Metformin Improves Dopamine Release and Motor Performance in C57BL/6N Mice induced by MPTP: Insights of Alternative Medication for Parkinson’s Disease
It is obvious that Levodopa can effectively ameliorate most of the motor symptoms associated with Parkinson’s disease. However, Levodopa-induced dyskinesia (LID) experienced due to its long-term usage poses a great concern over it is dependency. Alternative therapies that can equally improve and/or cure the motor symptoms without alarming side effects are unarguably needed. Recently, Metformin, besides being a key player in the management of Diabetes Mellitus type 2 (T2DM), was also found to have some potentials in the treatment of Parkinson’s disease (PD). In this study, MPTP (1-methyl4-phenyl-1,2,3,6-tetrahydropyridine) were used to create the PD models (an established fact supported by multiple studies). Initially, C57BL/6N mice were randomly divided into 3 groups (Saline, MPTP, MPTP + Metformin) (n=8, each). Later, MPTP + Metformin group was further sub-divided into 3 and 100, 200; 400 mg/kg metformin was administered to each sub-group accordingly. After a series of behavioral assays, the mice were sacrificed and the brain was taken out to measure the DA release in the striatum using carbon-fiber electrode (amperometric recording). In each case, Metformin proves to be decisive and display convincing/satisfactory results depicting its neuroprotective prowess.
Introduction
Parkinson’s disease (PD), being the second most common neurodegenerative disease after Alzheimer’s disease (AD) affects 1-2% (predominantly, 55-60 and above years old) of the population [1]. Clinically, PD is characterized by four cardinal motor symptoms (Tremor, Rigidity, Akinesia/ Bradykinesia, Postural instability, TRAP) Most of which are caused by progressive dopaminergic degeneration in the substantia nigra pars compacta (SNc) [2]. Manifestation of key motor symptoms appears in a subject when roughly 50-60% of the dopaminergic neurons are degenerated, producing 70-80% depletion in the Dopamine (DA) levels in the dorsal striatum as assessed mostly using High-Performance Liquid Chromatography (HPLC) and Carbon-Fiber Electrode [3].
Virtually, 95% of the PD cases were found to be sporadic. PD was believed to be caused by a combination of genetic factors, environmental factors and nervous system aging through oxidative stress, proteasome, calcium homeostasis, dysfunction, immune response, mitochondrial dysfunction, excitatory toxins and apoptosis. Collective insults produced by the aforementioned processes are the degeneration and loss of dopaminergic neurons of the substantia nigra [4]. The precise etiology and mechanisms behind the pathogenesis of the disease is still not vividly understood. MPTP (among the toxins) was for long found to be a good agent (drug) that causes PD (as such used universally in the creation of the disease model). Tanner and colleagues [5] found that the exposure to the two widely used insecticides and pesticides (Rotenone & Paraquat respectively) heavily increases the incidence of PD possibly via the competitive inhibition of Complex I of the electron transport chain (ETC), which subsequently leads to the build-up of Reactive Oxygen Species (ROS) [6]. The overall consequences results in the diminished function of the mitochondria. Aggregation of filamentous protein called α-synuclein (Lewy-bodies) within the inter-cytoplasmic region of neurons was believed to be another potential process that develops PD (even though α-synuclein aggregation is not peculiar to PD). Metformin too, was found to have some neuroprotective role against a pathogenesis developed due to this aggregation by preventing α-synuclein to adopt more compact confirmation [7] again, Tayara and colleagues [8] reported that metformin has an anti-inflammatory role. It inhibits the activation of microglia as measured by different anti-inflammatory markers (OX-6, IKKβ and aginase). More so, results of quantitative Polymerase Chain Reaction (qPCR) unveil that metformin treatment diminishes the expression levels of several pro- and anti-inflammatory cytokines.
MPTP Toxicity
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) is the most common toxin use extensively in the creation of PD models. MPTP administration is relatively simpler and require little expertise compared to 6-hydroxydopamne (6-OHDA) which requires stereotaxic frame and surgery on live animals [9], even though, in 6-OHDA has some edge over MPTP in terms of effectiveness. However, MPTP is more effective in the production of suitable and reproducible lesions of the nigrostriatal dopaminergic pathway following its systemic administration [10]. MPTP, being a highly lipophilic compound, crosses the blood-brain barrier (BBB) and is metabolized into 1-methyl-4-phenylpyridinium (MPP+) where it has been shown to block complex I of the electron transport chain (ETC) and increase ROS, lipid peroxidation, and protein oxidation [10]. MPP+ is a polar molecule unlike its precursor MPTP and as such it cannot freely enter cells, but depends heavily on plasma membrane carriers to gain access to dopaminergic neurons [11].
Rotenone Toxicity
Like MPTP, rotenone can easily cross the BBB due to its lipophilic nature and competitively inhibits the activity of complex-I in the mitochondrial respiratory chain, thereby increasing oxidative stress Gao and colleagues [12, 13] reported that rotenone induced dopaminergic neurodegeneration in animal model by means of microglial activation, causing NADPH oxidase-derived superoxide aggregation. However, recent studies conducted using human cell line microglia reveals that activated microglia produced extracellular ROS predominantly and as such do not contribute directly to neurodegeneration when subjected to chronic, low doses of rotenone [14]. In terms of killing nigrostriatal dopaminergic neurons and formation of lewy bodies, rotenone is more eligible than MPTP & 6-OHDA and Yet, it is the most rarely used PD models due to its non- specificity.
Metformin (C4H11N5)
A conventionally accepted oral antidiabetic drug that lowers hepatic blood glucose production and improves insulin sensitivity (Insulin sensitizers) in the target cells via activation of AMP-activated protein Kinase (AMPK). More recently, metformin establishes itself as one of the potential candidate in the treatment of neurodegenerative disease (most profoundly, PD) in animal models. Bayliss [15] reported that metformin prevents nigrostriatal dopaminergic degeneration independent of AMPK activation. This finding, therefore suggests that the mechanism of action of metformin in neurodegenerative cases is distinct to that involved in its antidiabetic role. Hence, the exact mechanism through which metformin exerts its neuroprotective role remain unknown. Targeting AMPK signaling is indeed a neuroprotective strategy in Parkinson’s disease as insisted by Curry [16]. Yet, no evidence that metformin depends on/mediates AMPK activation before exerting it is neuroprotective function. Additionally, two similar but independent studies uses neuroblastoma dopaminergic cells lines to investigate the role of metformin in PD. Lu, et al. [17] reported that metformin prevents dopaminergic cell death via autophagy and mitochondrial ROS clearance, whereas, Kang, et al. [18] affirms that metformin confers it is neuroprotection via activation of the ATF2/CREB-PGC-1α.
Materials & Methods
Study Design
Mice were randomly grouped in to 3. Group 1 were treated with Saline and served as control. Group 2 and 3 were treated with neurotoxin (MPTP & Rotenone respectively). Being an experiment that involved highly toxic agent, one mice extra was added to each group as back-up. The mice were serially trained for 14 consecutive days to acclimatize and adopt the different motor tests (Figure 1). Metformin, 100 mg/kg (low), 200 mg/kg (mid), 400 mg/kg (high) (dissolved in Saline) was administered orally using a plastic enteral feeding tube. 25mg/kg (dissolved in Saline) of MPTP was administered according to the body weight also for 5-consecutive days systematically (Intraperitoneally). Behavioral assessment was carried-out following the administration of MPTP and Metformin.
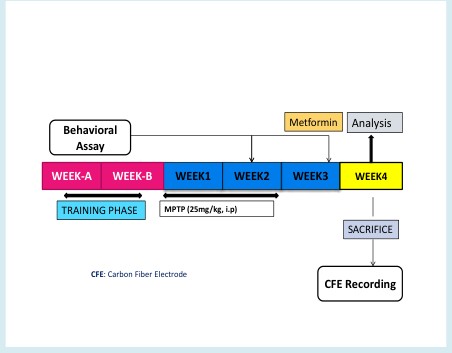
Figure 1: Experimental Design: 100mg/kg (low), 200mg/kg (mid), 400mg/kg (high) body weight metformin was administered orally, twice a day throughout the 7th week. Saline was administered to the respective group since day 1.25mg/kg body weight MPTP was administered for 5-consecutive days (acute system) intraperitoneally. Behavioral assays were carried out in the 2nd and 3rd week. CFE recording was performed after the mice were sacrificed
Animals/Subjects
C57 male mice (25-30 grams) were housed for in the animal centre until the 22nd day (the day they were killed) with a maximum of 5-mice per cage under a 12-hour light/ dark cycle with free access to food and water at a temperature and relative humidity of 22 ± 20C and 60% respectively. At the end of week-3, mice were sacrificed for carbon-fiber electrode (DA release measurement). The anesthetic agent used was isoflurane. The use and care of the animals was approved and put forward by the Animal Care and Use Committee of Xi’an Jiaotong University (XJTU) and the Association for Assessment and Accreditation of Laboratory Animal Care.
MPTP Administration
The discovery of MPTP as a drug that produces PD-like pathology comes accidentally in 1980 following it is abuse among American youth. Such incidence pave way to deeper understanding of the pathogenesis associated with PD. In this study, we adopted the protocol outlined and develop by Tatton & Kish [20] (with little modification), which involves injecting 25 mg/kg free-base MPTP intraperitoneally for 5-consecutive days starting from day 1. This regimen has a great potential of causing apoptosis and depletion of striatal dopamine by 40-50% in young adult C57/BL mice. We avoided using 10-week old mice (in contrast to conventional trends) as we believed 12-week old mice produce the best PD models. Mice lighter than 22 grams were equally avoided. Dopaminergic lesions stabilize fully 3-weeks after MPTP administration. Behavioral changes manifest early and mice were able to produce canonical symptoms similar to that of classical/typical PD, whereas PD-like pathology (DA release & morphology) appears fully in 3rd week. MPTP mice were carefully isolated from the saline group. The excretion of un- metabolized MPTP occurs mainly during the first day after injection and as such under no circumstances MPTP-treated mice should be mixed-up with mice from other groups.
Behavioral Assay
Open Field Test: In our animal center settings, open field apparatus consist of rectangular plastic arena measured 40 X 40 X 50 cm3. The area is well tight to prevent fluid absorption. The whole arrangement is connected with video system in which the area is divided into small squares of 5 cm by 5 cm. The trajectory path (Figure 2a) of the mice was traced/ recorded within the center and corner zones respectively using an automatic tracking system. The open field maze was cleaned thoroughly after each experiment with 70% ethyl alcohol to get rid of odor signal. The mice were taken from their respective cages to the behavior room directly and tested once at a time for 30 minutes. Sometimes a cylindrical
| Metformin | Analysis |
|---|
| WEEK-A | WEEK-B | WEEK1 | WEEK2 | WEEK3 | WEEK4 |
|---|---|---|---|---|---|
| MPTP (25mg/kg, i.p) |
flask is placed at the center to measure to measure the number of times the mice visited the region (center) and that inarguably dictate their social novelty, which, in essence, gives evaluation on the level of depression and anxiety. Footprints: Mice were trained to move linearly for a distance of approximately 1.5 meters. The forelimb and the hind-limb were painted red and blue respectively. The stride length as Saline MPTP a b $$ \mathrm {范 围} = \mathrm {大 范 围} - \mathrm {小 范 围} $$ $$ = \frac {1}{2} \times 1 0 ^ {3} \mathrm {m} ^ {3} $$ well as the overlap were recorded and analyzed accordingly.
Rotarod: This test is used to evaluate mice forelimb and hindlimb motor balance and Coordination. In the 3rd week mice were placed in a separate compartment on the rod and tested at 5 rpm initially until it reaches 40 rpm. The latency to fall (time on rod) was recorded (Figure 2b-d).
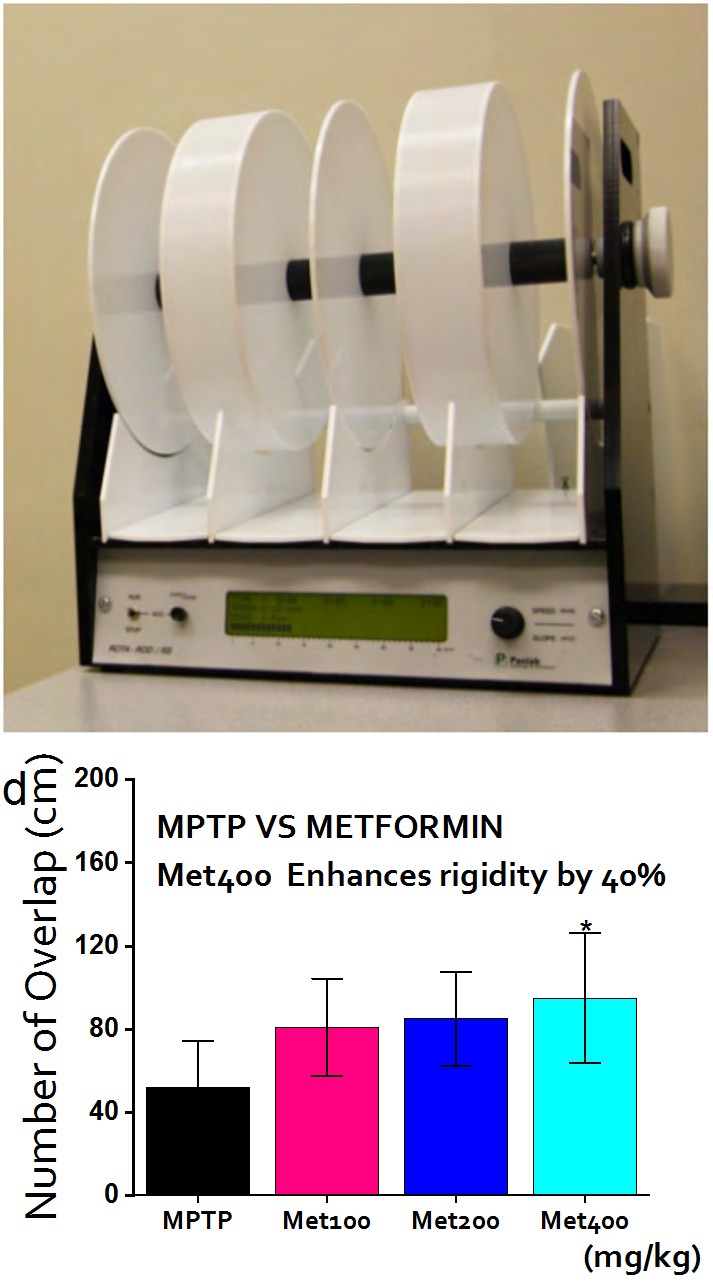
Figure 2: Behavioral Assays I Figure 2a: Trajectory of Open Field Test (OFT): The mice were allowed to move freely in an open field rectangular space and the corresponding movement (locomotion) was recorded by ANYMAZE software and video system. centre zones and corner zones were selected accordingly. MPTP-treated mice move more predominantly within the corner zones in contrast to the saline (control) that move more evenly. Figure 2b: Rotarod: Rotarod set-up showing the lane/rod on which mice is expected to maintain a balance at 5 t0 40 rpm. The latency to fall time was recorded and analyzed accordingly. Figure 2c: Footprints overlap (MPTP VS Saline): The number of overlap indicates the regularity and order of movement. Expectedly, MPTP models showed significant limited number of overlap (P < 0.001). Figure 2d: Footprints overlap (MPTP VS Metformin): Metformin reverses some of the motor deficits caused by MPTP (P < 0.01).
Amperometric DA Recording in Striatal Slices
Amperometric recording in dorsal striatum slices were made using Carbon-Fiber Electrodes (CFEs). Mice were anesthetized with isoflurane (1.5 g/kg, intraperitoneally) and transcardially perfused with approximately 50 ml ice- cold artificial cerebrospinal fluid-A (Sectioning CSF) (110 C5H14NClO, 2.5 KCl, 0.5 CaCl2, 7 MgCl2, 1.3 NaH2PO4, 25 NaCO3, 25 glucose mM, saturated with 95% Oxygen and 5% Carbon dioxide). Next, the brain was rapidly removed and cut into 300 micrometer horizontal slices on a vibratome (Leica VT 1000; Nussloch, Germany). Slices containing the striatum were collected at +0.0 to 1.2mm from bregma. Slices were allowed to recover for 30 min in another artificial cerebrospinal fluid-B (recording CSF) (125 NaCl, 2.5 KCl,
2 CaCl2, 1.3 MgCl2, 1.3 NaH2PO4, 25 NaCO3, 10 glucose mM, saturated with 95% oxygen and 5% Carbon dioxide at 370C, and then kept at room temperature for recording. CFEs 7 micrometer in diameter with an approximate 200-micrometer sensor tip was used to measure DA release in the striatum. The exposed CFE tip was completely inserted into the sub-surface of the striatal slice at an angle close to 300. A holding potential of 780 mV was applied to the electrode by an EPC9/2 amplifier and controlled by pulse software (HEKA Electronic, Lambrecht/pfalz Germany). Single electrical field stimulation (Estim) pulses (0.2 ms, 0.6 mA) or trains (10 pulses at 20 Hz) were delivered through a bipolar platinum electrode (150-micrometer in diameter) and generated by a Grass S88K stimulator (Astro-Med). The amperometric current (Iamp) was low-pass filtered at 100 Hz and digitized at 3.13 kHz. The amplitude of amperometric current Iamp is proportional to the local DA overflow concentration [DA] with a calibration factor of 1 pA for nearly 6 nM. Off-line analysis was performed using Igor software (WaveMetrix).
Statistical Analysis
Results are presented as the mean ± standard error of the mean (SEM). Statistical significance was assessed either via an unpaired 2-tailed Student’s t-test for two-group comparisons or an ANOVA test with Turkey’s HSD post hoc analysis for comparison of more than three groups. Statistical difference was considered significant at the level of P < 0.05, 0.01, 0.001 (5%, 1% & 0.1% alpha). The results were analyzed using IgorPro, OriginPro18, SPSS 13.0 (Statistical Packages for Social Sciences). Trajectory path of the open field were obtained directly from the software (ANYMAZE) attached to the video system.
Results
Metformin Enhances motor Performance
To investigate the neuroprotective effects of metformin in PD, we study the different behaviors displayed by the saline, MPTP and (MPTP + Metformin) groups 2-weeks after the last MPTP injection and compare them accordingly. ROTAROD experiment: MPTP-treated group vehemently reduced the latency to falling to 40% when compared to saline whereas, metformin-treated group improved MPTP-induced motor deficits by 24%. In essence, metformin recovered virtually more than half of the motor dysfunction imposes by MPTP (Figure 3a-e).
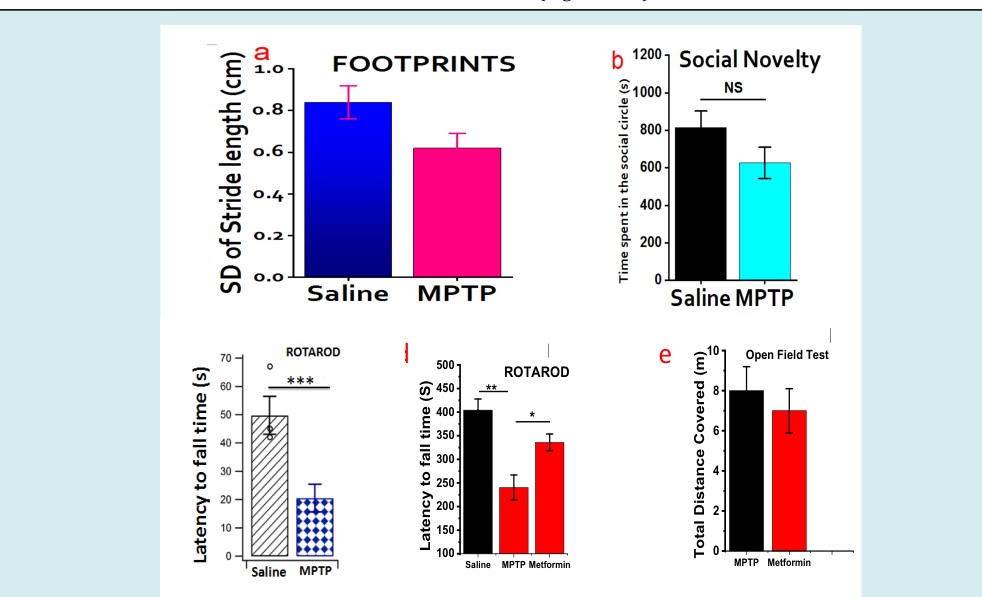
Figure 3: Behavioral Assay II: Figure 3a: Stride Length: Footprints of the MPTP and Saline. Concentrated stride length in the MPTP models indicates the lower numbers of overlap as well as irregularity in their movement. Figure 3b: Open Field Test (OFT): displayed the time spent within the center of the rectangular area and unarguably the PD- like symptoms manifested well in the MPTP model as they intend to avoid the center (even though, statistically insignificant at P < 0.05). In essence, this is due to the lack of proper balance or depression. Figure 3c: Rotarod (Saline Vs MPTP): Expectedly, Latency to falling time in the Saline is significantly longer than that of MPTP (P < 0.001). Figure 3d: Rotarod (Saline Vs MPTP Vs Metformin): Metformin greatly increases the time sustained on the rod in the same group of mice treated with MPTP. Surprisingly, during the 2-week training phase, no much difference with the MPTP group. Figure 3e: Open Field Test (MPTP Vs Metformin): Here, the distance travelled by MPTP- and Metformin-treated mice was almost the same. This indicate that PD model have no restricted movement. Only the lack of balance and perhaps social novelty.
Toxicokinetics and Time Course of MPTP
It is imperative to barely assess the toxicokinetics of MPTP and time course of dopaminergic damage. To achieve this, we investigated the DA release at different period (days) to be able to know the MPTP concentration that can produce best and negligible (if any) damage. The different DA-release we observed were compared to ensure that none of the differential toxicity is related to a simple interference with MPTP toxicokinetics. Having known most studies normally assess striatal MPP+ level few hours after MPTP injections. Here, we analyze the trajectory of MPP+ toxicity by evaluating the release of DA in the striatum. We observed
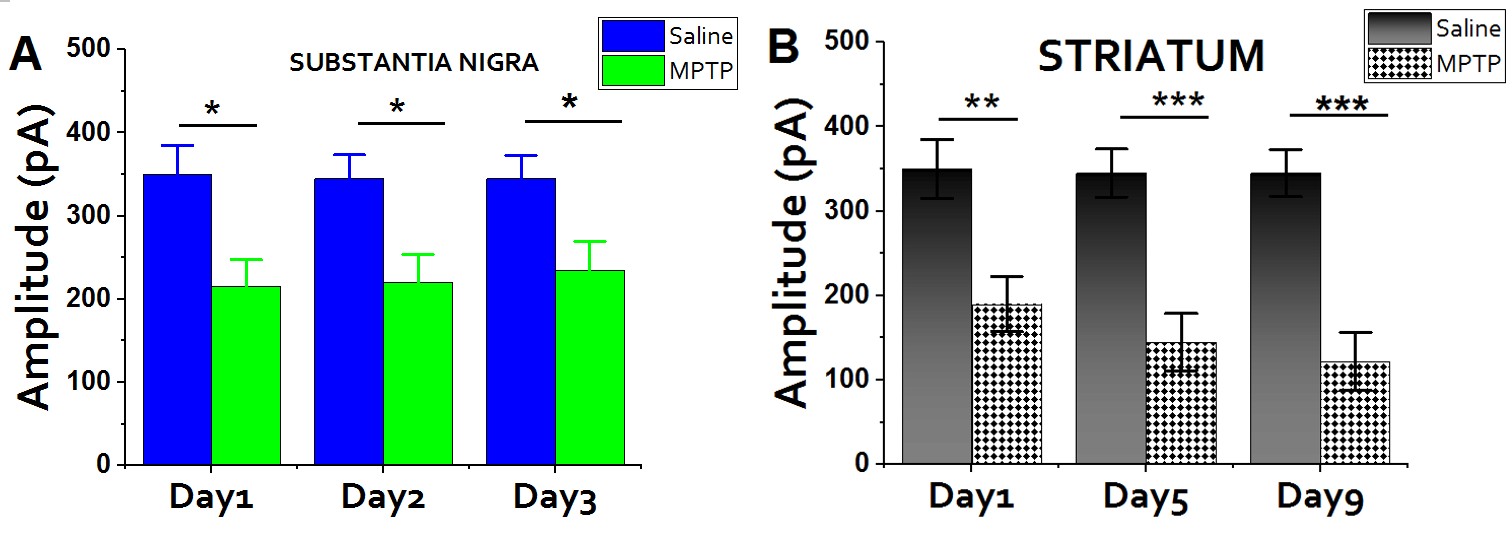
Figure 4: Time Course of MPTP: Having adopted the sub-acute (acute) system of administering MPTP, we deemed it is important to evaluate the level of damage in both the SNc and Striatum. Figure 4a: SNc: In the SNc, The level of DA release is slightly different. Figure 4b: Striatum: The profound damage was much more intense in the Striatum and also there is a gradual increment in the level of damage. We intentionally isolated some mice to test the see what will happen upon exceeding the day 5. Amazingly, the effects produce from day 6 through day 9 is the same. This confirmed that 25-30mg/kg of MPTP requires only 5 consecutive days to reproduce the desirable dopaminergic lesions.
Metformin Improves DA Release
A profound reduction in DA-release was found in striatal slices treated with MPTP as assessed by carbon- fiber electrode (Figure 5c). Metformin (100, 200 & 400 kg/mg) rescue/recovered about 42.5% of the DA neurons exposed to MPTP insults. Amazing, maximum amplitude was a massive and gradual reduction in amplitude from day1 to day5 (Figure 4a). The time course of neurodegeneration enables us to determine when the lesion starts and when it no longer progresses (steady state). Striatum, being the region of dopaminergic projections, the effect of MPTP was found to be more prominent and a wide gap was observed when compared with the control group (Figure 4b), contrary to the SNc (Figure 4a). Importantly, the overall effect of acute MPTP administration seem to be the same after the first 5 days; a hallmark proving that administrating 25 mg/kg consecutively for 5 days is good enough in reproducing the PD model. Meanwhile, days fewer than above are ineffective.
recorded at medium concentration of 200 mg/kg (Figure 5d). However, the neuroprotective effect of the magic drug, metformin was clearly evident even though not as esoteric as the one we observed behaviorally. In essence, Metformin can trigger the release of DA in the striatum haphazardly and arbitrarily dose-dependent manner (Figure 5c,d).
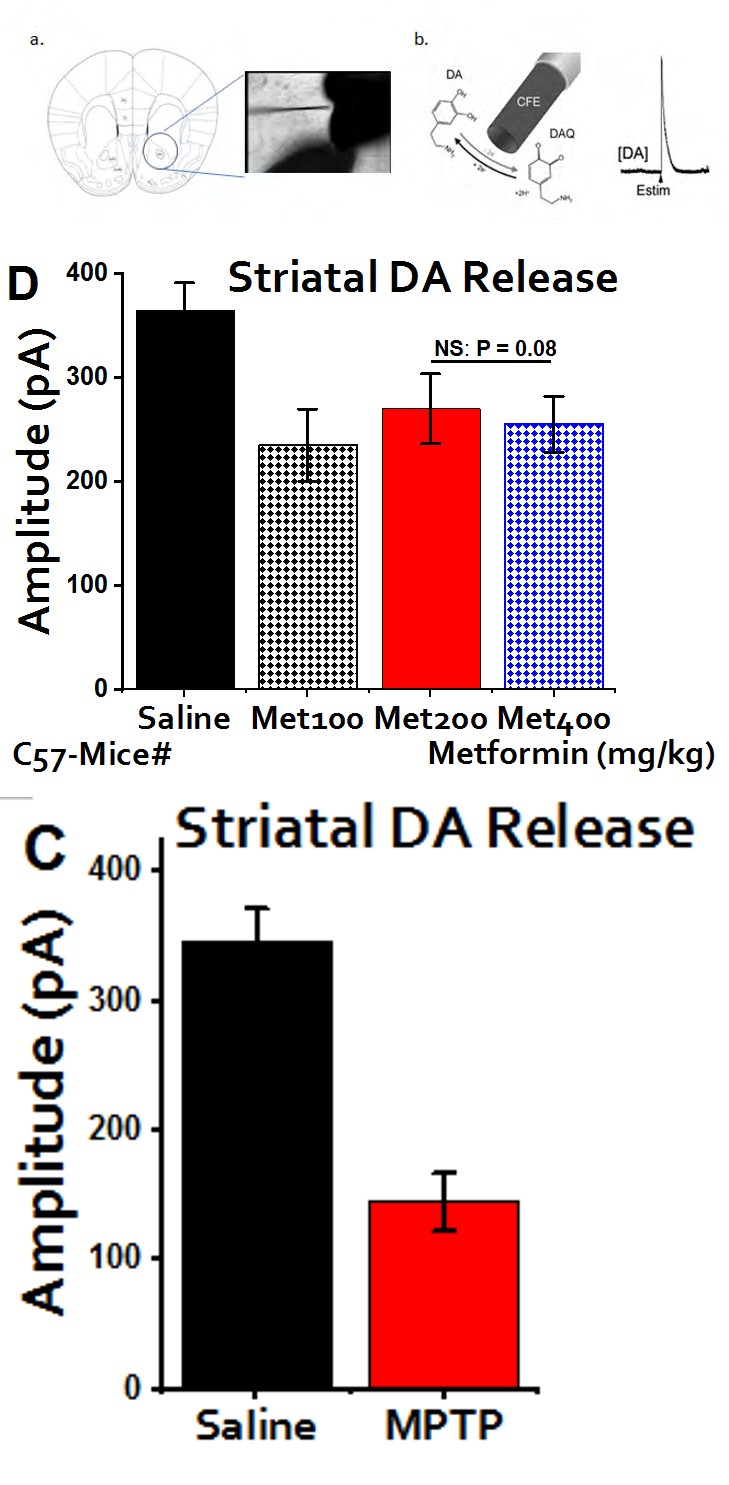
Figure 5: DA Release Figure 5a: Midbrain: Shows the arrangement for measuring the DA release in the striatum and substantia nigra. Figure 5b: Carbon-Fiber Electrode: Carbon fiber is used to detect the release of DA. Estim serve as electrophysiological stimulator that triggers the release of DA at different frequency. Figure 5c: DA release of MPTP-mice: striatal slices of the brain treated with MPTP shows a massive reduction in the DA release when compared to the saline (P < 0.05).Data were collected from three different experiments. This further confirms the successful administration of MPTP to the target area and paves way to further tests. Figure 5d: protective effects of metformin at different concentrations: Surprisingly, highest amplitude was detected at 200mg/kg. Metformin displayed a certain level of DA replenishment against the effect induced by MPTP even though, insignificant statistically (P = 0.08). Data shown are expressed as mean ± SEM and unpaired students t-test.
Discussion
Metformin has been a major and globally accepted antidiabetic drug since 1958. It is potency, efficacy and mechanism of action (MOA) were proved unquestionably for more than half a century. Chronic administration of metformin (100, 200, 400 mg/kg) orally for 3-weeks to C57 mice reduces oxidative stress, improves DA release and motor deficits associated with MPTP-treated mice [21]. Hence, metformin has a protective effect in PD. More so, metformin significantly (P < 0.05) improved the locomotor & postural balance in the MPTP-treated mice. DA release in response to Metformin seems to be surprising as the amplitude fails to show further increase at 4oo mg/kg. In the behavioral assays, PD models displayed some level of depression and this is in line with findings of El-Nabarawy [22].
Conclusion
Effective therapeutic drugs candidates against the episodic insults leading to the motor and non-motor symptoms of PD remain elusive. Although, Levodopa was considered a gold-standard in this regard, it is long-term side effects is really a big limitation in its magnanimity. Metformin is one of the options explored recently and in this study, we were able to establish its decisive role in improving the motor deficits induced by MPTP. Metformin significantly (p < 0.05) improves the DA release in the striatum (when compared to the MPTP group).
Further Direction
Our study concentrated mainly on assessing and addressing the motor symptoms. More research needs to be carried out to evaluate the potential of Metformin in averting the non-motor symptoms. If possible, clinical trials should be performed to figure out whether or not there is an established link between the state of depression & anxiety in PD patients and metformin-receiving patients (perhaps T2DM patients). The above exercise should be a bit tedious since metformin was yet to be considered a conventional drug for PD patients and as such, there must be involvement of T2DM patients.
Declaration of Conflicting Interest
The Authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was intuitively supported by the National Natural Science Foundation of China (31670843, 31400708, 81571235, 21790390 and 21790394), the Key Research and Development Program of Shaanxi Province, China (2017SF- 113) and Fundamental Research Funds for the Central Universities (2017qngz10).
Authors Contribution
DUI conceived and designed the experiments: DUI & BGK performed the experiment. MA Analyzed the data. AS contributed with reagents, materials & analysis tools. All the authors contributed equally in preparing the manuscript.
Acknowledgment
I will like to express my deepest and sincere gratitude to Fidelity Team for their support. My utmost thanks to Professor Wang Changhe for his support and training. A special thanks also goes to Professor Ibrahim Bello Kano for critically reviewing the manuscript before submission.
References
-
Pickrell AM, Youle R (2015) The Roles of PINK1, Parkin, and Mitochondrial Fidelity in Parkinson’s Disease. Neuron 85(2): 257-273.
-
Santiago JA, Potashkin JA (2013) Shared dysregulated pathways lead to Parkinson’s disease and diabetes. Trends Mol Med 19(3): 176-186.
-
Lang AE, Lozano AM (1998) Parkinson’s disease. First of two parts. N Engl J Med 339(15): 1044-1053.
-
Lees AJ, Hardy J, Revesz T (2009) “Parkinson’s disease” The Lancet 373(9680): 2055-2066.
-
Tanner CM, Kamel F, Ross GW, Hoppin JA, Goldman SM, et al. (2011) Rotenone, Paraquat, and Parkinson’s disease. Environ Health Perspect 119(6): 866-872.
-
Sherer TB, Richardson JR, Testa CM, Seo BB, Panov AV, et al. (2007) Mechanism of toxicity of pesticides acting at complex I: relevance to environmental etiologies of Parkinson’s disease. J Neurochem 100(6): 1469-1479.
-
Kakish J, Lee D, Lee JS (2015) Drugs which bind to alpha- synuclein. Neuroprotective or neurotoxic ? ACS Chem Neurosci 6(12): 1930-1940.
-
Tayara K, Espinosa-Oliva AM, García-Domínguez I, Ismaiel AA, Boza-Serrano A, et al. (2018) Divergent Effects of Metformin on an Inflammatory Model of Parkinson’s Disease. Front Cell Neurosci 12: 1-16.
-
Johannessen JN, Chiueh CC, Herkenham M, Markey CJ (1986) Relationship of the in vivo metabolism of MPTP to toxicity. In MPTP: A Neurotoxin Producing a Parkinsonian Syndrome, pp: 173-186.
-
Jackson-lewis V, Przedborski S (2007) Protocol for the MPTP mouse model of Parkinson’s disease. Nature Protocols 2: 141-151.
-
Somayajulu-Nitu M, Sandhu JK, Cohen J, Sikorska M, Sridhar TS, et al. (2009) Paraquat induces oxidative stress, neuronal loss in substantia nigra region and Parkinsonism in adult rats: neuroprotection and amelioration of symptoms by water-soluble formulation of coenzyme Q10. BMC Neuroscience 10(88).
-
Javitch JA, D’Amato RJ, Strittmatter SM, Snyder SH (1985) Parkinsonism inducing neurotoxin, N-methyl-4- phenylpyridinium: uptake of the metabolite N-methyl- 4-phenylpyridinium by dopamine neurons explain selective toxicity. Proc Natl Acad Sci USA 82(7): 2173- 2177.
-
Sharma S, Kumar P, Deshmukh R (2018) Neurochemistry International Neuroprotective potential of spermidine against rotenone induced Parkinson’s disease in rats. Neurochem Int 116: 104-111.
-
Gao HM, Liu B, Hong JS (2003) “Critical role for microglial NADPH oxidase in rotenone-induced generation of dopaminergic neurons. Journal of Neuroscience 23(15): 6181-6187.
-
Shaikh SB, Nicholson LFB (2009) Effects of chronic low dose rotenone treatment on human microglial cells. Molecular Neurodegeneration 4(1): 55.
-
Bayliss JA (2016) Metformin Prevents Nigrostriatal Dopamine Degeneration Independent of AMPK Activation in Dopamine Neurons, pp: 1-15.
-
Curry DW, Stutz B, Andrews ZB, Elsworth JD (2018) Targeting AMPK Signaling as a Neuroprotective Strategy in Parkinson’ s disease. Journal of Parkinson’s Disease 8(2): 161-181.
-
Ming L, Lu M, Su C, Qiao C, Bian Y (2016) Metformin Prevents Dopaminergic Neuron Death in MPTP/P- Induced Mouse Model of Parkinson’s Disease via Autophagy and Mitochondrial ROS Clearance. Int Journ Neurospharm 19(9).
-
Kang H, Khang R, Ham S, Jeong GR, Kim H, et al. (2017) Activation of the ATF2/CREB-PGC-1 α pathway by metformin leads to dopaminergic neuroprotection Oncotarget 8(30): 48603-48618.
-
Tatton NA, Kish SJ (1997) In situ detection of apoptotic nuclei in the substantia nigra pars compacta of 1-methyl-1,4-phenyl-1,2,3,6-tetrahydropyridine-treated mice using terminal deoxyribonucleotidyl transferase labelling and acridine orange staining. Neuroscience 77(4): 1037-1048.
-
Patil SP, Jain PD, Ghumatkar PJ, Tambe R, Sathaye S (2014) Neuroprotective effect of metformin in MPTP induced Parkinson’s disease in mice. Neuroscience 277: 747-754.
-
El-Nabarawy SK, Radwan OK, El-Sisi SF, Abdel-razek AM (2015) Comparative Study of Some Natural and Artificial Food Coloring Agents on Comparative Study of Some Natural and Artificial Food Coloring Agents on Depression, Anxiety and Anti-Social Behavior in Weanling Rats. IOSR Journal of Pharmacy and Biological Sciences 10(2): 83-89.
- Evaluation of Skin Aging Preventive Effects of Cherry Blossom Petal Extracts Through Antioxidant and Anti-Glycation Activities
- Is Cell Death Responsible for False Positive Results of In Vivo Comet Assay?
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism