yerushalmi G, Banin E (2013) The Effect of pstS and phoB on Quorum Sensing and Swarming Motility in Pseudomonas aeruginosa. PLOS ONE 8(9): e74444.
-
Chihiro T, Nozawa T, Tamikawa T, Nakagawa Y, Wakita J, et al. (2008) Swarming of Pseudomonas aeruginosa PAO1 without differentiation into elongated hyperflagellates on hard agar minimal medium. FEMS Microbiol Lett 280(2): 169-175.
-
Patrick JE, Kearns DB (2009) Laboratory Strains of Bacillus Subtilis Do Not Exhibit Swarming Motility. J Bacteriol 191(22): 7129-7133.
-
Tronnolone H, Tam A, Szenczi Z, Green JEF, Balasuriya S, et al. (2018) Diffusion-Limited Growth of Microbial Colonies. Scientific Reports 8(5992): 1-11.
-
O Rear J, Alberti R, Harshey M (1992) Mutations that impair swarming motility in Serratia marcescens 274 include but are not limited to those affecting chemotaxis or flagellar function. J Bacteriol 174(19): 6125-6137.
-
Joerg O, Shawn L, Alexandra MK, Robert HEW (2007) Identification of Genes Involved in Swarming Motility Using a _Pseudomonas aeruginosa_ PAO1 Mini-Tn5-lux Mutant Library. Journal of Bacteriology 189(5): 2164- 2169.
-
Remy C, Bin N, Leanid L, Victor S (2021) Multiple functions of flagellar motility and chemotaxis in bacterial physiology. FEMS Microbiology Reviews 4 5 ( 6 ) : 038.
- Origin, Evolution, and Functional Impact of Short Insertion- Deletion Variants in Human Genomes: A Review
- Harnessing Molecular Glues for Next-Generation Vaccine, Cancer and Cardiovascular Disease Drug Development: A Comprehensive Review
- Lateral Cervical Epidermal Inclusion Cyst in a Paediatric Patient: A Rare Case Report
- Malarial Plasmodium Falciparum with Hepatitis B and C Virus Infections among Blood Donors in Ife Central Local Government Area, Ile Ife, Osun State, Nigeria
- Withanolides and Withaferin A- What’s next in Ashwagandha Research
- Designing of Dual Pulse Photoacoustic Tomography for Imaging of Drug-Response and Tumor Growth