Flavin Monooxygenases: A Multifaceted Class of Enzymes with Abundant Biomedical Applications
Flavin-dependent monooxygenases (FMOs) catalyze oxidation of a variety of organic molecules many of which have a profound biomedical applications. Oxidation occurs through the incorporation of a single atom of dioxygen into the substrate while the other oxygen atom is reduced to water. Since 2014, FMOs are grouped into eight types listed as Groups A through H. Grouping is based on structural homology, flavin type and the strategy taken to reduce the isoalloxazine ring of the cofactor for subsequent oxygen activation and substrate monooxygenation. Groups A and B FMOs are single component enzymes that carry out their entire reaction cycle in a single protein core using a nicotinamide adenine dinucleotide coenzyme as electron source. Groups C through F are two comments FMOs. These systems contain a reductase to reduce the flavin cofactor that is then delivered to a monooxygenase component as a substrate. Reduced flavin then activates molecular oxygen in the second component to oxidize the organic metabolite of the system. Finally, Groups G and H are self- sufficient internal monooxygenases that receive electrons from one substrate for the oxidation of a second one all within a single protein unit. FMOs carry out an array of biomedically relevant reactions including the degradation of antibiotics by pathogens, the biocatalytic production of drugs in pharmaceutical industries and cholesterol anabolism in humans and fungi. Given this diversity of both reaction type and application, the field of FMO enzymology can rapidly become overwhelming. This review provides a general overview of each FMO group and is intended to guide researchers just entering the field. Canonical enzymes are detailed along with biomedically important members of each group. It is hoped the references herein will be provide a quick source to make more extensive literature reviews more accessible.
Introduction
In 1947, N.L. Edson described the isolation of an enzyme from Mycobacterium phlei that catalyzes the oxidative decarboxylation of lactate [1]. The enzyme became known as lactate 2-monooxygenase (E.C. 1.13.12.4) and represents the first flavin dependent monooxygenase (FMO) ever isolated [2]. Since Edson’s discovery around 250 FMOs have been identified, at least partially purified and described to carry out a vast array of biochemical reactions [3]. These range from the catabolism of xenobiotics or natural products to the anabolism of vitamins, hormones and antibiotics. FMOs have been studied for their roles in essential metabolic pathways, the biophysics of light emission or dioxygen activation and their remarkably rich redox chemistry. Many excellent and comprehensive reviews have been written for this multifaceted class of enzymes [2, 3, 4, 5, 6, 7, 8] and it is easy to quickly become overwhelmed by the thousands of research articles related to FMO chemistry and applications.
The current review is intended to orient researchers new to the field of flavoprotein enzymology to the fundamental chemistry of the monooxygenases and will emphasize some of the biomedical applications of FMOs. Reviews covering other flavoprotein classes such as the oxidases, reductases or dehydrogenases can be found elsewhere [6, 7]. Broadly speaking, FMOs are enzymes that incorporate a single oxygen atom from O2 into an organic substrate while reducing the other atom to water [3]. They fall into either the 1.13 or 1.14 groups of the Enzyme Classification (E.C.) system set by the International Union of Biochemistry and Molecular Biology [9]. That is they are oxidoreductases (1.) that act on either a single (13.) or a paired electron donor (14.). They have been further classified into eight groups (A through H) by van Berkel and coworkers [3, 5] as detailed below.
FMO reactions involve the participation of either flavin adenine dinucleotide (FAD) or flavin mononucleotide (FMO) as a cofactor or substrate. Figure 1 shows the isoalloxazine ring of flavins which is the site and source for the chemical versatility of FMOs. The majority of FMOs are reduced by nicotinamide adenine dinucleotide [NAD(P)H] coenzymes (Figure 1) during the reductive half reactions of their catalytic cycle. As reviewed extensively elsewhere [6, 7], they catalyze a highly controlled monooxygenation of an organic substrate in their oxidative half reaction. Distinct UV-visible and fluoresce properties of the different redox state of the isoalloxazine ring and the ability to independently study each half reaction have led to a deep and comprehensive understanding of flavin chemistry. These studies were largely pioneered during the career of Vincent Massey at the University of Mcguigan [10] who helped developed sophisticated rapid kinetic techniques to study flavoenzymes. The protein matrix of FMOs activate the isoalloxazine ring with oxygen to give rate constants as high as 106 M-1s-1 (as opposed only 250 M-1s-1 for free flavins). The flavoenzyme also ensures minimal release of destructive reactive oxygen species or wasteful uncoupled reactions of the flavin with molecular oxygen.
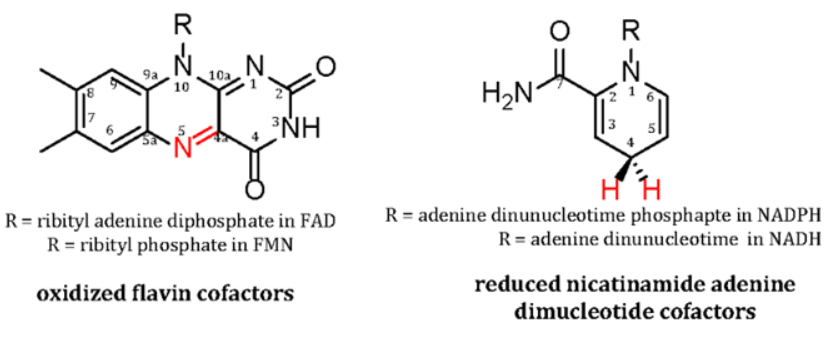
FMOs achieve a safe and efficient mode of catalysis typically by forming a C4(a)-(hydro)peroxy flavin- oxygen adduct (Figure 2). Until 2013 when the oxidative half reaction of EnzM [11], an enzyme involved in the biosynthesis of the antibiotic enterocin, was studied, it was thought that all FMOs oxidize substrates through flavin C4a-oxygen intermediates. EnzM, and a few enzymes studied since [2, 6], oxidize substrates through the formation of flavin N5-oxides. The discovery of a novel mechanisms even 65 years after the first FMO was isolated illustrates the rich chemistry afforded by the isoalloxazine ring of flavin cofactors. Since we intend to only provide a general overview of FMO classification and their biomedical importance, only the most common adducts are shown in Figure 2 will be discussed in this review. These represent over 95% of known FMOs mechanistically characterized to date [2]. The formation of either C4a-peroxy or C4a-hydroperoxyflavins depends solely on the nature of the organic substrate that the FMO
evolved to oxidize. Electron poor (electrophilic) substrates are monooxygenated by nucleophilic peroxyflavins whereas electron rich (nucleophilic) substrates are oxidized by hydroperoxy flavins that are electrophilic.
![Figure 2: Most Common Flavin-Oxygen Adducts in Flavin Monooxygenases Flavin reduction occurs in one of the three general ways illustrated in Figure 3. The reduced flavin then reacts with molecular oxygen to generate a highly reactive adduct that monooxygenates the organic substrate of the FMO. Please note only the two most common adducts are shown for clarity. Additional adducts are extensively reviewed in [2,6].](/fulltextimages/10737/fig_2.png)
Figure 2: Most Common Flavin-Oxygen Adducts in Flavin Monooxygenases Flavin reduction occurs in one of the three general ways illustrated in Figure 3. The reduced flavin then reacts with molecular oxygen to generate a highly reactive adduct that monooxygenates the organic substrate of the FMO. Please note only the two most common adducts are shown for clarity. Additional adducts are extensively reviewed in [2, 6].
This review article starts with a description of the three general strategies employed by FMOs to reduce the flavin cofactor for the subsequent controlled reaction with oxygen and the oxidation of an organic substrate. We then describe the eight classes of FMO by noting their general protein fold, electron source and the canonical enzyme of each group. If the best characterized enzyme of a group has limited medical relevance we also note a biomedically related example. Our listing is far from all inclusive, but instead is intended to highlight the most pertinent mechanistic or biophysical references for each FMO group for a quick reference source for readers who may encounter an FMO in their research and wish to explore it further to describe disease etiology or treatment. For quick an easy reference examples discussed in the article are provide in Table 1.
| Group | Protein Fold | Cofactor | Electron Donor | Canonical Member | Biomed Rep |
|---|---|---|---|---|---|
| A | Rossmann | FAD | NAD(P)H | PHBHa | rifampicin MO |
| B | Rossmann | FAD | NAD(P)H | SidA | SidA |
| C | TIM barrel | FMN | FMNH2 | luciferase | luciferase |
| D | Acyl CoA-dehydrogenase | FAD/FM N | FADH2/FMNH2 | HpaBb | isobutylamine MO |
| E | Rossmann | FAD | FADH2 | styrene MO | squalene MO |
| F | Rossmann | FAD | FADH2 | PrnAc | KyzR |
| G | Rossmann | FAD | substrate | tryptophan 2-MO | none |
| H | TIM barrel | FMN | substrate | lactate 2-MO | nitronate MO |
Strategies for Flavin Reduction and Substrate Monooxygenation
Before presenting the accepted grouping of FMOs established in 2014 [5], it is useful to consider a broader classification based on how the isoalloxazine ring of the flavin is reduced in the reductive half reaction and the organic substrate is monooxygenated in the oxidative half reaction. This is schematically illustrated in Figure 3. The earliest and most extensively studied type of FMOs catalyze both half reactions within the same protein that is encoded by a single gene. These are referred to as single component FMOs, form C4a-oxygen adducts in their oxidative half reactions and are highlighted in blue in the figure. Single component FMOs are reduced by NAD(P)H which either acts as a substrate (Group A) or remains tightly bound throughout the catalytic cycle acting as an effector molecule (Group B) [8].
![Figure 3: The earliest and most extensively studied type of FMOs catalyze both half reactions within the same protein that is encoded by a single gene. These are referred to as single component FMOs, form C4a-oxygen adducts in their oxidative half reactions and are highlighted in blue in the figure. Single component FMOs are reduced by NAD(P)H which either acts as a substrate (Group A) or remains tightly bound throughout the catalytic cycle acting as an effector molecule (Group B) [8].](/fulltextimages/10737/fig_3.png)
Figure 3: General Types of Flavin Monooxygenases (FMOs) These broad groupings are based on how the flavin cofactor is reduced and electrons are transferred to the substrate for oxidation. Single component FMOs use a nicotinamide coenzyme for flavin reduction. Two component FMOs use a separate protein to reduce the flavin which is then delivered to a sperate monooxygenase as a substrate. Internal FMOs use the organic substrate as the source of electrons for flavin reduction. The flavin then activates oxygen for substrate monooxygenation that occurs all within the same active site.
Another common type of enzyme which comprise the majority of groups of FMOs are the two component monooxygeases. These systems involve two proteins to carry out the overall reaction. The first component contains a NAD(P)H dependent reductase that transfers a hydride ion from the nicotinamide (most typically) to the flavin. Reduced flavin is then transferred to the monooxygenase component as a substrate. Remarkably, this is done through a diffusive process between the reductase and monooxygenase components without an unproductive oxidization of the reduced flavin by oxygen in solution [12, 13]. The organic substrate is then converted to product in the monooxygenase component using the reactions described above (mainly through formation of flavin C4a- oxygen adducts). Finally, internal FMOs represent an entirely self-sufficient class of enzymes in which a protein bound flavin is reduced by electrons from an organic substrate. The same reduced flavin molecule then activates molecular oxygen for the monooxygenation reaction to form the final product. All of these processes occur within the same enzyme molecule.
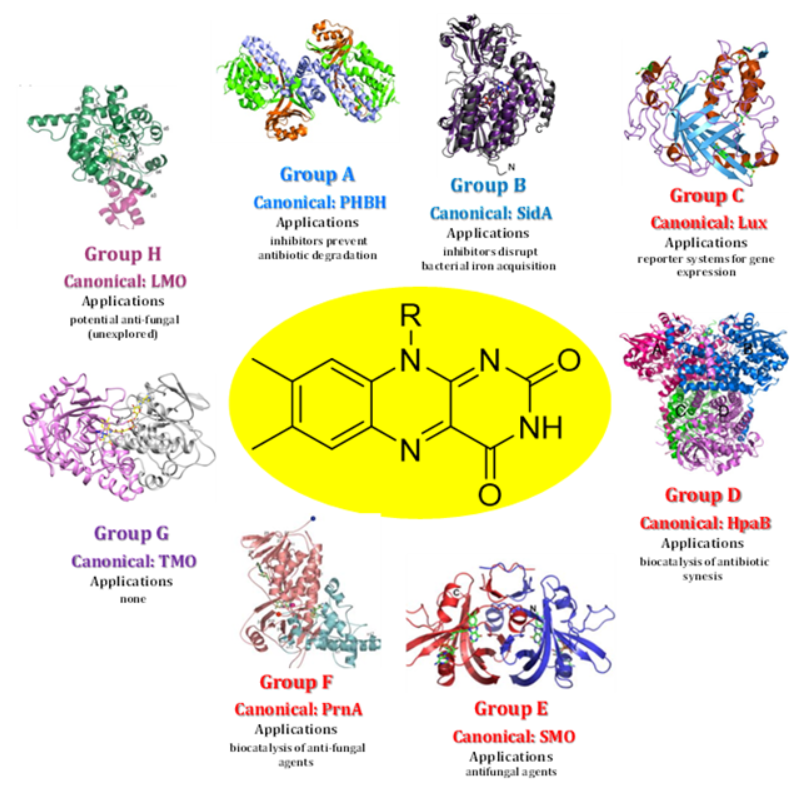
Figure 4: Structure and Application of Each Flavin Monooxygenase Group Structures shown are for the most studied enzymes of each group which is not necessarily the one most extensively used for biomedical applications. Those can be found Table 1 and the main text. Coloring of group names correspond to Figure 3 showing the general FMO types. Abbreviations and PDB IDs are as follows: a) PHBH: p-hydroxybenzoate hydrolase (PDB ID: 1pbe b) SidA: siderophore A (PDB ID: 4B63) c) Lux: bacterial luciferase (PDB ID: 1luc) d) HpaB: p- hydroxyphenylacetate 3-hydroxylase (PDB ID: 2jbt) e) SMO: styrene monooxygenase (PDB ID: 3ihm) f) PrnA: tryptophan 7-halognease (PDB ID: 2aqi) g) TMO: tryptophan-2-monoooxygenase (PDB ID:4iv9) h) LMO: lactae-2- monooxygenase (PDB ID: 6DVI)
One Cofactor Eight Groups Many Biomedical Applications: Overview of FMO Grouping with Biomedical Examples
In 2014, van Berkel and coworkers updated and greatly expanded their previously accepted classification system for FMOs [5] first described in 2006 [3]. The system is based largely on structural homology between group members especially in relation to the flavin binding domain(s). It is also based the electron source used to reduce the flavin cofactor and which of the three strategies describe above utilized to carry out substrate monooxygenation. Canonical enzyme members have emerged for each group typically due to the ease of expression and purification, the availability of crystal structure and convenient kinetic assays to characterize the catalytic mechanism. The sections below will provide an overview of each FMO group. If the best studied enzyme of a group has little or no biomedical relevance, one will be briefly mentioned. Importantly, both review articles and extensive references are provided for the reader to avoid the requirement for an exhaustive search of the extensive body of literature that exists for most FMO groups. Both Table 1 and Figure 4 summarized the characteristics of each FMO group and the examples provided in review.
Group A Flavin Monooxygenases: Canonical Enzyme p-Hydroxybenzoate Hydroxybenzoate Hydroxylase; Biomedical Representative Rifampicin Monooxygenase (Antibiotic Degradation by Pathogens)
Group A is the largest and most extensively characterized class of FMOs with over 90 members isolated and at least partially characterized [2]. They all contain a glutathione reductase-2 (GR-2) type Rossmann fold [14] for FAD binding. These folds, also present in Groups B and E through G, contains a three-stranded antiparallel β-sheet with a central five-stranded parallel β-sheet surrounded by two α-helices. This motif is arranged in such asway that many hydrogen bonding interactions are formed with the adenosine moiety of FAD. Hydrogen bonding interactions are further optimized by the elongated confirmation FAD adopts in the GR-2 Rossmann fold. Electrons for flavin reduction by Group A FMOs are provided NAD(P)H that does not remain bound throughout the catalytic cycle but instead is released after the hydride ion is transferred. The organic substrate, most often substituted aromatic compounds [2, 5, 6, 11], act as effectors to stimulate both flavin reduction and NAD(P) H release as best studied in the canonical Group A member p-hydroxybenzoate hydroxylase (E.C. 1.14.13.2) [15, 16]. Group A members are single component FMOs that form C4(a)-(hydro)peroxyflavins for substrate monooxygenation. Many microbes use Group A FMOs to both harvest carbon from aromatic compounds and to derive energy from these molecules [6]. Additionally, they are used by microbial pathogens to degrade aromatic antibiotics such the reactions catalyzed by tetracycline monooxygenase (E.C. 1.14.13.231) [17] and rifampicin monooxygenase (E.C. 1.14.13.231; Scheme 1) [18].
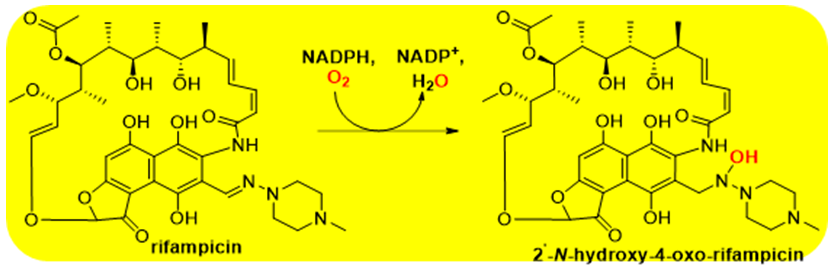
Scheme 1: The reaction catalyzed by rifampicin monooxygenase This Group A FMO is one of many that degrades antibiotics and is expressed by pathogenic microbes for protection.
Group B Flavin Monooxygenases: Canonical Member Phenylacetone Monooxygenase; Biomedical Representative SidA (Iron Acquisition in Pathogenic Bacteria for SidA; Biocatalysis of Antibiotics By Other members)
Around 30 Group B FMOs have been characterized, though many more genes have been annotated as putative members [2]. These enzymes also contain a GR-2 Rossman as described above to bind FAD, but they also have an additional α/β Rossman like domain for NAD(P)H binding [5]. This extra domain is needed due the slight differences in the mechanism employed by Group B FMOs. While they are also single component enzymes, FAD is reduced by NAD(P)H prior to the binding of the substrate to be monooxygenated. Additionally, the pyridine nucleotide remains tightly bound during the oxidative half reaction to modulate the reactivity of the reduced flavin cofactor to form the C4a-oxygen adduct for substrate oxidation. Group B FMOs are subdivide into four sequence related subgroups that relate to the type of oxidations they carry out. These contain Baeyer- Villiger monooxygenases [19], human and plant flavin monooxygenases [4], N-hydroxyl monooxygenases [20] and YUCCAAs [21]. The first structure of a Group B FMO was solved for phenylacetone monooxygenase (E.C. 1.14.13.92) in 2004 [22]. Group B FMOs have been used for the bio- catalysis of antibiotics and are involved in the hydroxylation of amino acids to produce hormones. The metabolic pathway for bacterial siderophore biosynthesis involves the Group B monooxygenase SidA (E.C. 1.14.13.195) [8, 23]. Since siderophores are critical for iron acquisition and bacterial growth they represent an important drug target.
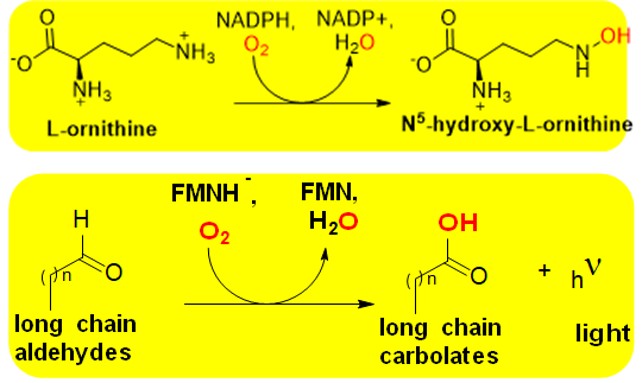
Scheme 2: The reaction catalyzed by SidA This Group B FMO is involved in the anabolic pathway to produce siderophores in pathogenic bacteria. Siderophores are critical for iron acquisition, so targeting SidA to abolish their formation could provide effective treatment of bacterial infections.
Group C Flavin Monooxygenases: Canonical and Biomedical Representative Enzyme Luciferase (Bioluminescent Properties Allows for Use as Reporter of Gene Expression in Study of Many Diseases)
There are currently ~20 known Group C FMOs and an extraordinary capacity of the most famous member, luciferase (E.C. 1.13.12.7), for bioluminescence has led to their extensive study [2, 3, 5, 6]. Like other two component enzymes, Class C FMOs are not able to independently reduce their FMN cofactors but instead contain a NADH dependent reductases (Fre or Frp [24]) to supply electrons for the monooxygenase reaction. Reduced flavin (FMNH2) is delivered to the monooxygenase component as a substrate. Class C FMOs have wildly different folds than other FMOs. Instead of Rossmann folds the monooxygenase component contains a (α/β)8 TIM-barrel [25] also utilized by Group H members. The best characterized Group C FMO, as well as one of the extensively applied of all types of flavoenzymes is luciferase (Scheme 3). This FMO was first described in 1940s [26] and is the only flavoprotein known that emits light during its catalytic catalysis (lmax 450-500 nm). Luciferases from a wide range of organisms have been used to construct reporter systems for gene expression [27]. Many of these reporter systems have been commercialized and used in the study of various of diseases [27]. The precise mechanism of bacterial luciferase it not fully established, but has been studied with long chain aldehydes and shown to involve the formation of a C4a- peroxy or hydroperoxy oxygen adduct [2, 6]. Other Group C member do not undergo bioluminescence, but instead have been used as biocatalysts to produce a wide variety of compounds with important biotechnological applications [2].
Scheme 3: The reaction catalyzed by Luciferase This Group C FMO has the unusual capacity for bioluminescence. This intriguing property has led to many theoretical studies and now common use of the enzyme as a reporter system for gene expression. This has wide ranging applications in the study of disease etiology and drug interactions.
Group D Flavin Monooxygenases: Canonical Enzyme 4-Hydroxyphenylacetate 3-Hydroxylase; Biomedical Representative Isobutylamine N1-Monooxygenase (Biocatalysis of Valinomycin for Cancer Studies)
There are currently more than 40 characterized Group D members all of which are two component systems comprising of a monooxygenase with an acyl CoA dehydrogenase type fold [2]. These folds have three distinct components. Both the N’- and C’-termini have α-helical domains and these are separated by a third domain consisting of β-sheets [28]. Group D FMOs are the only class that have a dual cofactor specify with some utilizing FMN and others FAD [2, 5]. The different types of Group D FMOs reflect ways in which the flavin cofactor and the organic substrate bind to the enzymes. FMN dependent members utilize large flavin reductase for cofactor delivery to the monooxygenase [29]. FAD dependent members, on the other hand, have relatively smaller reductase components to deliver FADH2 to the monooxygenase [30]. Different cofactor specificity does not represent alternate mechanisms of monooxygenation, but rather is the result of evolutionary convergence [5]. The mechanism of Group D members involves electrophilic attack of the substrate by a reactive C4a-hydroperoxyflavin adduct (Figure 2).
Substrate specificities of Group D FMOs are very similar to Group A enzymes in that they hydroxylate aromatic and heterocyclic rings. The prototypical Group D FMO is 4-hydroxyphenyl acetate 3-hydoxylase (E.C.1.14.14.9 [31, 32]. Biomedical applications of Group D members include the N-hydroxylation reaction catalyzed by isobutylamine N1-monooxygenasese (E.C. 1.14.14.30) from Streptomyces sp. This enzyme catalyzes the rate controlling step in biosynthesis of the antibiotic valinomycin (Scheme 4) [33]. Though not approved for clinical use in the United States, it has shown antiviral activity [34]. It is also used experimentally in studies of cancer biology since it induces apoptosis is. Biocatalytic studies have focused on isobutylamine N1-monooxygenasese in order to optimize commercial production of valinomycin [2].
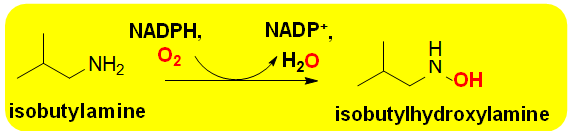
Scheme 4: The reaction catalyzed by Luciferase This Group C FMO has the unusual capacity for bioluminescence. This intriguing property has led to many theoretical studies and now common use of the enzyme as a reporter system for gene expression. This is wide ranging applications in the study of disease etiology and drug interactions.
Group E Flavin Monooxygenases: Canonical Enzyme Styrene Monooxygenase; Biomedical Representative Squalene Monooxygenase (Targating Cholesterol Anabolism for Treatment of Fungal Infections)
More than 20 Group E FMOs have been isolated and at least partially purified and characterized [35]. All contain Rossmann folds as decribed for Group A enzymes above and use FAD as cofactor [2, 6]. These enzymes carry out the regio- and stero- selective reactions to produce either epoxides or sulfoxides [2]. They use a FAD dependenpt reductase to reduce the flavin and deliver it to the minoxygenase component. This component reacts with molecular oxygen to generation a C4a-hydroperoxy adduct for monooxygenation. Unlike the case of other two component FMOs, monooxygention is strongly promoted through interactions of the redcctase with the monooxyganse [36, 37]. Brodly speaking, Group E enzymes are sudivided into two groups that act either styrene derivtives or unsubstituded indoles [2, 5]. Catalyzed reactions occur on substitued alkenes and have been extensively studied for useful biotechnilogical applications [2, 35]. The caniconical enzyme of Group E is styrene monooxygenase (E.C. 1.14.14.11) [38, 39]. The Group E enzyme squalene monooxugenase (E.C. 1.14.14.17) is an imprtanat drug target for the tereatment of fungal infections [40]. The enzyme is targeted by terbinfine (brand name Lamicil®) for the topical treatment of athletes foot.
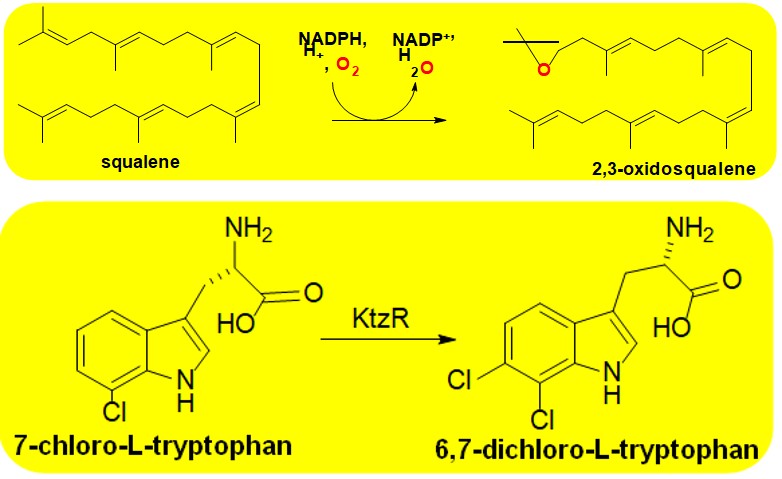
Scheme 5: The reaction catalyzed by Squalene Monooxygenase This Group E FMO catalyzes the second rate- controlling step of cholesterol biosynthesis. It is the target of the topical anti-fungal terbinafine (brand name Lamisil®). Inhibition of the FMO prevents cholesterol synthesis in fungi weakening their cell membrane to make them susceptible to the innate immune system.
Group F Flavin Monooxygenases: Canonical Enzyme Tryptophan 7-Halogenase; Biomedical Representative KtzR (Anti-fungal Target; Other Member Widely Used as Biocatalysis in the Pharmaceutical Industry)
Around 40 Group F FMOs have been described each of which contain a GR-2 Rossmann folds to bind a FAD cofactor used in the reaction [2, 6]. These two component systems are halogenases with most catalyzing chlorination of aromatic heterocyclic rings. However, brominating and even iodinating enzymes such as VirX1 are known [41]. Group F members have two fingerprint motifs in their monooxygenase component. One is a GxGxxG region (x is any amino acid) and the other is a so called “W-box fingerprint” that contains the sequence WxWxIP [42]. The latter sequence plays an important role in controlling the strict regioselectivity of the catalyzed halogenation. The regioselectivity of Group F FMOs make them widely used in chemical industries for the generation of natural products and antibiotics including chloramphenicol [2].
Group F enzymes have been most extensively studied in the context of the chlorination of tryptophan [2, 5, 6]. The canonical Group F member is tryptophan-7-monooxygenase (PrnA; E.C.1.14.19.9). PrnA has been extensively characterize both mechanistically and in terms of how information is communicated between the reductive and oxidative units of the two component FMO [42]. The catalytic mechanism of Group F FMOs involves the electrophilic aromatic substitution reaction of the organic substrate with an inorganic halo acid induced by the C4a-hydroperoxy form of FAD (Figure 2) [2]. Communication of between active sites of each of the two components is achieved largely through conformational changes of the isoalloxazine ring of the flavin induced by conserved lysine and glutamate residues [2, 42]. Such functional role of flavin dynamics are reminiscent of the single component Group A enzyme PHBH [6, 8]. As mentioned, Group F enzymes are widely used in the biosynthesis of antibiotics so have important biomedical applications. A notable example is KtzR (E.C. 1.14.19.60) which is involved in the synthesis of the antifungal kutzneride [43]. The enzyme, whose reaction is shown in Scheme 6, is one of several tryptophan halogenates involved in the production of the antifungal agent.
Scheme 6: The reaction catalyzed by KtzR This Group F FMO catalyzes the halogenation of amino acids in microbes which is an important step in antibiotic biosynthesis. This property of group F FMOs have made them extremely useful as biocatalysts in the pharmaceutical industry to produce an array of compounds of biomedical import.
Group G Flavin Monooxygenases: Canonical Enzyme Tryptophan 2-Monooxygenase; No Biomedical Representatives
Group G is small and currently contains only four members all of which contain monoamine oxidase type fold [44] to bind FAD for the oxidative decarboxylation of amino acids [2, 5, 6]. The enzymes are involved in amino acid metabolism in vivo and consist of trptophan-2- monooxygenase (Scheme 7; E.C. 1.13.12.3) [45], arginine monooxygenase (E.C. 1.13.12.1) [2], lysine monooxygenase (E.C. 1.13.12.2) [46] and phenylalanine 2- monooxygenase (E.C. 1.13.12.9) [47]. Group G members are internal monooxygenases containing a single component capable of taking electrons from a substrate to reduce the FAD cofactor. FAD remains bound to the enzyme to activate molecular oxygen for amino acid monooxygenation.
Group G FMOs utilize a two-step catalytic mechanism in which the amino acid is oxidized through a hydride ion transfer reaction from the α-carbon to the flavin N(5)-position of the isoalloxazine ring (Figure 1) in a similar manner as the flavin oxidases [6] to generate FADH2. Evidence exists for two sperate types of mechanisms for oxygenation of the imino acid intermediate that from the hydride transfer step. One route involves the formation of a C4a-hydroperoxoflavin (Figure 2) that undergoes an electrophilic attack of the α-carbon of the imino acid to eliminate carbon dioxide. This would be followed by tautomerization to form the final amide product. In the other proposed mechanism of oxygenation, the flavin C4a-hydroperoxide is not stabilized but instead the hydrogen peroxide formed upon re-oxidation of the flavin reacts with the imino acid to generate the final amide product. It remains unclear which mechanism is operative or if both occur depending on the enzyme and specific reaction conditions [2, 6].
Regardless of the mechanistic details Group G FMOs have little biomedical relevance since they are mainly involved in gall formation and other aspects of plant physiology. They have found moderate use in chemical industries, but their narrow substrate specificities limit their applications. Kinetic studies of these enzymes has revealed unusual behaviors resulting from the branching of reaction intermediates that could indirectly benefit mechanistic studies of biomedically important systems [48].
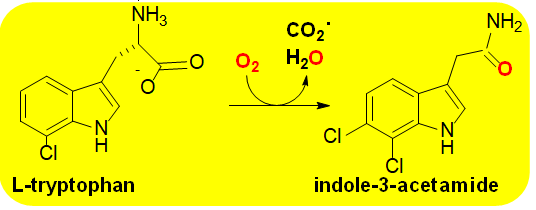
Scheme 7: The reaction catalyzed by tryptophan monooxygenase Group G FMOs catalyzes the oxidative decarboxylation of amino acids. Activity is shown to be correlated with gall formation in plants. While Group G members do not have known biomedical relevance mechanistic studies have reveled important kinetic consequences that could have indirect applications to biomedical enzymes.
Group H Flavin Monooxygenases: Canonical Enzyme Lactate 2-Monooxygenase; Biomedical Representative Nitronate Monooxygenase (Potential Anti-fungal)
Group H FMOs are another small category of enzymes that currently only contain four members [2, 5, 6]. They are similar to Group G members in the sense that they are self-sufficient internal monooxygenases, that catalyze decarboxylation reactions. They have different over folds though, are also known to catalyze denitrification reactions and utilize different catalytic mechanisms for monooxygenation. Group H FMOs contain (β/α)8 TIM barrel folds to bind FMN cofactors [25]. These enzymes display an unusual features among flavoproteins lacking metal ions in that they undergo radical mechanisms involving flavosemiquinone intermediates. This has been most extensively studied in nitronate monooxygenase (Scheme 8; E.C. 1.13.12.16) where an anionic flavosemiquinone was observed and well characterized through both anaerobic substrate reductions and rapid kinetic measurements [49, 50]. In this mechanism, a single electron is transferred from the substrate to the flavin N5-postion of the isoalloxazine ring (Figure 1) to generate an enzyme bound substrate radical. Molecular oxygen then reacts with the flavin radical to re-oxidize the cofactor. The resulting superoxide radical then rapidly reacts with the substrate radical to produce a peroxide that is released from the active site and undergoes a non-enzymatic decomposition to produce the final carbonyl product. The canonical Group H enzyme is lactate 2-monooxygenase that was first identified by Edson in 1947 [1] and whose structure was solved in 2018 [51]. Nitronate monooxygenase whose structure is also known has modest biomedical relevance given that the preferred substrate, propionate-3- nitonate, is a potent and irreversible inhibitor of succinate dehydrogenase, an essential metabolic enzyme for aerobic organisms [49]. Over 2,000 genes encoding putative nitronate monooxygenases have been identified including in many pathogens [2]. Studies have shown that the enzyme is expressed in Mycobacterium sp. under hypoxic conditions [49]. However, to date no studies have been carried out to target the enzyme for treatment of disease.
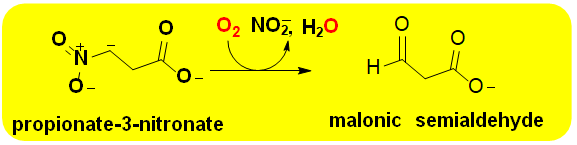
Scheme 8: The reaction catalyzed by nitronate monooxygenase This Group H FMO catalyzes the oxidative denitrification alkyl nitronates. Propionate-3-nitronate, the physiological substrate of nitronate monooxygenase, is a potent irreversible inhibitor of succinate dehydrogenase. Both plants and symbiotic fungi produce the FMO for protection.
Conclusion and Summary
As outlined here the chemical versatility of the flavin cofactor gives rise to a plethora of reactions catalyzed by FMOs many of which have important applications in medicine and disease. FMOs are grouped based on their overall protein fold, source of electrons to reduce the flavin cofactor and strategy to carry out the overall reaction. Groups A and B FMOs carry out their entire catalytic cycles within the same protein and are thus referred to as single component FMOs. Groups C-F are two component systems in which a NAD(P)H dependent reductase first transfers electrons to the flavin via a hydride ion and then transfers the reduced molecule to the monooxygenase component for substrate oxidation. Finally, internal monooxygenases carry out their entire reaction solely with an enzyme bound flavin. They take electrons from an organic substrate to reduce the flavin and then react with another substrate to produce the final product of the enzymatic reaction. With over 250 FMOs described to date and thousands of research articles it is easy to get lost in the pile of data collected for these enzymes. We hope this review lays a solid foundation for more detailed studies of the highly recommend and comprehensive reviews that are noted [2, 4, 6, 7, 8]. The readings should aid in the study of new FMOs for the treatment of disease.
Acknowledgements
This work was supported by a Departmental Research Grant from The Robert A. Welch Foundation AC-0006.
References
-
Edson NL (1947) The oxidation of lactic acid by Mycobacterium phlei. Biochem J 41(2): 145-151.
-
Paul CE, Eggerichs D, Westphal AH, Tischler D, van Berkel WJH (2021) Flavoprotein monooxygenases: Versatile biocatalysts. Biotechnol Adv 51: 107712.
-
van Berkel WJH, Kamerbeek NM, Fraaije MW (2006) Flavoprotein monooxygenases, a diverse class of oxidative biocatalysts. J Biotechnol 124(4): 670-689.
-
Deng Y, Zhou Q, Wu Y, Chen X, Zhong F (2022) Properties and Mechanisms of Flavin-Dependent Monooxygenases and their Applications in Natural Product Synthesis. Int J Mol Sci 23(5): 2622.
-
Huijbers MME, Montersino S, Westphal AH, Tischler D, van Berkel WJH (2014) Flavin dependent monooxygenases. Archives of Biochemistry and Biophysics 544: 2-17.
-
Romero E, Gómez Castellanos JR, Gadda G, Fraaije MW, Mattevi A (2018) Same Substrate, Many Reactions: Oxygen Activation in Flavoenzymes. Chemical Reviews 118(4): 1742-1769.
-
Massey V (1994) Activation of molecular oxygen by flavins and flavoproteins. J Biol Chem 269(36): 22459- 22462.
-
Reis RAG, Li H, Johnson M, Sobrado P (2021) New frontiers in flavin-dependent monooxygenases. Archives of Biochemistry and Biophysics 699: 108765.
-
McDonald AG, Tipton KF (2023) Enzyme nomenclature and classification: the state of the art. Febs J 290(9): 2214-2231.
-
(2016) International Man of Enzymology: Vince Massey’s Work on Flavin Catalysis. J Biol Chem 291(29): 15404-15405.
-
Teufel R, Miyanaga A, Michaudel Q, Stull F, Louie G, et al. (2013) Flavin- mediated dual oxidation controls an enzymatic Favorskii-type rearrangement. Nature 503: 552-556.
-
Sucharitakul J, Tinikul R, Chaiyen P (2014) Mechanisms of reduced flavin transfer in the two-component flavin- dependent monooxygenases. Arch Biochem Biophys 555-556: 33-46.
-
Ellis HR (2010) The FMN-dependent two-component monooxygenase systems. Arch Biochem Biophys 497(1- 2): 1-12.
-
Dym O, Eisenberg D (2001) Eisenberg, Sequence- structure analysis of FAD-containing proteins. Protein Sci 10(9): 1712-1728.
-
Entsch B, Cole LJ, Ballou DP (2005) Protein dynamics and electrostatics in the function of p-hydroxybenzoate hydroxylase. Arch Biochem Biophys 433(1): 297-311.
-
Entsch B, van Berkel WJ (1995) Structure and mechanism of para-hydroxybenzoate hydroxylase. Faseb J 9(7): 476- 483.
-
Bernal J, Ojutiku Z, Cordero A, Martinez A, Berkey K, et al. (2020) An Improved Protocol for the Expression and Purification of Tetracycline Monoozyhenase: An Enzyme Involved in Antibiotic Resistance. Annals of Advanced Biomedical Science 3(2): 1-7.
-
Liu LK, Abdelwahab H, Martin Del Campo JS, Mehra- Chaudhary R, Sobrado P, et al. (2016) The Structure of the Antibiotic Deactivating, N-hydroxylating Rifampicin Monooxygenase. J Biol Chem 291(41): 21553-21562.
-
Fürst MJLJ, Gran-Scheuch A, Aalbers FS, Fraaije MW (2019) Baeyer–Villiger Monooxygenases: Tunable Oxidative Biocatalysts. ACS Catalysis 9(12): 11207- 11241.
-
Ernst S, Mährlein A, Ritzmann NH, Drees SL, Fetzner S (2022) A comparative study of N-hydroxylating flavoprotein monooxygenases reveals differences in kinetics and cofactor binding. Febs J 289(18): 5637- 5655.
-
Ballou DP, Entsch B (2013) 1 The reaction mechanisms of Groups A and B flavoprotein monooxygenases. In: Russ H, et al. (Eds.), Complex Flavoproteins, Dehydrogenases and Physical Methods, De Gruyter, Berlin, Boston, 2: 1-28.
-
Malito E, Alfieri A, Fraaije MW, Mattevi A (2004) Crystal structure of a Baeyer-Villiger monooxygenase. Proc Natl Acad Sci U S A 101(36): 13157-13162.
-
Frederick RE, Ojha S, Lamb A, Dubois JL (2014) How pH modulates the reactivity and selectivity of a siderophore- associated flavin monooxygenase. Biochemistry 53(12): 2007-2016.
-
Campbell ZT, Baldwin TO (2009) Fre is the Major Flavin Reductase Supporting Bioluminescence from Vibrio harveyi Luciferase in Escherichia coli. J Biol Chem 284(13): 8322-8328.
-
Gromiha MM, Pujadas G, Magyar C, Selvaraj S, Simon I (2004) Locating the stabilizing residues in (alpha/ beta)8 barrel proteins based on hydrophobicity, long- range interactions, and sequence conservation, Proteins 55(2): 316-329.
-
Hastings JW (1978) Bacterial bioluminescence light emission in the mixed function oxidation of reduced flavin and fatty aldehyde. CRC Crit Rev Biochem 5(2): 163-184.
-
Phonbuppha J, Tinikul R, Wongnate T, Intasian P, Hollmann F, et al. (2020) A Minimized Chemoenzymatic Cascade for Bacterial Luciferase in Bioreporter Applications. Chembiochem 21(14): 2073-2079.
-
Battaile KP, Molin-Case J, Paschke R, Wang M, Bennett D, et al. (2002) Crystal structure of rat short chain acyl- CoA dehydrogenase complexed with acetoacetyl-CoA: comparison with other acyl-CoA dehydrogenases. J Biol Chem 277(14): 12200-122007.
-
Sucharitakul J, Chaiyen P, Entsch B, Ballou DP (2006) Kinetic Mechanisms of the Oxygenase from a Two- component Enzyme, p-Hydroxyphenylacetate 3-Hydroxylase from Acinetobacter baumannii. J Bio Chem 281(25): 17044-17053.
-
Chakraborty S, Ortiz-Maldonado M, Entsch B, Ballou DP (2010) Studies on the Mechanism of p- Hydroxyphenylacetate 3-Hydroxylase from Pseudomonas aeruginosa: A System Composed of a Small Flavin Reductase and a Large Flavin-Dependent Oxygenase. Biochemistry 49(2): 372-385.
-
Chenprakhon P, Dhammaraj T, Chantiwas R, Chaiyen P (2017) Hydroxylation of 4-hydroxyphenylethylamine derivatives by R263 variants of the oxygenase component of p-hydroxyphenylacetate-3-hydroxylase. Arch Biochem Biophys 620: 1-11.
-
Deng Y, Faivre B, Back O, Lombard M, Pecqueur L, et al. (2020) Structural and Functional Characterization of 4-Hydroxyphenylacetate 3-Hydroxylase from Escherichia coli. Chembiochem 21(1-2): 163-170.
-
Parry RJ, Li W (1997) An NADPH:FAD oxidoreductase from the valanimycin producer, Streptomyces viridifaciens. Cloning, analysis, and overexpression. J Biol Chem 272(37): 23303-23311.
-
Zhang D, Ma Z, Chen H, Lu Y, Chen X (2020) Valinomycin as a potential antiviral agent against coronaviruses: A review. Biomed J 43(5): 414-423.
-
Heine T, Scholtissek A, Hofmann S, Koch R, Tischler D (2020) Accessing Enantiopure Epoxides and Sulfoxides: Related Flavin-Dependent Monooxygenases Provide Reversed Enantioselectivity ChemCatChem 12(1): 199- 209.
-
Morrison E, Kantz A, Gassner GT, Sazinsky MH (2013) Structure and Mechanism of Styrene Monooxygenase Reductase: New Insight into the FAD-Transfer Reaction. Biochemistry 52(35): 6063-6075.
-
Tischler D, Kermer R, Groning JA, Kaschabek SR, Van Berkel WJ, et al. (2010) StyA1 and StyA2B from Rhodococcus opacus 1CP: a multifunctional styrene monooxygenase system. J Bacteriol 192(19): 5220-5227.
-
Gassner GT (2019) The styrene monooxygenase system. Methods Enzymol 620: 423-453.
-
Kantz A, Gassner GT (2011) Nature of the Reaction Intermediates in the Flavin Adenine Dinucleotide- Dependent Epoxidation Mechanism of Styrene Monooxygenase. Biochemistry 50(4): 523-532.
-
Padyana AK, Gross S, Jin L, Cianchetta G, Narayanaswamy R, et al. (2019) Structure and inhibition mechanism of the catalytic domain of human squalene epoxidase. Nat Commun 10(1): 97.
-
Gkotsi DS, Ludewig H, Sharma SV, Connolly JA, Dhaliwal J, et al. (2019) A marine viral halogenase that iodinates diverse substrates. Nat Chemi 11(12): 1091-1097.
-
Zhu X, De Laurentis W, Leang K, Herrmann J, Ihlefeld K, et al. (2009) Structural Insights into Regioselectivity in the Enzymatic Chlorination of Tryptophan. J Mol Bio 391(1): 74-85.
-
Heemstra JR, Walsh CT (2008) Tandem Action of the O2- and FADH2-Dependent Halogenases KtzQ and KtzR Produce 6,7-Dichlorotryptophan for Kutzneride Assembly. J Am Chem Soc 130(43): 14024-14025.
-
De Colibus L, Li M, Binda C, Lustig A, Edmondson DE (2005) Three-dimensional structure of human monoamine oxidase A (MAO A): relation to the structures of rat MAO A and human MAO B. Proc Natl Acad Sci 102(36): 12684-12689.
-
Gaweska HW, Taylor AB, Hart PJ, Fitzpatrick PF (2013) Structure of the flavoprotein tryptophan 2- monooxygenase, a key enzyme in the formation of galls in plants. Biochemistry 52(15): 2620-2626.
-
Mashiguchi K, Tanaka K, Sakai T, Sugawara S, Kawaide H, et al. (2011) The main auxin biosynthesis pathway in Arabidopsis. Proc Natl Acad Sci 108(45): 18512-18517.
-
Ida K, Suguro M, Suzuki H (2011) High resolution X-ray crystal structures of l-phenylalanine oxidase (deaminating and decarboxylating) from Pseudomonas sp. P-501. Structures of the enzyme-ligand complex and catalytic mechanism. J Biochem 150(6): 659-669.
-
Fitzpatrick PF (2023) PF The aromatic amino acid hydroxylases: Structures, catalysis, and regulation of phenylalanine hydroxylase, tyrosine hydroxylase, and tryptophan hydroxylase. Arch Biochem Biophys 735: 109518.
-
Francis K, Smitherman C, Nishino SF, Spain JC, Gadda G (2013) The biochemistry of the metabolic poison propionate 3-nitronate and its conjugate acid, 3-nitropropionate. IUBMB Life 65(9): 759-768.
-
Gadda G, Francis K (2010) Nitronate monooxygenase, a model for anionic flavin semiquinone intermediates in oxidative catalysis. Arch Biochem Biophys 493(1): 53- 61.
-
Koteva K, Cox G, Kelso JK, Surette MD, Zubyk HL, et al. (2018) Rox, a Rifamycin Resistance Enzyme with an Unprecedented Mechanism of Action. CelL Chem Bio 25(4): 403-412.
- Origin, Evolution, and Functional Impact of Short Insertion- Deletion Variants in Human Genomes: A Review
- Harnessing Molecular Glues for Next-Generation Vaccine, Cancer and Cardiovascular Disease Drug Development: A Comprehensive Review
- Lateral Cervical Epidermal Inclusion Cyst in a Paediatric Patient: A Rare Case Report
- Malarial Plasmodium Falciparum with Hepatitis B and C Virus Infections among Blood Donors in Ife Central Local Government Area, Ile Ife, Osun State, Nigeria
- Withanolides and Withaferin A- What’s next in Ashwagandha Research
- Designing of Dual Pulse Photoacoustic Tomography for Imaging of Drug-Response and Tumor Growth