General Overview of Yeast and Yeast Cell as a Novel Model for Investigations
Biological scientists have been looking for a suitable experimental organism which can provide better understanding of biomolecular mechanisms involved in the physiological processes and observations can be readily interpolated. It can be easily procured, cost effective, and better maintained in laboratory conditions. Yeast is practically omnipresent in most of the biomes and exhibits higher genetic diversity in comparison to most of the angiosperms or chordates. These eukaryotes have very simple and short life cycle exhibiting budding, and sexual reproduction. Mostly, there are no ethical issues related to this organism being used as experimental model. Its small genome is the prime factor that makes easy manipulations in the field of molecular genetics, genomics, evolutionary genomics, senescence, cell cycle, biomedical genetics, and biotechnology. This experimental model opens new horizons in the direction of functional genomics that may be helpful in encoding metabolic mechanisms and ecological diversities.
Overview
Yeast is a bio system that has undergone tremendous investigations in the recent past. This organism is one of the most suitable models for studying gene sequence and comparative genomic investigations. The molecular investigations along with gene sequencing related to phylogeny helps to evaluate the biotechnological applications, Investigation and metabolic pathways [1].
General Biology and Structure
Yeast as a species comes under unicellular fungi, and the cells of yeast are different from bacteria. These organisms have well defined cell organelles such as nucleus, Golgi apparatus, mitochondria, endoplasmic reticulum, vacuole and cytoskeleton, like any matured cells of eukaryotes. These cells typically measures 5X10 µm. Yeast reproduces via budding, sometimes fission, and sexual reproduction (preferably during unfavorable conditions). Their identification is based on cell morphology, physiology, immunological and molecular biological techniques. These are present in soil, water, plants, animals, insects which have special habitat of plant tissues [2].
Viewing A Yeast Cell
Under normal light microscope, yeast cell cannot be seen distinctly, even at 1000X magnification, mostly, vacuole and cytosolic inclusions are visible. To view yeast cell distinctly, phase contrast microscopy is useful. The stained yeast cells are better for viewing but the stained cells become unsuitable for physiological studies. Fluorescence microscopy is helpful to highlight the features of yeast cell using fluorescent dyes such as Calcofluor, DAPI, and Rhodamine etc. Calcofluor helps to visualize white bud scar and it interacts with chitin in scar and it fluoresces. The 4, 6-diamidino-2-phenylindole (DAPI), specifically, causes fluorescence in DNA of the nucleus. DAPI causes the mitochondria to fluorescence and these organelles appear pink and white. Rhodamine also helps to visualize mitochondria. Neutral red stains vacuoles appear purple red [3, 4, 5]. The application of mono-specific antibodies technique against structural proteins in combination with fluorescent dyes like fluorescein isothiocyanate (FITC) or Rhodamine-B facilitate the visualization of more cellular features of yeast cell [6]. The iodine stains glycogen and its deposits get stained as red-brown. Methylene blue stains non-viable whole cells as blue. Aminoacridine and F-C ConA stain cell walls and indicate specific mannan present [5]. Electron microscopy, scanning electron microscopy, transmission electron microscopy, and atomic force microscopy facilitate the investigations related to ultrastructure of organelles and molecular architectural aspects [5, 7, 8].
Generalized Structure of Yeast
The generalized cell structure of a yeast cell is dependent on the physical and chemical fluctuations occurring during their growth. Their size varies between 2 mm to 50 mm in length while width varies between 1 to 10 mm depending on the species. These appear to be ovoid, ellipsoid, hemispherical, filamentous-as septate hyphae or sometimes cylindrical in shape. On an average the larger diameter is around 5 to 7mm and smaller diameter is about 1 to 7mm. The cell size increases during brewing process while in lab conditions the size is smaller. Each yeast cell is encased with thick cell wall (Figure 1).
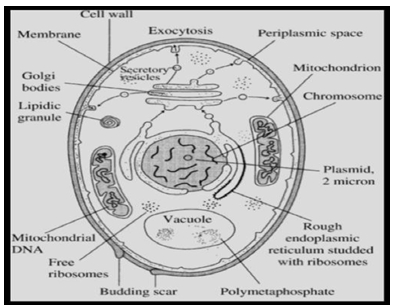
Topography of Cell Surface
Topography of cell surface reflects on the nature and the morphology of cell wall of yeast and it appears different during its growth, and during budding phase. There is a distinct birth scar and it is well marked as circular ridge like structure or raised ridge like structure while the scar appears as convex depression or plate like with or without raised annulus. In some species, the cell surface shows many protrusions in case of Leucosporidium yeast and in some species like C. slooffii, the surface is smooth [9]. The scanning electron microscopy illustrates such features in case of young bud, yeast cell and a grown-up yeast cell.
Cell Wall of Yeast
The cell wall is structurally and functionally dynamic cell organelle, and it secures shape and size of a yeast cell. The cell wall participates in the physiological, biochemical and molecular processes in addition to growth, budding, and cell division. Cell wall also maintains the specific functional mechanical strength, osmotic pressure, and other forms of stresses; these stresses are due to ambient environment [10]. The cell wall of yeast is stiff and hard layer having 100
to 200 nm thickness. It comprises about 25% of dry mass of the cell. The basic structural components of cell wall are highly glycosylated glycoproteins-the mannoproteins, b-glucans and chitin. It consists of glucomannoprotein which associates with β-1.3 glucan and β-1.6 glucan. The molecules of glucomannoprotein link via disulphide bonds. The mannoproteins get bound with chitin β-1.4 polyN- acetylglucosamine, these in turn, get associated with plasma membrane. The enzyme invertase acts as marker and is associated with cell wall of yeast. There are three membrane bound synthetases such as Csh-1 (a repair enzyme), Csh-2 (participate in the formation of septum), and Csh-3 (Cds) helps in cell wall maturation and also in the formation of bud ring) [5]. The composition of cell wall exhibits variations in different conditions of growth and the synthesis of the required macromolecules. The space and time factors also play an effective role in maintaining its composition. The yeast cell wall can be removed if it is treated with lytic enzymes under the influence of osmotic stabilizers [9]. During this process mostly, no harm is done to its structure and functionality. The resultant cellular product is called ‘spheroplast’ or commonly ‘naked cell’. This state of yeast is suitable to study the regeneration and development of cell wall, and also to investigate intergeneric and intrageneric cellular fusion. The glycosylphosphatidylinositol (GPI) is one of the main constituents of cell wall protein and its attachment site referrers to as Pst1p, and it is produced during regeneration of protoplast. Commonly, yeast cell exhibit aggregation and due to the aggregating behavior among the cells and it is called asexual aggregation [5]. The ability to adhere reversibly to each other results in macroscopic flocsedimentation/flocculation of yeast cells. This feature is commonly observed during brewing process. The probable explanation for the mechanism of flocculation is not well understood and it is believed that specific lectins (also named as flocculins), are present in cell wall of yeast. These surface glycoproteins directly bind with mannoproteins of the neighboring yeast cells, and result in flocculation. This feature may participate in sexual aggregation. The subunit of agglutination like A-galp is O-glycosylated protein and has N-terminal producing signals and C-terminal signal to glycosylphosphatidylinositol (GPI) protein that ensures anchorage. Agglutinin proteins help in differentiating a- and b-cells [5, 9].
Periplasmic Space
The periplasmic space is an enclosure bounded by cell wall externally and plasma membrane internally. In this space, mostly, proteins like invertase and acid phosphatase are present which do not permeate via cell wall. These enzymes catabolize those substrates which do move across plasma membrane. These enzymes act as markers for the functionality of cell wall and plasma membrane. The signal sequence of invertase and phosphatase are very helpful in the recombinant DNA technology [5].
Plasma Membrane
This membrane is basically a lipid bilayer harboring specific proteins that anchor cytoskeleton components, enzymes needed for synthesis of cell wall, signaling pathways, and cellular transportation. Its thickness is around 7.5 nm and at places it invaginates forming protrusions in cytoplasm. The lipid components of this membrane include phospholipids like phosphatidylcholine, phosphatidylethanolamine, and minor proportions of phosphatidylinositol, phosphatidylserine, and phosphatidylglycerol, along with sterols such as ergo sterol and zymosterol. The composition of plasma membrane exhibits fluctuations in its structure and functionality in accordance to growth and the ambient conditions [5]. There is a total seizure of phosphatase activity in the presence of ammonium molybdate (01mol/ml) and sodium vanadate (1.0mol/ml) at 6.0 pH. There exists vanadate sensitive ATPase activity (pH 6.0) in presence of KCl (as activator). This behavior of plasma membrane reflects on the proton flux activity [11]. Vanadate sensitive ATPase activity is a useful parameter to characterize plasma membrane in a bio system [5].
Cytoskeleton of Yeast
A cell has to perform integrated and coordinative functions that ensure its supportive, flexibility, cellular rigidity, normal spatial and life sustaining functionalities. Cytoskeleton facilitates appropriate functional performances depending on the types, location, and assigned functions of the cells. This component is also helpful in the delivering cellular responses to the intracellular, intercellular, biochemical, and molecular fluctuations within a biosystem, and provides survival ‘robust conditions’. Cytoskeleton plays essential and primary role in maintaining cellular shape, size, and elaborate intracellular consortium [12, 13]. Cytoskeleton participates in an effective coordinative role with plasma membrane to accomplish diverse functions like development, control of cellular morphology, polarity, adhesive ability for anchorage, and membrane stability in a cell and yeast cells are no exception. It also cooperates during localizing growth area, budding, development of mutant phenotypes, and maintenance of polarized growth in yeast cell and its buds [14]. The patches of cortical actin are present more in the zone of active cell growth, and deposition of cell wall within the bud. Cable or thick strands of actin are seen within mother and bud yeast cell specifically along the ‘mother-daughter bud axes’. During the early phase of cell cycle patches of ring of actin come to the surface of mother yeast cell, thus, marking the spot for the emergence of new daughter bud. This actin ring is persistent and demarcates the neck of baby bud [15]. This ring disappears as the baby bud grows but cortical actin patch develops and locates at the tip and the lateral sides of the developing bud. During the late phase of development the actin patches disperse randomly on the surface of mother yeast cell and matured yeast bud [14]. The cytoskeleton is very important cellular component, and deals with structural and mechanical dynamic cellular functionalities. This ability is attained due to the assembly and disassembly of the compositional protein sub-units of the components of cytoskeleton. For example, microtubules and microfilaments participate in the physiological processes like mitosis, meiosis, mobility of cell organelles, and formation of septum. The cytoskeleton is asymmetrical in organization, and these aspects become very obvious during vegetative growth of yeast cell [5]. There are about 100 accessory proteins which are conserved, and these proteins coordinate the spatiotemporal assembly and disassembly of the actin protein of cytoskeleton. This represents the mechanism of dynamics of functions of actin. Investigations based on molecular genetics, genome-analysis, real time imaging technique, and ultrastructure are very helpful in understanding the molecular dynamics and physiological functionality [15].
Intracellular Transportation in Yeast
Components of cytoskeleton play significant roles in the processes of intracellular transport, thus, maintain the appropriate distribution of molecular cargo between cytoplasm, cell organelles and non-membranous organelles [5]. Yeast cells exhibit actin cellular based transport, this cellular transport system uses the mechanism which regulates polarized cell growth and separation of its cell organelles before cell division sets in. During bud formation assembly of actin cable starts from bud, and undergoes reorientation of actin cables. This process focuses towards growth and secretion of tip of proposed bud. These cables function as polarized molecular path that resemble the delivery system made of type-V myosin and this set up is the need of formation of daughter yeast cell [5]. In yeast cell, proteins like, myosin, kinesins and dynein are the three major motor proteins which bring about the intracellular transportation. The myosins and kinesins proteins link with polarized cytoskeletal filaments. This process meats its energy requirements from ATP-hydrolysis that enable the movements. These molecules carry the molecular-cargo to the distant site in one direction within a cell. There are motor proteins that make the filaments to slide against each other thereby, creating a driving force; this driving force is similar to that which brings about the migration of nucleus and cell division [5, 16, 17]. There are two isoforms of tropomyosin proteins in yeast cell, namely, Tmp1p and Tmp2p. These are present in addition to myosins. Tmp1p is more pronounce isoform tropomyosin, and it binds with actin cable and also with filaments giving them the required stability. The stabilized actin cables and filaments participate in cellular growth and carries metabolic molecular cargo including many organelles. The acylated form of these proteins exhibit better binding efficiency of actin. The Tmp2p is a minor isoform of tropomyosin and it facilitates the working of the major isoform tropomyosin [5].
Cytoplasm of Yeast Cell
Cytoplasm is the fluid component of any cell and yeast cell is no exception. This medium is the major stage for all the metabolic, biochemical, and physiological transformations. It holds the membrane bound cell organelles, and it is bounded by plasma membrane [5]. The pH is of great physiological significance. This is slightly acidic (pH 5.2) aqueous colloidal medium and lodges micro and macromolecules. Some of the prime macromolecules are proteins, glycogen along with larger macromolecular entities like ribosomes, proteasomes, lipid particles; these inclusions give the suspension appearance to the cytoplasm of yeast cell. In addition to these constituents, some cytosolic non-organellar inclusions like glycolytic enzymes, fatty acid synthase complex, along with these components and enzymes involved in protein metabolic system are also present in cytoplasm. The functional entities relates to the formation and regulation of rostrum of cytoskeleton [5]. Under stress-state, the cytoplasm of yeast cell undergoes down-regulation. The life processes like cell division, growth, and the overall metabolic states function at the lowest ebb during the stress period. But the viability of the yeast cell is retained until the return of the favourable conditions. The mechanism involved during this stress-state is still ambiguous and offers a field to venture. The non- membrane organelle regenerates as a result of reversible polymerization of translation of 2B-initiation factor. This factor is an important enzyme that brings about the synthesis of protein those results in the formation of filaments in the form of large bundles. The correlative light and electron microscopic techniques illustrate this observation. Further, energy requirements, degradation of protein, and functions of proteins get restricted [18].
Nucleus
In yeast cell, the nucleus is spherical, centrally or sometimes excentral in position with diameter around 1.5 mm. A well-defined double membrane encloses the nuclear contents and it distinguishes nucleus from the cytoplasm. This envelop is perforated at intervals, each perforation is called nuclear membrane pore and has diameter within the range of 50 to 100 nm [5]. This set up appears to be channel like and facilitates the transportation of metabolites between nucleoplasm and cytoplasm. The outer membrane is in continuation with that of endoplasmic reticulum. (This feature is similar to the other eukaryotes). This link is retained even during mitotic division. Regeneration of pores in nuclear membrane appears as an important process [5, 19]. The spindle pole body ensures anchorage of nucleus with both continuous and discontinuous microtubules. The spindle pole body is present on the outer phase of outer nuclear membrane. The spindle pole body divides into two poles and helps in doubling the number of chromosomes; this happens prior to isolation of two daughter cells. The nucleolus appears denser in comparison to the remaining nucleoplasm within nucleus. It is the site of location of r-RNA gene, and synthesis of r-RNA. It facilitates the assembling the subunits of ribosomes and also helps in processing of pre-m- RNA [5, 20].
Chromosomes in Yeast
Chromosomes are congustedly arranged within nucleoplasm as chromatin material. Mostly, the nucleosome in case of yeast is around 145 bp of DNA in length. There are variations in the numbers and sizes of chromosomes in different species of yeast. The pulse-field gel electrophoresis is a suitable technique to study their karyotypes [5, 21].
Endoplasmic Reticulum
The endoplasmic reticulum is well organized and is highly branching tubular cell-organelle, constituting around 10% of the volume of a cell. A double lipid layer membrane encapsulates this cell-organelle, and the same layer communicates intimately with the nuclear membrane [5]. Cellular stability, modifications, protein transport, and other regulatory processes are under the control of endoplasmic reticulum. The transmission electron microscopic and dual-axis electron tomographic techniques reveal that endoplasmic reticulum comprises of three primary domains, namely, the plasma membrane associated endoplasmic reticulum (pmaER), the central region central-cisternal endoplasmic reticulum (cecER) and the tubular endoplasmic reticulum (tuber) [22, 23]. The reticulon family members like Rtn1p and Rtn2p present in budding yeast and interact with members of family of protein like Yop10 and DP1 resulting in the stabilization of sheet and tubules present in the region of membrane that exhibit high curvature [23, 24, 25]. The three primary classes of membrane lipids are sphingolipids, phospholipids, and sterols [23, 26, 27]. The endoplasmic reticulum also participates in the storage of calcium, homeostasis and maintenance of concentration of about 10µM free Ca++, at least in wild variety of yeast. The synthesis of calcium sensitive photoprotein i.e., aequorin-a specific targeted protein, is also produced in endoplasmic reticulum [23, 28]. The Golgi apparatus, lysosomes (vacuoles in yeast), endosomes, secretory vesicles, plasma membrane are the product of endoplasmic reticulum, including the endoplasmic reticulum itself. Thus, endoplasmic reticulum is the production site of the most of the transmembrane proteins and lipid of cell-organelles. The membrane of endoplasmic reticulum transports the proteins produced in polyribosomes from cytosol into is lumen. The folding of protein due to chaperon and partly the protein glycosylation occur within endoplasmic reticulum. It is very important that protein must get correctly folded for their effective transfer from the endoplasmic reticulum. The process of packing of proteins takes place in the endoplasmic reticulum within vesicles. These vesicles bud-off from the membrane of endoplasmic reticulum and move to the Golgi apparatus and here these vesicles fuse with its membrane [5].
Golgi Apparatus: (Golgi Complex)
The morphological aspects of Golgi apparatus exhibit variations in different species of yeast. Basically, this cellular organelle is made of membranous stacks arranged in parallel pattern or as dispersed cisternae. The proximal region is cis-Golgi network while distal or internal region is trans- Golgi network. The synchronized process of proteins gains access in Golgi apparatus through cis-face and emerges from trans-face. This synchronized processing of proteins involves formation of complex oligosaccharide chains of N-glycosylated proteins, phosphorylation of oligosaccharide which will be reaching the vacuole [5]. The processes like synthesis of proteoglycan (O-linked glycosylation of proteins), alteration of lipids, sulfation of tyrosine, etc., come in this category [5]. In spite of the existence of varied morphological patterns of Golgi apparatus, the mechanism related to the molecular transport in different patterns is basically similar. The concerned functional aspects like maturation of cisternae, to and fro protein movements, retention of secretory cargo proteins also follow similar mode. Phosphatidylinositol-4- phoshate and phosphatydyl-4-kinase both play major roles in the isolation of multiple proteins within Golgi apparatus. The hydrolysing enzyme Sac-1, phosphatydylinositol-4-kinase and phosphatidylinositol-4-phosphate together regulate the amount of phosphatidylinositol-4-phospahte under the state of glucose deficiency. This indicates that the level of carbon source possibly acts as an external signal to maintain molecular traffic in the Golgi body of yeast cell [29, 30, 31, 32]. The Golgi vesicles carry the molecular cargo to and fro within the parts of Golgi apparatus and endoplasmic reticulum. The endoplasmic reticulum associates the degradation system, thus prevents the transportation of defective molecular cargo. The defective molecular cargo includes misfolded proteins, and any molecule that disrupts functionality of endoplasmic reticulum and Golgi apparatus [5, 29].
Transport Vacuole System
In yeast cell, intracellular transport is one of the vital significant functions. The intracellular distribution of metabolites results in appropriate supply and drainage of biomolecules for successful functioning of yeast cell. The vacuole system of yeast cell carries out this responsibility efficiently. The plasma membrane and other cellular membranes participate in the formation of vacuole system. Mostly, the membrane of different organelles takes part in formation of vesicle and the vesicle gets detached as a result of budding process. The nature of vesicles formed depends on the nature of cargo it has to carry and the cellular target. The membrane of these vesicles has basic bilayer nature but gets additional coating of specific molecules depending on their specific function. Based on the special coating the vesicles are grouped as COP-I, COP-II, and clathrin coated vesicles [5]. COP-I coated vesicles have coating of multimeric protein complex along with GTP-binding protein Arf1p (an ARF). This coating is coatomers. These vesicles are responsible for retrieval (reclaiming) process and starts with cis-Golgi network extending to trans-Golgi network [5, 33]. The three biomolecules are present in endoplasmic membrane, namely, small GTP-binding protein (Sar1p), the Sec1p, and the Sec23/24p complex. These help in the formation of coating of second group of transport vacuoles- the COP-II. The COP-II coated vesicles carry their cargo in forward direction, i.e., anterograde movements from endoplasmic reticulum towards Golgi apparatus [34]. The outward movement gets counter balanced involving inward movement (retrieval move); this mechanism helps in bringing back the selected molecules to their place of origin and these molecules are available for reuse from endoplasmic reticulum. The loading of cargo is not random but this process is systematically selective [35, 36, 37]. As and when there is a need for COP-II vesicles then the enzymes like small GTPase Sar1p and GEF Sec12p which are present in endoplasmic reticulum get activated and these cause in the induction of COP-II vesicle in the transportation of biomolecules [38]. The third group of the vesicle is clathrin coated vesicles. These vesicles bud off from the membranes of cell organelles and plasma membrane. Clathrin coated vesicles participate in direct molecular transport and act as late secretory pathway ensuring endocytic transportation of biomolecules. These vesicles carry cargo between trans- Golgi network and secretory vesicles return them to Golgi network procured back [5, 39]. Drainage and degradation of biomolecules appear to be two primary functions. The drainage vacuoles acts as receptors of proteins from plasma membrane, internalized protein via endocytosis,proteins which need to be weeded out away from secretory process at Golgi apparatus. The drainage vacuoles also receive proteins emerging or representing cataclysmic molecular interactions and organelles remains to be eliminated from cell [5, 40]. The vacuoles in which enzymatic molecular degradation is carried are termed as ‘vacuole degraders’ and the process is referred to as vacuolar degradation. The varied intravacuolar hydrolases like endopeptidases, aminopeptidases, carboxypeptidases, neuleases, glycocidases, lipases, phospholipases, phosphatases etc., bring about the molecular degradation in vacuoles [5, 40, 41]. The specific secretory pathway ensures the delivery of the hydrolysing enzymes and the respective substrates in the concerned vacuoles [5, 40]. Generally hydrolytic activities take place around 5.0 pH while the pH of cytoplasm is around 7.2, this indicates that the vacuole concerned with molecular degradation regulate the pH. These vacuoles are also concerned with some of the physiological processes such as storage for basic amino acids, polyphosphates, and some metallic cations. The membrane of such vacuoles regulates the cytosolic ionic concentrations via homeostatic regulatory process [5, 40].
Peroxisomes in Yeast
The other name of these cell organelles is microbodies, and is present as an encapsulated in single double lipid layered body. These cellular components have hydrogen peroxide forming oxidase and detoxifying catalase. These metabolize the hydrogen peroxide molecules. Peroxisomes metabolize β-oxidation of fatty acids. The number and structural aspects vary depending on the conditions for growth. These are very important for establishing cellular viability [5, 42, 43]. Mitochondria in Yeast Cell The mitochondria of yeast cell is similar to those found in other eukaryotes, and has been a target of investigation due to its significant role during fermentation process. Two types of lipid bilayers encapsulate it, outer membrane and inner membrane. These two layers have a space in between; the inner membrane encloses the matrix of mitochondria. The outer layer acts as a harbor which holds enzymes which metabolize lipids. The inner membrane has cytochromes concerning respiration, ATP synthase to facilitate functioning of respiratory chain, and transport proteins needed for trafficking of components with lower-molecular-weight. The matrix of mitochondria contains mitochondrial DNA and acts as a site of tricarboxylic acid cycle; molecular gadgetry for protein synthesis, and mitochondrial ribosomes. The proteins required in mitochondria are either synthesized or imported. The size and the morphologies of mitochondria depend on the conditions for growth of yeast cell. An appropriate mode of aerobic respiration is ensured as aerobic conditions and suitable amount of oxygen and carbon source is ensured [5].
Nature and Behavior of Yeast
Yeast is able to tolerate higher concentrations of sugars, salts, acetic acid, and under these conditions maintains appropriate rate of growth. Such stress tolerant traits are useful for biotechnological point and enhances the efficacy of the industrial processes and declines the possibilities of contamination due to unwarranted microbes. At the same time the tendency of yeast to resist extreme condition may result in spoilage of food in spite if presence of common preservatives and higher osmotic ambient environment [1, 44, 45]. Yeast is also very effective osmotolerant and is able to maintain its growth under higher osmotic pressure. Molecular mechanism is related to the concentrations of sugar and salts; these two components individually affect the overall degree of osmotolerance of yeast [46, 47, 48]. The process of permeability in yeast is dependent on the rate-limited due the passage of water through plasmalemma and/or cell wall. These two are the first structural and functional barriers for permeability. This hydrophysical process is temperature dependent. The coefficient for hydraulic water permeability is 1 to 2x10ˉ13 cm3/dyne/sec., at 20ºC [49].
Yeast and its Genomics
Yeast- unicellular eukaryotic fungus is mostly omnipresent in any type of biomes and this organism exhibits relatively more genetic diversity as compared to other angiosperms and chordates. Yeast is a member of sub-phylum Saccharomycotina (sometime also called Hemiascomycota. These biosystems can be easily cultured and have small genome having around 10 to 20 Mbp. These features make them one of the most suitable experimental organisms to investigate molecular genetics, biotechnology, and evolutionary genomics. The genetic related studies on yeast open new horizon in the taxonomic, genomic diversify functional metabolism, and ecological aspects. Some of the species of yeast are commensals (Saccharomyces) or pathogens (Candida albicans) [50]. Since, on the onset of genetic engineering large scale reading and writing of DNA along with understanding, modifications and alterations in the genetic code have been the prime aspect of investigations. These investigations have also involved software version Sc-1.0 and Sc-2.0. The Baker’s yeast – Saccharomyces cerevisiae is one of the most favoured experimental organisms [51]. Yeast is useful model for studying genetic engineering. In organisms where inefficient homology- directed repair restricts CRISPR-Cas 9 genome editing, use of non-homologous end joining helps to secure DNA double strand breaks. There are efforts to reduce the application of non-homologous end joining technique. The successful deployment of linear precision editing and random intermediation of non-homologous end joining facilitates optimization of microbial production of (S)-norcoclaurine is possible [52]. Norcoclaurine is a compound present in plant like fruits of Nandina domestica, roots of Aconitum carmechaelii, stems of Galium divaricatum, Annona suamosa, and Nelumbo nucifera (seeds of lotus).Norcoclaurine is also known as ‘higenamine’ and is a dietary ingredient for sports and weight loss schedules but it also causes health risks [53]. Thus, yeast is a valuable experimental model for such type of investigations and can be utilized to the maximum extent during antagonizing pair of DNA double strand break path ways for microbial industries [52].
Yeast and Senescence, Life Cycle and Cell Cycle
Specifically, in medical practices, prolonged life span of an individual is highly expected at least during or post treatment. Otherwise the long life span relates with the age of cells and mechanism of senescence. The most common parameters that relate with life-span of an individual are chronological lifespan and replicative lifespan. The process of longevity is well illustrated in yeast whose genome is well characterized. Other aspect of such studies relates directly or indirectly contributes to the estimation of replicative or chronological lifespan. The genome of human and yeast are well understood and amenable the genetic manipulation and screening of aging related diseases [54, 55]. Chronological lifespan involves growth depending on the nutrients, number of cells per unit volume, metabolic interactions that involve growth factors and attainment of specific level prior to reproductive phase. The replication lifespan is the duration in which a cell or an individual forms daughter cell or offspring and after this phase there is a decline in the replicative abilities [54, 55]. It is possible to regulate the process of longevity in case of yeast and link with metabolic genome [56]. Among eukaryotes and mammalian cells the quantum of food intake i.e., caloric dependent food, is one if the prime parameter that influences the life span, physiological aging, and senescence associated pathogenesis but the mechanism involved is not well understood [57, 58, 59]. The mechanism involved during the utilization of food in the process of aging in not well understood. The metabolites such as alpha-ketoglutarate inhibit the synthesis of ATP-synthase and target of rapamycin (TOR) involve during increase of life span of yeast [60]. Quasi-programmed aging in budding yeast relates with cell proliferation, differentiation, stress-response, survival and death [61]. The scarcity of nutrient like carbon, phosphate and nitrogen influence the cell cycle, like arresting the G1 phase and entry of dividing yeast cell in to G0 (a quiescent phase). The target of rapamycin (TOR) and protein kinase-A (PKA) pathway of signaling impacts the protein kinase Rim- 15 and this regulates the initiation of G0 phase of cell cycle [62]. The Pho80-Pho85 cyclin and dephosphorylation of cyclin dependent kinase (CDK) complex play a major role in controlling the entry of cell in the Gophase [63]. Adversely
affected signaling pathways related to cellular proliferation, physico-biochemical status of cell, like, osmotic, oxidative or replicative stress influence the progress of cell cycle. These parameters indicate the significance of nutritional factors in maintaining a specific rate of cell proliferation [56, 57]. There exists a temporal relationship between the parameters affecting cell cycle and cyclin dependent kinases and other cyclins. There quantum of cyclin-3 proteins remain fairly constant during the progress of cell cycle. The yeast is one of the most suitable experimental models to investigating the mechanisms that regulate life span and cell cycle [55, 60, 61].
Genetic Engineering and Yeast
The ability of DNA to write and read is the bases of genetic engineering and its applications are reaching new technological horizons in the current scenario of genetic engineering and biotechnology. These off shoots of biological sciences involve changes in the genetic code of a biosystem using molecular tools and applications in the industries. The disciplines of synthetic biology concerned with chemical synthesis of genome and chromosome and replace their natural counterparts. The investigations related to their functions are of great significance in biological sciences and industries [52]. The genetic engineering and epigenetic investigations are primarily based on the two experimental models, i.e., Escherichia coli and Saccharomyces cerevisiae. Among these two, Saccharomyces cerevisiae, enjoys preference over the E coli, because this eukaryote is generally regarded as safe and has very efficient homologous recombination system, its genome is completely sequenced [64, 65]. The third generation sequencing technique is quite recent and has the benefit of both long sequencing reads and the massive parallel of the sequencing of second generation but this technique has low degree of accuracy [66, 67, 68].
Epigenetics and Yeast
A consolidated definition of epigenetics may be ‘a phenomenon in which a trait gets inherited as phenotype without any changes in its DNA sequence’. This inheritance is the result of ether mitosis or meiosis. The current attention relates with understanding the basic mechanism involving many distinct but closely knitted molecular pathways that control initiation, maintenance, and the degree of heritability of the epigenetic trait. Basically, there are three primary signaling parameters, namely, epigenator, initiators, and maintainers; these accomplish the process of epigenesis. The specific features of yeast that make it suitable for the epigenetic studies are short compact genome, well characterized genetic and reproductive cycle, short generation time, and the developed suitable technologies [69, 70]. A DNA-binding protein, a noncoding RNA, or another structural unit acts as an initiator and these decide the future assembly of chromatin structure [71]. The intracellular changes have an ability to stimulate the epigenetic phenotype. The changes are intracellular and in the ambient environmental niche, these are epigenator signals. These signals initiate epigenator signaling pathways that lead to the initiation and activation of the epigenetic process. The epigenator signals activate and pinpoint the precise location of epigenetic process, i.e., chromatin environment. This phenomenon maintains the chromatin environment during the first and the subsequent generations.
The process of transcriptional silencing has been well established in Saccharomyces cerevisiae. It takes place at genomic sites like, silent mating-type loci, telomeres and ribosomal DNA, and telomeres. Epigenetic silencing at these sites is because of the absence of most of the histone modifications and deficiency of histone H14 lysine 16 acetylation [72]. Silent loci in Saccharomyces cerevisiae (also referred as budding yeast) and Saccharomyces pombe (also called fission yeast) act as proxy during heterochromatin among higher eukaryotic. These species have been the model to provided enormous information in establishing the epigenetics and its applications. Firstly, the phenomenon of epigenetics helps to provide information about the structure of chromatin, this helps to modify the cellular phenotype that too without any alteration in the DNA sequence. Secondly, it also reflects on the possible inheritance due to the changed chromatin state (modified genetic memory), probably during S-phase of cell cycle [73]. Further, the simplest eukaryotic epigenome and the reductionist epigenomic version of yeast make it very easy to conduct the related studies, specifically in the Saccharomyces cerevisiae. The budding and fission yeast do not exhibit the process of DNA CpG methylation. The RNA interference mechanism and repressive histone H3K9 are absent in Saccharomyces cerevisiae but these do take place in fission yeast and most of the eukaryotes. The existence of methylation of gene body in eukaryotes and its absence in yeast (some forms) reflects on the lineage loss during evolution; thus, the role of histones modification acts as means of demarcating various epigenetic states [74].
Toxicology and Yeast
Yeast cells are suitable model for investigating toxicity because of its convenient genetic manipulation, short life cycle, and easy access to the genomic sequence and are relatively easy to culture. The parameters that act as the bases of modulating the sensitivity in relation to the toxic materials include cellular thiols, like glutathione, phytochelatins, labile sulphide and metallothioneins. The products of gene also participate in transference or partition or isolation of metallic or other toxic materials. These features and pathological conditions have been investigated and confirmed in genomic sequence of Saccharomyces cerevisiae genomic sequence. Even, the molecular mechanisms that participate in regulating the ability of toxicity of metal or other toxic materials can be suitably investigated [75].
An organism encountering toxic or xenobiotic materials exhibits toxicological responses during this process; there are many variations in genetic and protein expression at molecular levels during the process of response. The traditional testing of toxicity has limitations like, slow nature, lower capacity, expensive and restriction to limited endpoints. Since the enormous number of toxic or xenobiotic materials are present in environment affecting the biota. Their toxicity data are inadequate, hence, it is imperative to look for an alternative and effective mode of such investigations. Toxicogenomics is a suitable technique that meets the major criteria. Toxicogenomics includes DNA microarrays, high throughput NMR and analysis of protein expression. Saccharomyces cerevisiae provides integrated observations concerning toxicity mechanisms. Such studies are based on the combined assessment of transcriptomics and quantitative proteomics and these facilitate in evaluating the probable ‘genome-wide expressions’ during toxicological responses. To identify and conclude meaningful conclusions the observations from metabonomics/metabolomics and chemogenomics there must be integration of these observations and suitable tools of bioinformatics [76, 77, 78].
Yeast and Nanotechnology
Nanotechnology is multifaceted, multidimensional, bio remedial enginery and has applications in almost all technologies. It encrypts the basic activities in living and non-living entities of earth and also explains the modes that provide the existence, sustenance, reuse, recycle, and replenish the matter and energy on our blue planet. The biotic and abiotic components show a synchronized and orchestrated mutual coordination. Yeast, a member of ‘fungi’, plays its role as a multidimensional unit as an agent in biosynthesis and delivery agent for nanoparticles, genetic materials, and pharmaceutical drugs involving principles of nanotechnology [79, 80, 81, 82].
Nanomaterials exhibit self-assembly in layered pattern also, and electrostatic interactions hold these layers. This technique is applicable in the non-viral gene delivery process. The self-assembled DNA as nanomaterial (in nanoforms) can be readily delivered to the desired cell. The DNA-nanoparticles caged within ‘hollow yeast cell wall particles’, is successful mode to transfer the genetic material. The core component consists of t-RNA or polyethylenimine having layer of DNA layer, the final layer is of t-RNA or polyethyleneimine. This form of transferring genetic material using yeast cell wall is useful for oral as well as systemic drug administration. The effective aspect is protection against phagocytes. The tagged t-RNA, polyethyleneimine, and DNA with fluorescein and or rhodamine are easy to detect or visualize involving fluorescent microscopy, fluorescence spectroscopy, and flow cytometry quantitatively. The DNA payload depends on the specific ratio between t-RNA and yeast cell wall particles [79, 80]. Synthesis of nanoparticles has been one of the prime aims of nanotechnology to meet the demands of their applications but the cost effectiveness, eco-friendliness, and the environmental derogative implications of nanomaterials compel nanotechnologists to look for the alternative methods. The green biosynthesis is apparently a suitable choice. Nanoparticles like, organic and inorganic origin nanoparticles and quantum dots have been successfully procured form plants, bacteria, yeast, and lower organisms. Louis Pasteur is a prominent pioneer in this field and found that microorganism like yeast, is capable of chemical conversion in absence of oxygen, and results the formation of alcohol from sugars [83, 84]. Yeast plays major role in the fermentation industry because of the presence of reducing enzymes in them; this process is either extracellular or intracellular in nature. Same concept is applicable for the green biosynthesis of nanoparticles [80]. During the synthesis of metallic nanoparticles, the source of the element of the nanoparticle in question should be present in the reaction mixture in a nontoxic range i.e., 1ppm to 1000 ppm. This amount is to be decided during early pilot experiments. In the normal course of experimentation, the presence of metabolites like carbonyl groups, phenols, amines, terpenoids, proteins, alkaloid and/or any suitable reducing metabolites is a need for the successful biosynthesis of metallic nanoparticles [85, 86].
Yeast strains, like Candida glabrata and Saccharomyces pombe, are most suitable for intracellular synthesis of silver, selenium, titanium, gold, and cadmium sulfide nanoparticles [87, 88]. Extracellular synthesis of silver and cadmium sulfide nanoparticles using these organisms is also feasible [89, 90]. Probably, in yeast, the membrane bound enzymes, like oxidoreductases and quinones, participate in the synthesis of nanoparticles. These enzymes are pH sensitive [91], and become active during stress conditions [89]. Generally, the stress is the result of additional or reduced presence of metallic nutrients in the culture media that commences the stream of interactions resulting in the formation of bio- production of phytochelatins synthase and glutathione. These enzymes help to counteract the cellular stress [89]. Further, these enzymes have specific redox and nucleophilic properties and these facilitate bioreduction of metal ions; this aspect is the bases of formation of nanoparticles. The binding of metallic ions helps in the formation of nanoparticles, like cadmium, selenium, zinc, gold, nickel, copper and other metals [92]. In all probability the intracellular biosynthesis of metal nanoparticles is a mode of regulating cellular metallic ionic stress [90]. Yeast cells produce nanoparticles intracellularly or extracellularly. Such cells are suitable sources for nanoparticles and such nanoparticles can be readily extracted and purified, using techniques, like centrifugation, dialysis of media in which yeast has been cultured for the biosynthesis. The yield of nanoparticles can be examined physically and chemically [93]. There are chances that nanoparticles exhibit aggregation or degradation; such behaviors relate with the technique involved and/or physicochemical characters of the reaction medium. The applications of such biosynthesized nanoparticles relate with their specific behavior like, antimicrobial, antiviral or anticancer [94, 95, 96, 97, 98]. The mechanism of such behaviors is due to the production of active oxygen species, damaging cell wall of bacteria, induction of inflammation or blocking the metabolic pathways of the concerned pathogen [98].
Yeast and Pharmaceutical Sciences
Unicellular yeast is unique organism that exhibits fundamental mechanisms relating with replication, cell division, recombination, and metabolism that are seen in eukaryotes and other prokaryotes. The species of Saccharomyces cervices is one of the well-studied eukaryotic organisms as its complete genome is adequately established [99, 100]. Further, its short life cycle, ability to be cultured easily, and fast growth rate make it more favorable model for investigations concerning with pharmaceutical sciences [101]. Baker’s yeast (S. cerevisiae) is suitable material to synthesize antibiotic nanobiosomal peptide penicillin. Pharmaceutical products like insulin, human serum albumin, vaccines, and antibiotics like penicillin [101, 102, 103]. Some of the unconventional yeast species like Hansenula polymorpha, Pichia pastoris, Yarrowialipolytica, Kluyveromyces marxianus, Kluyveromyces lactis and Scheffersomyces stipites are utilized as sources for biopharmaceutical products [103].In the recent times a trend is set relating yeast i.e., yeast has been the focal point for the biopharmaceutical science [104].
Yeast biotechnology provides products like crude glycerol, lignocellulose, corn starch, cane sugar, primary and secondary metabolites, and also recombinant proteins. Recombinant pharmaceutical proteins play prime role in the financial, production, and pharmaceutical aspects. Yeast is one of the most sought after cellular organism for this multi-billion dollar industry. The current advances in glycoengineering of yeast along with secretory mechanisms involved in yeast provide favorable and encouraging results. Two modes of investigations namely, strain and pathway engineering help in understanding the mechanism involved and provide appropriate homogeneity and higher production/yield. In this aspect tools of synthetic biology and omics technology are very helpful [105, 106]. Some non- conventional species of yeast like, Kluyveromyces marxianus, K. lactis, Yarrowia lipolytica, Pichia pastoris, Scheffersomyces stipitis, Hansenula polymorpha, and Rhodotorula toruloides show desirable phenotypes such as theromotolerance, ability to assemble carbon sources, and to secrete higher protein and lipid titers. There is a need to carry investigations in the field of advanced metabolic engineering and to explore the possible biological systems along with suitable methods [107].
Behavior of Fungi in Relation to Radiation
Fungi are able to take advantage of radiation and utilize their energy in synthesizing food for their survival. It is proposed that melanin plays significant role in utilizing energies present in these radiation as chlorophyll pigment does from solar radiations in green plants. Melanin is ubiquitous in biological world and most of the fungi possess this biomolecule if not all fungi. Its role in fungi is mysterious and need thorough investigation. Fungi are able to use the energy contained in these radiations during the synthesis of their food which, results in appropriate growth. This is the case specifically in fungi have melanin. The soil micro- organisms which are contaminated with either present in traces or otherwise, have melanin show this behavior indicating the probable role of melanin in the biochemical pathways. When the melanin present in fungi gets exposed to ionizing radiation and even other forms of electromagnetic radiations, it undergoes electronic modification. This feature is not seen in those organism in which melanin is absent. Thus, possibly melanin plays an important role in apprehending energy of electromagnetic radiations. The ability of fungi to exploit their oxidative reduction abilities and the efficacy of melanin enables them to trap energy of these radiations to gain and maintain its metabolic energy requirements. Crytococcus neoformans, a fungal species not having natural melanin and Wangiella dermatitidis, a fungal species having natural melanin did show elevated concentration of melanin as a result of exposure to ionizing radiation with 500 times higher level than the background level and exhibit higher degree of growth. Melanin formation gets induced in Cryptococcus neoformans and its amount increases after the exposure. This feature is established based on the number of colonies formed and the gain in their respective dry-weights. The electronic fluctuation in the structure of melanin enables their ability harness energy from these radiations and exploits it for its metabolic needs (Figure- 2). One of the prime feature of fungal melanin is that it is similar to that present in mammalian skin [108]. Any bio-cell when provided with culture medium or any medium, like, the frequencies for different types of non- ionizing radiation causes some degree of stress and it is in response to the properties of the medium. The physiology of cellular response is in accordance to the fluctuations in the ambient environment and the ability of cell of a biological system to adjust and reflects on its efficacy to survive in the changed circumstances [109, 110]. The culture of Serpula himantioides, fungus, on exposure to Wi-Fi radiations exhibits elevated fatty acid contents and ergosterol.
Ergosterol plays significant role in vitamin D metabolism. Some form of modification takes place in their cell membrane and enhances antioxidative ability. This feature provides means to improve their dietary efficacy [111] Figure 2.
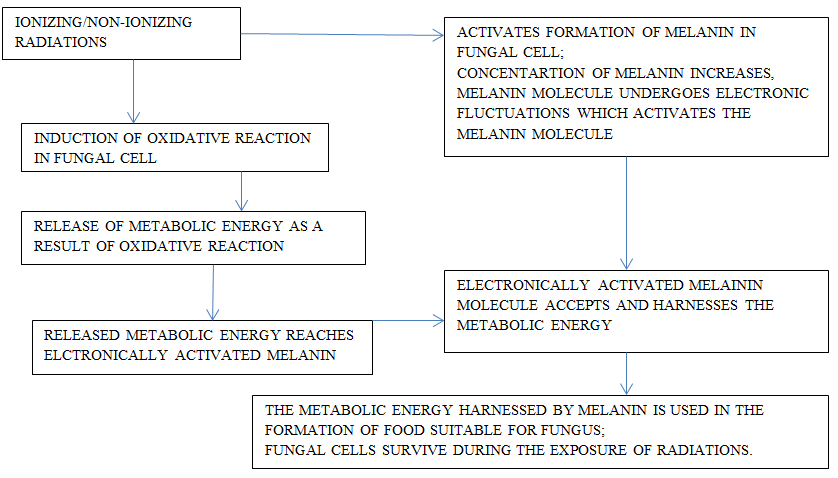
Responses of Yeast to the Non-Ionizing Radiations
The Saccharomyces cerevisiae on exposure to non- ionizing millimeter waves with range 85 to 105 GHz (as recommended by international commission on non-ionizing radiation protection) with non-thermal 20 mW/cm2 (within safety limits) and power density around 1.0 mW/cm2SAR for 5 to 6 hours result in 62% decline in their growth rate. This impact is specifically within 85 to 105 GHz and depends on the parameter like energy and the density. The growth profile of colony of wild type and delta rad52 (DNA repair gene) cells show no impacts of these irradiations. This reflects that probably the non-thermal effect does not induce permanent changes in the genetic make-up. The horn antenna (~1.0 mW/cm2 and compact waveguide (17.17 mW /cm2) emits radiations which causes complete stoppage of cell-division and this is due to non-thermal effect. This response is dose dependent. There is a possibility of its application in biomedical field [112]. The exposure of yeast cells to microwave radiations with 968MHz and power of 10dBm restricts collagenase and L-lactate dehydrogenase activities up to 26% while proliferation rate increases 15% [113]. Radiations from 905 MHz electromagnetic radiations closely match with the Global System for Mobile Communication (GSM) pulse modulation signals from mobile phone having specific absorption rate (SAR) OF 0.12W/kg. When S. cerevisiae strains FF18733 (wild-type), FF1481 (rad1mutant) and D7 (commonly used to detect reciprocal and nonreciprocal mitotic recombinations), do not induce changes in the colony growth rate statistically. The exposed strain FF1481 is relatively more sensitive to such radiations [114]. Yeast cells of Candida slooffii, Candida lipolytica, and Leucosporidium nivalis species can be readily cultured in laboratory and used for the appropriate investigations. For scanning electron microscopy these cells (in dry state) are fixed in buffered 3% glutaraldehyde for 1 hour, The coated dried cell of yeast coated with gold-are observed under scanning microscope using 10kV voltage. The cell wall of Candida species exhibits smooth surface but many protrusions are there on the surface of leucosporidium cells. There appears to be no relationship between the nature of the cell surface and thermophilic adaptive behavior of these spices of yeast [115]. Radiations having low frequency magnetic field of 60 Hz and 500 mT could not induce growth rate and frequency of mutation among wild and radiation sensitive mutant species (rad or rev) of S. cerevisiae. The radiations from static and extremely low frequency magnetic field are not potent enough to affect the growth of yeast cells and even kill the model organism [116].
The microwave irradiations of 42 GHz and 50 mW power (the nonionizing millimeter-waves, MMW) cause no significant changes in the rate of growth of yeast cells [117, 118]. The nonionizing millimeter-waves, MMW radiation having range 41.650 - 41.798 GHz for 4 h and 20 mW power result in frequency dependent sensitive effects like the growth rate elevates or declines in accordance to the frequency [119]. The exposure to radiation with 968 MHz and power of 17dBm for 7 hours induces higher rate of growth in yeast cells [120]. The non-ionizing radiation are likely to induce changes non-thermal effects, modulation in cell membrane and its impacts on cell signalling, biomolecular resonance effects, these changes are dose and power/energy dependence [113]. The exposure (5 to 6 hours) form non- ionizing millimeter-waves, (MMW) (85-105 GHz; W-band with minimum density ~1.0 mW/cm2 of 50cells/µl) causes 62% decline the growth rate in Saccharomyces cerevisiae yeast cells. The degree of the intensity of this effect varies in accordance to frequency, energy dose, and density of the model yeast cells. Such impacts are non-thermal in nature. The growth profile in case of wild type and delta rad52 (in which the repair mechanism of DNA is not present/or deleted) do not show any derogative effect on the respective stability of genomic set-up. The threshold parameters which favour inhibition of infection depend on the cell density specifically when exposed to the nonionizing millimeter- waves. The non-thermal impacts along with radiations for nonionizing millimeter-waves change the physical aspects of cell membrane resulting in disturbed cell signalling cascade and vesicular structure. These radiations absorb the water contents of yeast cell; this in turn changes the proteomics and conformational aspects of chromatin of the experimental cells. Modifications in genetic set-up can be directed to induce resistance for antifungal drugs in the test yeast cells. The super high frequency (SHF) 3 to 30 GHz bandwidth ensure the adaptability of the non-thermal techniques to modify the internal biochemical built up to overcome the burns developed during thermal effects as a result of conventional mode. Thus, there seems to be a possibility of retention of biochemical integrity of genetic set up of the experimental cells and induce a combined physical and bioelectromagnetic impact. This effect causes modifications in the proteome that is responsible for the behavior of the experimental yeast cells. Further, the most probable practical application of radiations from non-ionizing millimeter-waves, (MMW) involves the control of the targeted biological responses, and for this there is no need to modify activation of biochemical aspects or induce DNA damage to meet the fungal infection clinically [113, 121, 122, 123, 124].
Conclusion
The yeast organism has mostly similar cell organization and cell organelles to most of the eukaryotic cells. Generally, yeast cells have dimensions 5x10 µm, and their morphology, molecular aspects are readily observed. These biological entities are found in soil, plants, animals, and insects. Fungi have ability to exploit radiations to synthesize their food and able to tolerate higher concentrations of sugars, salts, and acetic acid and maintain their growth and budding processes. Their traits of stress tolerance are the bases of their applications and enhance the efficacy of different biotechnological and food processes. Yeast cells exhibit an efficient propensity of resistance or tolerance, some of the extreme conditions like influence of preservatives and elevated osmotic ambient surrounding. The non-ionizing millimeter-waves modify the proteomes that regulate the behavior of the yeast cell. These radiations may be of practical importance during understanding the molecular mechanism in targeted cells without modifying the biochemical aspects and/or induction of DNA damage to investigate clinical aspects of fungal infection.
Acknowledgement
Author gratefully acknowledges the encouragement from Dr. P M Dongre, Head, and Department of Biophysics -University of Mumbai. Mumbai, India
Conflicts of Interest
Author declares no conflict of the interest.
Declaration of Funding
No funding required for this presentation.
References
-
Kurtzman CP, Mateo RQ, Kolecka A, Theelen B, Robert V, et al. (2015) Advances in yeast systematics and phylogeny and their use as predictors of biotechnologically important metabolic pathways. FEMS Yeast Research 15(6): 17.
-
Oca MD, Salem R, Kholif AZM, Monroy AE, Perez LS, et al. (2016) Yeast: Description and structure. In: Salem AZM (Eds.), Yeast additive and animal production, pub Bio Med Central Research, Tamil Nadu, India, pp: 4-13.
-
Pringle JR, Adams AE, Drubin DG, Haarer BK (1991) Immunofluorescence methods for yeast. Methods in Enzymology 194: 565-602.
-
Bloecher A, Tatchell K (2000) Dynamic localization of protein phosphatase type 1 in themitotic cell cycle of Saccharomyces cerevisiae. The Journal of cell biology 149(1): 125-140.
-
Feldmann H (2012) Yeast cell architecture and functions. Yeast-molecular and cell biology.
-
Davey HM, Kell DB (1996) Flow cytometry and cell sorting of heterogeneous microbial Populations: the importance of single-cell analyses. Microbiol Rev 60(4): 641-696.
-
Osumi M (2012) Visualization of yeast cells by electron microscopy. Journal of Electron Microscopy 61(6): 343-
-
Lipke PN, Kurjan J (1992) Sexual agglutination in budding yeasts: structure, function, and regulation of adhesion glycoproteins. Microbiol Rev 56(1): 180-194.
-
Stewart GG (2017) The Structure and Function of the Yeast Cell Wall, Plasma Membrane and Periplasm. In: Brewing and Distilling Yeasts. The Yeast Handbook, Cham, Springer, pp 55-75.
-
Fricker MD, Willmer CM (1987) Vanadate Sensitive ATPase and Phosphatase Activity in Guard Cell Protoplasts of Commelina. Journal of Experimental Botany 38(4): 642-648.
-
Lahir YK (2015) A dynamic component of tissues-extra cellular matrix: structural, functional and adaptive approach. Biochem Cell Arch 15(2): 331-347.
-
Lahir YK (2016) Some aspects of interactions between nanomaterials and the cytoskeleton of eukaryotic Cells. Advances in Clinical Toxicology 1(2): 1-10.
-
Mulholland J, Preuss D, Moon A, Wong A, Drubin D, et al. (1994) Ultrastructure of the yeast actin cytoskeleton and its association with the plasma membrane. J Cell Biol 125(2): 381-391.
-
Mishra M, Huang J, Balasubramanian MK (2014) The yeast actin cytoskeleton. FEMS Microbiological Reviews 38(2): 213-227.
-
Stearns T (1997) Motoring to the Finish: Kinesin and Dynein Work Together to Orient the Yeast Mitotic Spindle. Journal of Cell Biology 138(5): 957-960.
-
Moore JK, Cooper JA (2010) Coordinating mitosis with cell polarity: Molecular motors at the cell cortex. Seminars in Cell & Developmental Biology 21(3): 283- 289.
-
Mariki G, Nuske E, Leng W, Alberti S, Pigino G (2020) Reorganization of budding yeast cytoplasm upon energy depletion. Molecular Biology of cell2020 31(12): 1232- 1245.
-
D’Angelo MA, Hetzer MW (2008) Structure, dynamics and function of nuclear pore complexes. Trends in Cell Biology 18(10): 456-466.
-
Thiry M, Lafontaine DLJ (2005) Birth of a nucleolus: the evolution of nucleolar compartments. Trends Cell Biol 15(4): 194-199.
-
Carle GF, Olson MV (1985) An electrophoretic karyotype for yeast. Proceedings of National Academy of Science, USA, 82(11): 3756-3760.
-
West M, Zurek N, Hoenger A, Voeltz GK (2011) A 3D analysis of yeast ER structure reveals how ER domains are organized by membrane curvature. J Cell Biol 193(2): 333-346.
-
Austriaco N (2012) Endoplasmic reticulum involvement in yeast cell death. Front Oncol 2: 87.
-
Voeltz GK, Prinz WA, Shibata Y, Rist JM, Rapoport TA (2006) A class of membrane proteins shaping the tubular endoplasmic reticulum. Cell 124(3): 573-586.
-
Shibata Y, Shemesh T, Prinz WA, Palazzo AF, Kozlov MM, et al. (2010) Mechanisms determining the morphology of the peripheral ER. Cell 143(5): 774-788.
-
Carman GM, Henry SA (2007) Special issue: regulation of lipid metabolism in yeast. Biochimica Biophysica Acta 1771(3): 239-240.
-
Henry SA, Kohlwein SD, Carman GM (2012) Metabolism and regulation of glycerolipids in the yeast Saccharomyces cerevisiae. Genetics 190(2): 317-349.
-
Strayle J, Pozzan T, Rudolph HK (1999) Steady-state free Ca2+ in the yeast endoplasmic reticulum reaches only 10 microM and is mainly controlled by the secretory pathway pump pmr1. EMBO J 18(17): 4733-4743.
-
Strahl T, Hama H, DeWald DB, Thorner J (2005) Yeast phosphatidylinositol4-kinase, Pik1, has essential roles at the Golgi and in the nucleus. J Cell Biology 171(6): 967- 979.
-
Suda Y, Nakano A (2012) The Yeast Golgi Apparatus. Traffic the Moving Front of Biology 13(4): 505-510.
-
Strahl T, Thorner J (2007) Synthesis and function of membrane phosphoinositides in Budding yeast, Saccharomyces cerevisiae. Biochimica Biophysica Acta 1771(3): 353-404.
-
Mayinger P (2009) Regulation of Golgi function via phosphoinositide lipids. Seminars in Cell & Developmental Biology 20(7): 793-800.
-
Beck R, Ravet M, Wieland FT, Cassel D (2009) The COPI system: Molecular mechanisms and function. Federation of European Biochemical Societies Letters (FEBS Letters) 583(17): 2701-2709.
-
Hughes H, Stephens DJ (2008) Assembly, organization, and function of the COPII coat. Histochemistry and Cell Biology 129(2): 129-251.
-
Lee MC, Miller EA (2007) Molecular mechanisms of COPII vesicle formation. Seminars in Cell Development Biology 18(4): 424-434.
-
Sato K, Nakano A (2007) Mechanisms of COPII vesicle formation and protein sorting. Federation of European Biological Society letters (FEBS Lett) 581(11): 2076- 2982.
-
Fromme JC, Orci L, Schekman R (2008) Coordination of COPII vesicle trafficking by Sec23. Trends Cell Biol 2008 Jul 18(7): 330-336.
-
Nakano A, Brada D, Schekman R (1989) A Membrane Glycoprotein, Secl2p, Required for Protein transport from the endoplasmic reticulum to the Golgi apparatus in Yeast. The Journal of Cell Biology 107(3): 851-863.
-
Demmel L, Gravert M, Ercan E, Habermann B, Reichert MT, et al.(2008) The Clathrin adaptor Gga2p is a phosphatidylinositol 4-phosphate effector at the Golgi exit. Molecular Biology of the Cell 19(5): 1991-2002.
-
Fratti RA, Jun Y, Merz AJ, Margolis N, Wickner W (2004) Interdependent assembly of specific regulatory lipids and membrane fusion proteins into the vertex ring domain of docked vacuoles. Journal of Cell Biology 167(6): 1087-1098.
-
Knop M, Schiffer HH, Rupp S, Wolf DH (1993) Vacuolar/lysosomal proteolysis: proteases, substrates, mechanisms. Current Opinion in Cell Biology 5(6): 990- 996.
-
Lazarow PB, Fujiki Y (1985) Biogenesis of Peroxisomes. Annual Reviews of Cell Biology 1: 489-953.
-
Zhang JW, Luckey C, Lazarow PB (1993) Three Peroxisome Protein Packaging Pathways Suggested by Selective Permeabilization of Yeast Mutants Defective in Peroxisome Biogenesis. Molecular biology of the cell 4(12): 1351-1359.
-
Boekhout T, Robert V (2003) Yeasts in Food: Beneficial and Detrimental Aspects. Boca Raton, Woodhead Publishing, FL: CRC Press, Cambridge, pp: 488.
-
Blackburn CDW (2006) Food Spoilage: Micro-organism. Elsevier Science and Technology.
-
Martorell P, Stratford M, Steels H, Espinar MTF, Querol A (2007) Physiological Characterization of Spoilage Strains of Zygosaccharomyces Bailii and Zygosaccharomyces Rouxii Isolated From High Sugar Environments. Int J Food Microbiol 114(2): 234-242.
-
Tokuoka K (2008) Sugar‐ and Salt‐Tolerant Yeasts. Journal of Applied Microbiology 74(2): 101-110.
-
Zakey AS, Tucker GA, Daw ZY, Du C (2014) Marine Yeast Isolation and Industrial Application. FEMS Yeast Res 14(6): 813-825.
-
Levin RL, Ushiyama M, Cravalho EG (1979) Water Permeability of Yeast Cells at Sub-Zero Temperatures. J Membr Biol 46(2): 91-124.
-
Hittinger CT, Rokas A, Bai FY, Boekhout T, Gonçalves P, et al. (2015) Genomics and the Making Yeast Biodiversity. Curr Opin Genet Dev 35: 100-109.
-
Schindler D (2020) Genetic Engineering and Synthetic Genomics in Yeast to Understand Life and Boost Biotechnology. Bioengineering (Basel) 7(4): 137.
-
Ploessl D, Zhao Y, Cao M, Ghosh S, Lopez C, et al. (2022) A repackaged CRISPR platform increases homology- directed repair for yeast engineering. Nature Chemical Biology 18: 38-46.
-
Cohen PA, Travis JC, Keizers PHJ, Boyer FE, Venhuis BJ (2018) The stimulant higenamine in weight loss and sports supplements. Clin Toxicol 57 (2): 125-130.
-
Longo VD, Shadel GS, Kaeberlein M, Kennedy B (2012) Replicative and chronological aging in Saccharomyces cerevisiae. Cell Metab 16(1): 18-31.
-
Jimenez J, Bru S, Ribeiro M, Clotet J (2015) Live fast, die soon: cell cycle progression and lifespan in yeast cells. Microb Cell 2(3): 62-67.
-
Bitterman KJ, Medvedik O, Sinclair DA (2003) Longevity regulation in Saccharomyces cerevisiae: linking metabolism, genome stability, and heterochromatin. Microbiol Mol Biol Rev 67(3): 376-399.
-
Longo VD, Mittledorf J, Skulachev VP (2005) Programmed and altruistic aging. Nature Reviews Genetics 6: 866- 872.
-
Longo VD, Finch CF (2003) Evolutionary medicine from dwarf model systems to healthy centenarians? Science 299(5611): 1342-1346.
-
Masoro EJ (2009) Biochemical and molecular mechanism of aging: from model system to human longevity. Biochimica et Biophysica Acta (BBAD)- general subject 1790(10): 949-950.
-
Chin RM, Fu X, Pai MY, Vergens L, Huwang H, et al. (2014) The metabolite alpha-ketoglutarate extends lifespan by inhibiting ATP-synthase and TOR. Nature 510(7505): 397-401.
-
Arlia CA, Piano A, Leonov A, Svistkova V, Titorento VI (2014) Quasi-programmed aging of budding yeast a trade-off between programmed process of cell proliferation, differentiation, stress response, survival and death defines yeast lifespan. Cell Cycle 13(21): 3336-3349.
-
Pedruzzi I, Dubiulo F, Cameroni E, Wanke V, Rosen J, et al. (2003) TOR and PKA signaling pathways converge on protein kinase Rim15 to control in to entryin Go. Molecular cell 12(6): 1607-1613.
-
Wanke V, Pedruzzi I, Cameroni E, Dubouloz F, De VC (2005) Regulation of Go entry by the Pho 80-Pho 85 cyclin, CDK complex. EMBO J 24(24): 4271-4278.
-
Hinnen A, Hicks JB, Fink GR (1978) Transformation of yeast. Proc Natl Acad Sci, USA 75(4): 1929-1933.
-
Kasianowicz JJ, Brandin E, Branton D, Deamer DW (1996) Characterization of individual polynucleotide molecules using a membrane channel, Proc Natl Acad Sci, USA 93(24): 13770-13774.
-
Levene MJ, Korlach J, Turner SW, Foquet M, Graighead HG, et al. (2003) Zero-mode waveguide single molecule analysis at high concentration. Science 299: 682-686.
-
Check HE (2009) Genome sequencing: third generation, Nature 457: 768-769.
-
Snyder M, Gallagher JE (2009) Systems biology from a yeast omics perspective. FEBS Lett 583(24): 3895-3899.
-
Botstein D, Fink GR (2011) Yeast: an experimental organism for 21st century biology. Genetics 189: 695- 704.
-
Berger SL, Kouzarides T, Sheikhattar R,Shilatifard A (2009) An operational definition of epigenetics. Genes Dev 23: 781-783.
-
Gartenberg MR, Smith JS (2016) The Nuts and Bolts of Transcriptionally Silent Chromatin in Saccharomyces cerevisiae. Genetics 203(4): 1563-1599.
-
OKane CJ, Hyland EM (2019) Yeast epigenetics: the inheritance of histone modification states. Biosci Rep 39(5): BSR20182006.
-
Zemach A, McDaniel IE, Silva P, Zilberman D (2010) Genome-wide evolutionary analysis of eukaryotic DNA methylation. Science 328(5980): 916-919.
-
Perego P, Howell SB (1997) Molecular mechanisms controlling sensitivity to toxic metal ions in yeast. Toxicol and Appl Pharmacol 147(2): 312-318.
-
Hamadeh HK, Amin RP, Paules SR, Ashfari CA (2002) An overview of toxicogenomics. Curr Issues Mol Biol 4(2): 45-56.
-
North M, Vulpe CD (2010) Functional toxicogenomics: mechanism-central toxicology. Int J Mol Sci 11(12): 4796-4813.
-
Santos SCD, Teixeira MS, Cabrito TR, Correia IS (2012) Yeast toxicogenomics: genome-wide responses to chemical stresses with impact in environmental health, pharmacology and biotechnology. Frontiers in Genetics 3: 63.
-
Sato ER, Ostroffer GR (2008) Delivery of DNA by yeast. Bioconjugate Chemistry 10(4): 840-848.
-
Skalickova S, Baron M, Sochor J (2017) Nanoparticles Biosynthesized by Yeast: A Review of their application. Kvasny Prum 63(6): 290-292.
-
Zhou X, Zhang X, Han S, Dou Y, Liu M, et al. (2017) Yeast Microcapsule-Mediated Targeted Delivery of DiverseNanoparticles for Imaging and Therapy via the Oral Route. Nano lett 17(2): 1056-1064.
-
J Xu, Ma Q, Zhang Y, Fei Z, Sun Y (2022) yeast derived nanoparticles remodel the immunosuppressivemicroenvironment in tumor growth. Nature Communications 13(1): 1-15.
-
Gal J (2008) The discovery of biological enantioselectivity, Louis Pasteur and the fermentationof tartaric acid, 1857- A review and analysis 150 yr later. Chirality 20(1): 5-19.
-
Iravani S (2011) Green synthesis of metal nanoparticles using plants. Green Chemistry 13(10): 2638-2650.
-
Duan HH, Wang DS, Li YD (2015) Green chemistry for nanoparticle synthesis. Chemical Society Reviews 44(16): 5778-5792.
-
Kauzegaran VJ, Farhadi K (2017) Green synthesis of Sulphur Nanoparticles assisted by a herbal surfactant in aqueous solutions. Micro & Nano Letters 12(5): 329- 334.
-
Moghaddam AB, Fariden N, Moniri M, Tahir PM, Mohammad R, et al. (2015)Nanoparticles Biosynthesized by Fungi and Yeast: A Review of Their Preparation, Properties, and Medical Applications. Molecules 20(9): 16540-16565.
-
Feng H, Liu SY, Huang XB, Ren R, Zhou Y, et al. (2017) Green biosynthesis of CdS nanoparticles using yeast cells for fluorescence detection of nucleic acids and electrochemical detection of hydrogen peroxide. Int J Electrochem Sci 12(1): 618-628.
-
Kawshik M, Ashtputre S, Kharrazi S, Volter W, Urban J, et al. (2003)Extracellular synthesis of silver nanoparticles by a silver-tolerant yeast strain MKY3. Nanotechnology 14(1): 95-100.
-
Eugenio M, Muller N, Frases S, Paes RA, Muricio TRL, et al. (2016) Yeast derived biosynthesis of silver/silver chloride nanoparticles and their anti-proliferating activity against bacteria. RSC Adventures 6(12): 1-38.
-
Salunke BK, Sawant SS, Lee SI, Kim BS (2015) Comparative study of MnO2 nanoparticle synthesis by marine bacterium Saccharophagus degradans and yeast Saccharomyces cerevisiae. Appl Microbiol Biotechnol 99: 5419-5427.
-
Zhang X, Qu Y, Shen W, Wang J, Huijie I, et al. (2016) Biogenic Synthesis of gold nanoparticles by yeast Magnusiomyces ingens LH-F1 for catalytic reduction of nitro phenols. Colloids and Surfaces A: Physicochemical and Engineering Aspects 497: 280-285.
-
Besner S, Kabashin AV, Winnik FM, Meunier M (2008) Ultrafast laser based “green” synthesis of non-toxic nanoparticles in aqueous solutions. Applied Physics A 93: 955-959.
-
Li H, Chen Q, Zhao J, kkulal U (2015) Enhancing the antimicrobial activity of natural extraction using the synthetic ultra-small metal nanoparticles. Scientific Reports 5: 11033.
-
Sowani H, Mohite P, Munot H, Shouche YS, Bapat T, et al. (2017) Green synthesis of gold and silver nanoparticles by an actinomycete Gordoniaamicalis HS-11: Mechanistic aspects and biological application. Process Biochemistry. 51: 374-383.
-
Sharma H, Kumar K, Choudhary CK, Mishra PK, Vaidya B (2016) Development and characterization of metal oxide nanoparticles for the delivery of anticancer drug. Artif Cells Nanomed Biotechnol 44(2): 672-679.
-
Rai M, Deshmukh SD, Ingle AP, Gupta IR, Galdiero M, et al. (2016) Metal nanoparticles: The protective nanoshield against virus infection. Critical Reviews in Microbiology 42(1): 46-56.
-
Mandal D, Bolander ME, Mukhopadhyay D, Sarkar G, Mukherjee P (2006) The use of microorganisms for the formation of metal nanoparticles and their application. Applied Microbiology and Biotechnology 69: 485-492.
-
Goffeau A, Barrell BG, Bussey H, Davis RW, Dujon B, et al. (1996) Life with 6000 genes. Science 274(5287): 546-
-
Huang M, Bao J, Nelson J (2014) Biopharmaceutical protein production by Saccharomyces cervices: current state and future prospects. Pharmaceutical Bioprocessing 2(2):167-182.
-
Kim H, Yoo ST, Kang HA (2015) Yeast synthetic biology for the production of recombinant therapeutic proteins. FEMS Yeast Research 15(1):1-16.
-
Nelson J (2013) Production of biopharmaceutical proteins by yeast. Bioengineered 4(4): 207-211.
-
Awan AR, Blount BA, Bell DJ, Shaw WM, Ho JCH, et al. (2017) Biosynthesis of the antibiotic nonribosomal peptide penicillin in baker’s yeast. Nature Communications 8: 15202.
-
Wan JY, Phan CW, Abdullah NW, Apparao Y, Macredie I (2021) Yeast in biopharmaceutical science: Visualisation of research landscape and bibliometric analysis. Malaysian Journal of Pharmaceutical Sciences 19(1): 133-144.
-
Fidan O, Zhan J (2015) Recent Advances in Engineering Yeast for Pharmaceutical Protein Production. RSC Advances 5(105): 86665-86674.
-
Meehl MA, Stadheim TA (2014) Biopharmaceutical discovery and production in yeast. Current Opinion in Biotechnology 30: 120-127.
-
Patra P, Das M, Kundu P, Ghosh A (2021) Recent Advances in Systems and Synthetic Biology Approaches for Developing Novel Cell-Factories in Non-Conventional Yeasts. Biotechnology Advances 47: 107695.
-
Dadachova E, Bryan RA, Huang X, Moadel T, Schweitzer AD, et al. (2007) Ionizing Radiation Changes the Electronic Properties of Melanin and Enhances the Growth of Melanized Fungi. PLoS ONE 2(5): e457.
-
Bordie C (2007) Sensitive Cells: Stem Cells Take Cue from their Physical Surrounding. American Scientist 95(2): 123.
-
Samant MP, Lahir YK, Chitre AV, Kale A (2018) Review- Influence of Nanomaterials on Cellular Stress and Cellular Behavior. Advances in Clinical Toxicology 3(3): 000140.
-
Oyarce G, Arrepol GB, Trivino S, Nanco F, Hasbun R, et al. (2021) Effects if Wi-Fi Radio Frequencies on the Contents of Fatty Acids and Ergosterol, a Precursors for Vitamin-D, Vitro Culture of Serpula Himantiodes. J Chil Chem Soc 66(1): 5125-5229.
-
Barbora A, Rajput S, Komoshvili K, Levitan J, Yahalom A, et al. (2021) Non-Ionizing Millimeter Waves Non- Thermal Radiation of Saccharomyces cerevisiae-Insights and Interactions. Applied Science 11(14): 6635.
-
Alsuhaim HS, Vojisavlijevic V, Pirogova E (2013) Effects of Low Power Microwave Radiation on Biological Activity of Collagenase Enzyme and Growth Rate of S. Cerevisiae Yeast. Proc. SPIE 8923, Micro/Nano Materials, Devices, and Systems, 892357.
-
Vrhovac I, Reno Hrascan R, Franekic J (2010) Effect of 905 MHz Microwave Radiation on Colony Growth of the Yeast Saccharomyces Cerevisiae Strains FF18733, FF1481 and D7.Radiol Oncol 44(2): 131-134.
-
Castro RA, Barreira PHK, Teixeira PA, Sanches GF, Teixeira MM, et al. (2013) Differences in Cell Morphometry, Cell Wall Topography and Gp70 Expression Correlate with the Virulence of Sporothrix Brasiliensis Clinical Isolates. PloS one 8(10): e75656.
-
Shimizu K, Nakaoka Y, Yamamoto T (2000) Biological effect of non-ionizing radiations on Micro-organism.
-
Grundler W, Keilmann F, Frohlich H (1977) Resonant Growth Rate Response of Yeast Cells Irradiated by Weak Microwaves, Physics Letters A 62(6): 463-466.
-
Grundler W, Keilmann F, and Frohlich H (1977) Resonant Growth Rate Response of Yeast Cells Irradiated by Weak Microwaves. Physics Letters A 62(6): 463-466.
-
Grundler W (1992) Intensity- and Frequency- Dependent Effects of Microwaves on Cell Growth Rates. Journal of Electro analytical Chemistry 342(3): 361-365.
-
Furia L, Hill DW, and Gandhi OP (1986) Effect of Millimeter-Wave Irradiation on Growth of Saccharomyces cerevisiae. IEEE Trans Biomed Eng 33(11): 993-999.
-
Vojisavljevic V, Alsuhaim H S, Pirogova E (2016) Low power microwave exposures at 968MHz increase the growth rate of Breanomyces bruxellensis yeast cells. 2016 IEEE International Conference on Microwave and Millimeter Wave Technology (ICMMT), Beijing, pp: 1061-1063.
-
Miranda I, Rocha R, Santos MC, Mateus DD, Moura GR, et al. (2007) A Genetic Code Alteration is a Phenotype Diversity Generator in the Human Pathogen Candida albicans.PLoS ONE 2(10): e996.
-
Silva RM, Paredes JA, Moura GR, Manadas B, Lima- Costa T, et al.(2007) Critical roles for a genetic code alteration in the evolution of the genus Candida. EMBO J 26(21): 4555-4565.
-
Cowen LE, Steinbach WJ (2008) Stress, drugs, and evolution: the role of cellular signaling in fungal drug resistance, Eukaryote Cell 7(5): 747-764.
-
Gonzalez VD, Ortiz PAM, Hundeiker M, Bonifaz A (2013) Opportunistic yeast infections: Candidiasis, cryptococcosis, trichosporonosis and geotrichosis, J Dtsch Dermatology Ges 11(5): 381-394.
-
ICNIRP (2020) Principles for nonionizing radiation protection. International Commission on non-ionizing radiation protection (ICNIRP). Health Physics 118(5): 477-482.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells