Exposure of Zebrafish (Danio rerio) to Clioquinol Reveals Embryological and Developmental Toxicity
Clioquinol (CQ) belongs to the 8-hydroxyquinoline family of antimicrobial agents. In the 1970s, CQ was withdrawn from the market due to reported neurotoxicity in humans. Although oral use was discontinued, topical formulations for treating fungal and parasitic infections remained available. With the potential reintroduction of oral CQ formulations for novel indications, acomprehensive understanding of its toxicity is essential. Thus, this study aimed to investigate the embryonic developmental toxicity of CQ in zebrafish. Embryos were exposed to concentrations of 0.1, 0.5, 1.0, 1.5, and 2.0 μg/ml of CQ for 168 hours, during which physiological, morphological, and behavioral parameters were assessed. Results indicate that CQ exhibited toxicity, causing 100% mortality within the first 48 hours at concentrations of 1.5 and 2.0 μg/ml. Additionally, a delay in hatching rate was observed after 72 hours of exposure to 1 μg/ml, and morphological abnormalities were evident at lower concentrations. Furthermore, exploratory behavior was evaluated in 7-day-old post-fertilization larvae exposed to the lowest CQ concentration, where severe morphological alterations were not bserved. Exposure to CQ at 0.1 μg/ml led to altered swimming patterns, suggesting possible locomotor damage induced by CQ exposure. In conclusion, this study is the first to demonstrate the potential embryotoxicity of CQ in zebrafish, highlighting the importance of understanding the toxicity profile before considering its reintroduction for oral use.
Introduction
Clioquinol, 5-chloro-7-iodo-quinolin-8-ol (CQ), with a molecular weight of 305.499 g/mol, is a halogenated derivative of 8-hydroxyquinoline and belongs to the class of chemotherapeutic agents known as antimicrobial agents. These compounds are utilized to prevent infections and inhibit the proliferation of microorganisms [1]. Initially introduced as an antiseptic in 1934, CQ was later marketed as an antibiotic for treating intestinal diseases, including chronic non-specific diarrhea and traveler’s diarrhea [2]. Subsequently, in the 1950s and 1970s, it was employed as an oral anti-parasitic agent against intestinal amebiasis, although its exact antimicrobial mechanism was not yet understood [1, 2, 3]. However, in the 1970s, oral CQ was withdrawn from the market due to reported neurotoxicity in Japanese patients and its association with a neurodegenerative syndrome called subacute myelo-optic neuropathy (SMON) [3, 4]. Despite this withdrawal, topical formulations of CQ for treating fungal and parasitic infections continued to be available.
Currently, CQ is commercially available as a topical antibiotic under the trade name Vioform® [5]. Beyond its antimicrobial applications, CQ and some analogues have demonstrated promising effects in treating non-infectious diseases such as Alzheimer’s disease [6] and Parkinson’s disease [7, 8]. Despite the efficacy observed in certain diseases, the precise neuroprotective mechanisms of in vivo CQ remain elusive. Considering the extensive exploration of therapeutic research involving CQ, renewed attention has been directed towards its systemic use. Consequently, with the potential reintroduction of oral formulations for new indications, a comprehensive understanding of the toxicology, biological characteristics, and toxicological evaluations of both CQ and its analogues is imperative. Moreover, while the antifungal activity of CQ is established, there is still a notable lack of consistent studies elucidating its mode of action and the extent of its toxicity, warranting further investigation.
Zebrafish (Danio rerio) has emerged as robust model for drug discovery and biosafety studies due to their striking resemblance in organ development to mammals, including the nervous, digestive, reproductive, immune, excretory, and cardiovascular systems [9]. The exceptional embryology and amenability to genetic manipulation make them one of the most crucial vertebrate organisms for biological research. This animal model offers numerous advantages throughout all stages of development, facilitating investigations into the pharmacological potential of novel drugs, toxicology of environmental contaminants, embryology, behavioral biology, and more [10, 11, 12, 13]. A key advantage lies in the transparency of zebrafish embryos, which allows for visualization of their development and assessment of various endpoints such as mortality, heart rate, somite formation, and hatching rate [14, 15].
Zebrafish has also emerged as a model system for studying behavior While many studies have focused on the behavior of adult zebrafish, certain behaviors manifest early and can be observed during the first week of development [11]. Understanding the behavioral profile is particularly valuable for modeling the effects on brain activity Hoffman EJ, et al. [16] which can be influenced by exposure to drugs and toxic substances [17]. Although traditional models have been effective for studying drug action, there is a growing interest in developing methods for large-scale studies using alternative models such as zebrafish [18].
The present study aims to investigate the potential embryotoxicity and behavioral alterations induced by embryonic exposure to Clioquinol (CQ) during zebrafish (Danio rerio) development. Specifically, we seek to assess the impact of various CQ concentrations on mortality, heart rate, somite formation, hatching rate, and locomotor behavior during the larval stage. By comprehensively examining these endpoints, we aim to gain insight into the potential neurotoxic effects of CQ and shed light on its safety profile, which is vital for considering its reintroduction as an oral drug for novel therapeutic indications.
Material and Methods
Zebrafish (Danio rerio) Husbandry and Egg Collection
The zebrafish husbandry adhered to strict ethical guidelines and received approval from the institution’s ethics committee (protocol 003-2016). Wild-type adult zebrafish were procured from a local supplier and housed in a Zebtec® recirculation system, ensuring optimal living conditions. The water parameters were meticulously maintained at pH 7.2 ± 0.5, 400 ± 50 μS conductivity, and a temperature of 28 ± 1 °C. The zebrafish were exposed to a photoperiod of 14 hours light and 10 hours dark to mimic their natural environment. They were nourished with a combination of commercial flocked feed and live food (Artemia salina) provided four times daily, following established protocols [19]. For breeding purposes, male and female zebrafish were paired overnight at a 2:1 ratio, and the reproduction process was induced the next morning. Prior to exposure, fertilized eggs resulting from natural crosses were carefully examined under a 2 hpf (hours post-fertilization) stereomicroscope, and only embryos at the normal zygote stage were selected. After successful reproduction, the selected embryos were maintained and housed in a controlled incubator at a constant temperature of 28 °C until exposure and throughout the course of the experiment.
Treatments
CQ (CAS Number: 130-26-7) was procured from Sigma Aldrich. To prepare the stock solution, CQ was dissolved in dimethylsulfoxide (DMSO) and further diluted in Zebtec® water, ensuring a maximum concentration of 0.005% DMSO in the subsequent exposures. Zebrafish embryos at 3 hours post-fertilization (hpf) were randomly distributed into 24- well plates and exposed to different concentrations of CQ, namely 0, 0.1, 0.5, 1, 1.5, and 2 μg/ml, until reaching 168 hpf. The CQ concentration curve was based on preliminary pilot experiments (data not shown). Each well contained 2.5 ml of the CQ solution and housed 5 embryos. The experimental timeline of CQ exposure and subsequent assessments is depicted in (Figure 1). The assays were conducted following the OECD guidelines for the testing of chemicals 236 – Fish Embryo Acute Toxicity (FET) Test [20], with minor modifications to suit the experimental requirements.
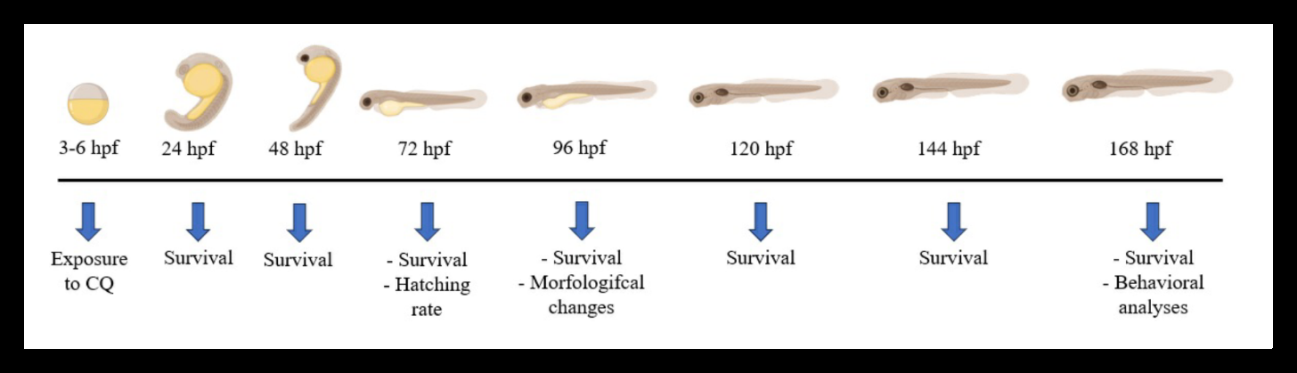
Survival Curve, Hatching Rate and Morphology
During the entire exposure period, embryos exposed to CQ were subject to daily monitoring, and their survival curve was analyzed based on indicators of unviability, such as egg coagulation and the absence of a heartbeat. To ensure the acceptability of the assay, the control group’s survival rate was maintained at ≥ 90% using dilution water, following OECD guidelines [20]. At 72 hours post-fertilization (hpf), the hatching rate was evaluated, and the percentage of viable individuals was quantified. At 96 hpf, when the embryos initiate their swimming process, a 3% methyl cellulose solution was applied to reduce movement for morphological analysis. Parameters, including body size, eye area, pericardial edema area, yolk sac edema area, and body axis distortion, were carefully evaluated, adhering to established protocols [19]. Images were acquired using a Carl Zeiss Stemi 2000-C stereomicroscope coupled with HD Lite 1080 P and 1.25× zoom and subsequently analyzed using Image J software. The biomarker results were utilized to calculate the “integrated biomarker response” (IBR) Beliaeff B, et al. [21], representing the ratio of morphological changes observed among different groups. This comprehensive approach allows for a thorough assessment of CQ-induced effects on zebrafish development and provides valuable insights into its potential embryotoxicity.
Behavioral Analysis
The exploratory behavior of the larvae was conducted at 7 days post-fertilization (dpf) following the method outlined by Colwill RM [17]. Only larvae that exhibited resilience throughout the entire exposure period and did not manifest severe morphological changes were included in the behavioral tests. Each larva was placed individually in a well of a 24-well cell culture plate, containing 2 mL of water per well, to assess exploratory behavior. Various behavioral parameters were evaluated, including the total distance traveled, absolute body turn angle, maximum speed, and immobile episodes of each animal. Immobility episodes were identified by the complete cessation of movement for a duration of two seconds. A 60-second habituation period was allowed before the sessions were filmed and recorded for a total of five minutes. For subsequent analysis, ANY- Maze software (Stoelting Co., Wood Dale, IL, USA) was employed to track and record the swimming activity of the animals at a rate of 15 positions per second. This rigorous behavioral assessment provides valuable data on potential locomotor alterations induced by CQ exposure, contributing to a comprehensive understanding of its effects on zebrafish development.
Statistical Analysis
The collected data underwent normality tests using D’Agostino & Pearson, Shapiro-Wilk, and Kolmogorov- Smirnov. Results were presented as mean ± standard error (S.E.M.). For non-parametric data, the Mann-Whitney test was utilized, while parametric data were analyzed using the One-Way Analysis of Variance (ANOVA) followed by the post- Tukey test. Statistical significances were considered at p ≤ 0.05.
Results
Exposure to CQ Increases Mortality and Affects the Embryos Eclosion
In the preliminary assessment of CQ toxicity on zebrafish embryonic development, we investigated common developmental phenotypes, including survival and hatching rates. The results revealed that lethality exhibited a dose and time-dependent pattern. Specifically, concentrations of 1.5 μg/ml and 2.0 μg/ml of CQ induced 100% mortality within a maximum of 48 hours of treatment (Figure 2). Additionally, the hatching process was notably impaired in embryos exposed to CQ at 1.0 μg/ml (Figure 2B). However, embryos subjected to concentrations of 1.5 μg/ml and 2.0 μg/ml did not survive up to 72 hours post-fertilization (hpf), precluding the evaluation of their hatching rates. Due to the observed high toxicity, concentrations of 1.5 μg/ml and 2.0 μg/ml were subsequently excluded from the study. These findings indicate a significant impact of CQ on zebrafish embryonic
Figure 2: Survival and hatching rate of embryos treated with different concentrations of CQ; a) Survival curve generated by software package. The colors for the different concentrations exposed to the Control and Clioquinol groups are distributed as indicated in the legend; b) Evaluation of the hatching rate of exposed embryos at 72 hours. Data were submitted to the One- way ANOVA statistical test. Data are expressed as mean ± SEM.
Morphological Abnormalities Induced by Clioquinol
To assess the toxicity of CQ on zebrafish morphogenesis, we recorded and plotted four major types of abnormalities at 4 days post-fertilization (dpf), including body size, eye size, pericardial edema, and yolk sac edema. Notably, all tested concentrations of CQ induced morphological changes, with the highest concentrations leading to severe malformations (Figure 3). The exposed groups exhibited a significant B survival and hatching, emphasizing the importance of further investigating the remaining concentrations for potential adverse effects on developmental parameters.
120 Hatching rate at 72hpf (%)
*
reduction in both body size (Figure 3C) and eye area (Figure 3D) (p < 0.0001). Moreover, there was a notable increase in the area of pericardial edema (Figure 3E) at the concentration of 1.0 μg/ml, and an increase in the area of yolk sac edema (Figure 3F) was significant at concentrations 0.1, 0.5, and 1.0 μg/ml (p < 0.0001). Furthermore, (Figures 3A & 3B) shows the IBR values for each endpoint of morphological change in each exposed group relative to the control. These results demonstrated a dose-dependent relationship between CQ concentration and the rate of morphological changes.

Figure 3: Effect of CQ on zebrafish morphogenesis. (A) Integrated biomarkers response analysis showing the overlapping occurrence of the notochord deformities with eye deformities, pericardial edema and yolk sac edema; (B) Representative images of morphological changes observed in 4 dpf zebrafish embryos at different concentrations of CQ; (C) body size, (D) eye size, (E) area of pericardial edema; (F) area of the yolk sac edema. Data are expressed as mean ± SEM. **** p <0.0001.
Clioquinol Causes Injury in Zebrafish Behavioral Parameters
Considering that exposure to CQ at a concentration of 0.1 μg/mL did not result in severe morphological abnormalities, such as notochord distortion, this specific concentration was chosen for behavioral analysis. Zebrafish exposed to 0.1 μg/mL of CQ displayed significant behavioral alterations compared to the control group. Specifically, the animals exposed to 0.1 μg/mL CQ exhibited a significant increase (p < 0.05) in the displacement distance traveled (Figure 4A) and a marked increase in the absolute body turn angle parameter (Figure 4B,4C&4D) p < 0.0001), indicating altered swimming patterns. Additionally, the immobile time was significantly increased (Figure 4E) (p < 0.01), indicating a prolonged period of immobility. Moreover, the maximum velocity of movement was significantly elevated (Figure 4F, p < 0.0001), suggesting hyperactivity in the exposed animals compared to the control group.
Interestingly, the time of permanence in both the center and periphery of the testing arena showed no significant differences when compared to the control group. These findings suggest that exposure to CQ at 0.1 μg/mL induces significant alterations in locomotor behavior in zebrafish larvae, without affecting their exploratory behavior in distinct areas of the testing arena.
0.25 Area of eye at 96hpf (µm2)
0.00 -0.05 1,0 µg/ml 0,5 µg/ml 0,1 µg/ml Control
Clioquinol
Area of yolk sac at 96hpf (µm2)
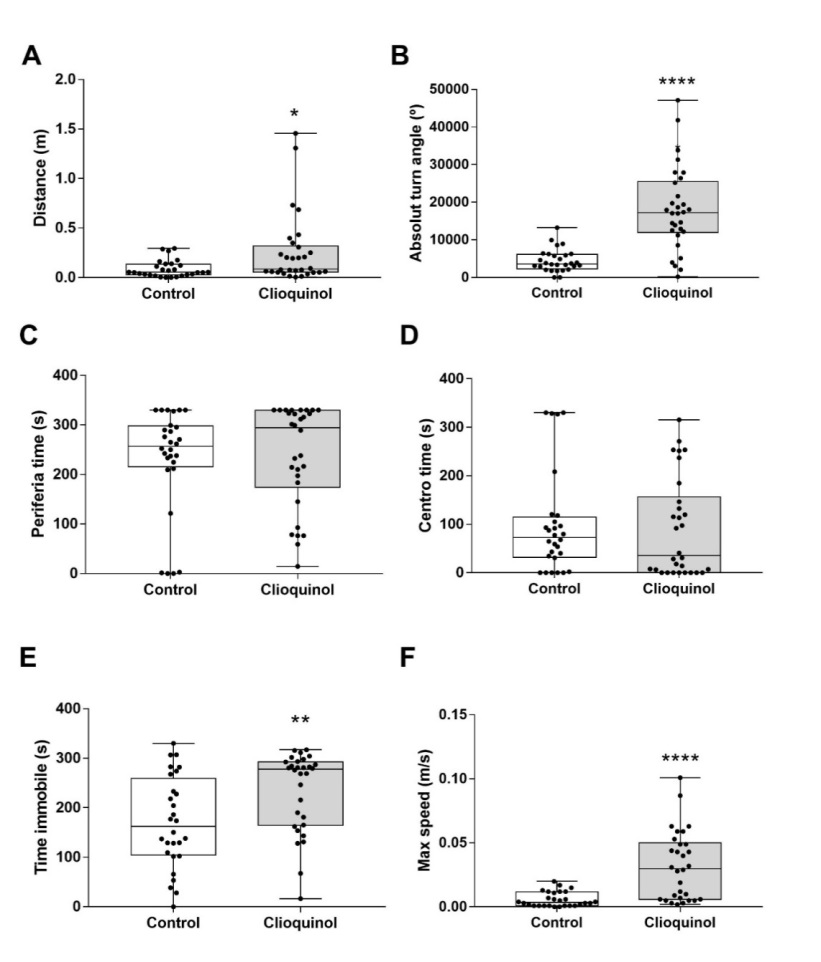
Figure 4: Exploratory behavior of zebrafish larvae. Distance (A), absolute turn angle (B), time at the periphery (C), time at the center (D), immobile time (E) and maximum speed attained (F) were evaluated at 7 dpf in control and 0.1 μg/ mL CQ treated larvae. Data were expressed as the mean ± standard error mean. * p <0.05, p <0.01, ** p <0.0001.
Discussion
This study investigated the toxic effects of CQ on zebrafish embryos, in terms of mortality, hatch rate, morphology, and behavior. The concentrations used were based on the levels of CQ found in commercially available formulations. A survival curve was generated at concentrations ranging from 0.1 to 2.0 μg/ml. At 48 hours, 100% of the embryos exposed to 1.5 and 2.0 μg/ml CQ had died, indicating the high toxicity of this compound to zebrafish embryos.
Hatching is an important developmental endpoint in fish and can be used as a powerful tool in developmental toxicology [10, 22, 23]. It is related to body movements, which are necessary to break the egg envelope. Loss of muscle capacity due to abnormal body formation can also delay hatching. Delays in hatching rate have been reported as a sign of stress response in zebrafish embryos [24]. Drugs and environmental toxicants can cause hatching inhibition and behavioral defects related to attenuated spontaneous muscle movements of the embryos [22]. In our study, we observed a significant delay in hatching rate after exposure to CQ at 1 µg/ ml, indicating toxicity towards this biological phenomenon.
Pericardial edema is a sensitive and specific indicator of cardiac toxicity. It is the accumulation of fluid in the pericardial sac, which is the sac that surrounds the heart. This can put pressure on the heart and prevent it from pumping blood effectively. A significant increase in pericardial edema can be a sign that a toxicant is having a negative impact on cardiac function [25]. Our study observed a significant increase in pericardial edema at a concentration of 1.0 μg/ml of CQ, suggesting that the compound has potential cardiotoxic properties (Figure 3E). Additionally, all tested concentrations of CQ resulted in a notable decrease in both yolk sac and body size. Deformities in eye size and notochord were also evident. Intriguingly, some or all of these deformities occurred simultaneously in the zebrafish larvae, indicating that CQ has a comprehensive impact on multiple developmental processes. The observed toxicological signs in zebrafish larvae align with previous studies reported in the literature [26, 27]. These signs typically include edema (cranial, pericardial, and abdominal), hemorrhages, tail curvature, and deformities in the head and spinal cord (marked scoliosis). The similarity of our findings to established toxicological markers reinforces the validity of our study and provides strong evidence for the potential toxicity of CQ in zebrafish embryonic development. Understanding these adverse effects is essential for assessing the safety and risks associated with CQ and its potential therapeutic applications.
Locomotor behaviors are essential for zebrafish throughout their lifespan, playing a key role in feeding, social, and defensive activities [17]. In addition, studies recommend using 6- or 7-day post-fertilization (dpf) larvae to examine the effects of drug treatments on locomotor activity, highlighting the importance of considering maturational and experimental effects on zebrafish larval behavior [11]. Behavioral changes are also known to reflect how an animal copes with its environment and are the first line of defense when the animal is exposed to harmful stimuli [28]. In this study, it was demonstrated that the concentration of 0.1 μg/ ml of CQ significantly increased the distance traveled, the maximum speed, and the absolute turn angle of the larvae compared to the control group. Differences in rotation angle may be directly related to changes in the larval swimming pattern [17]. Exposure to CQ also decreased the time the larvae remained immobile, and there was no significant difference in residence time in the peripheral and central areas. Taken together, our results indicate that embryonic exposure to CQ increased locomotor behavior and did not impair non-motor behaviors.
The ability of CQ to chelate and redistribute metals plays an important role in its neurotoxic effects, which depend on Zn, Cu, and Fe homeostasis [2]. These essential metals play key roles in embryonic, fetal, and postnatal development in higher eukaryotes. In this study, the concentrations tested were able to impair survival, morphological, and behavioral aspects of early developmental stages of zebrafish. Additionally, CQ may act as a pro-oxidant, leading to redox imbalance. This could contribute to the damaging effects of CQ exposure during developmental stages, as the sensitivity of developing organisms to environmental contaminants has been associated with cellular redox imbalance, which can impair normal development (Dennery, 2007). This mechanism of toxicity could be suggested in larval models.
Conclusion
In conclusion, our results provide a better understanding of the potential toxic effects of CQ on zebrafish embryogenesis for the first time. However, further studies are needed to clarify the biochemical mechanisms involved in order to have a comprehensive understanding of the impacts of CQ on the embryonic development of zebrafish.
Conflict of Interest
Authors declare no conflict of interest.
Data Availability
No data set was used in this manuscript.
Acknowledgements
Authors acknowledge Brazilian agencies for financial
support. CAPES code 001, FAPERGS (PRONUPEQ, 23828.388.17137.16092016) and CNPq (405426/2021-6, 406442/2023-3). JLF and TP hold CNPq research fellowships (314565/2020-5 and 307316/2022-0, respectively).
References
-
Ghalkhani M, Fernandes IPG, Oliveira SCB, Shahrokhian S, Oliveira-Brett AM (2011) Anodic behavior of clioquinol at a glassy carbon electrode. Bioelectrochemistry 80 (2): 175-181.
-
Bareggi SR, Cornelli U (2012) Clioquinol: Review of its mechanisms of action and clinical uses in neurodegenerative Disorders. CNS Neuroscience and Therapeutics 18 (1): 41-46.
-
Yassin MS, Ekblom J, Xilinas M, Gottfries CG, Oreland L (2000) Changes in uptake of vitamin B12and trace metals in brains of mice treated with clioquinol. Journal of the Neurological Sciences 173 (1): 40-44.
-
Mao X, Schimmer AD (2008) The toxicology of Clioquinol. Toxicol Lett 182(1-3): 1-6.
-
Wehbe M, Malhotra AK, Anantha M, Lo C, Dragowska WH, et al. (2018) Development of a copper-clioquinol formulation suitable for intravenous use. Drug Delivery and Translational Research 8(1): 239-251.
-
Hung LW, Barnham KJ (2012) Modulating metals as a therapeutic strategy for Alzheimer’s disease. Future Medicinal Chemistry 4(8): 955-969.
-
Finkelstein DI, Hare DJ, Billings JL, Sedjahtera A, Nurjono M, et al. (2016) Clioquinol Improves Cognitive, Motor Function, and Microanatomy of the Alpha-Synuclein hA53T Transgenic Mice. ACS Chemical Neuroscience 7(1): 119-129.
-
Lei P, Ayton S, Appukuttan AT, Volitakis I, Adlard PA, et al. (2015) Clioquinol rescues Parkinsonism and dementia phenotypes of the tau knockout mouse. Neurobiology of Disease 81: 168-175.
-
Bambino K, Chu J (2017) Zebrafish in Toxicology and Environmental Health. Curr Top Dev Biol 124: 331-367.
-
Costa-Silva DG da, Leandro LP, Lopes AR, Schimith LE, Nunes MEM, et al. (2018) N-acetylcysteine inhibits Mancozeb-induced impairments to the normal development of zebrafish embryos. Neurotoxicology and Teratology 68: 1-12.
-
Creton RMC, Creton R (2011) Imaging escape and avoidance behavior in zebrafish larvae Ruth. Neurosciense 22(1): 63-73.
-
Dammski AP, Müller BR, Gaya C, Regonato D (2011) Manual de criação em biotério. Zebrafish.
-
Li H, Cao F, Zhao F, Yang Y, Teng M, et al. (2018) Developmental toxicity, oxidative stress and immunotoxicity induced by three strobilurins (pyraclostrobin, trifloxystrobin and picoxystrobin) in zebrafish embryos. Chemosphere 207: 781-790.
-
Dennis P, Silva C (2018) Neurotoxicidade E Embriotoxicidade Induzidas Pelos Agentes Tóxicos Em Peixe-Zebra.
-
Kimmel C, Ballard W, Kimmel S, Ullman B, Schilling FT (1995) Stages of Embryonic Development of the Zebrafish. Dev Dyn 203: 255-310.
-
Hoffman EJ, Turner KJ, Fernandez JM, Cifuentes D, Ghosh M, et al. (2016) Estrogens Suppress a Behavioral Phenotype in Zebrafish Mutants of the Autism Risk Gene, CNTNAP2. Neuron 89(4): 725-733.
-
Colwill RM, CretonR (2011) Locomotor behaviors in zebrafish (Danio rerio) larvae. Behavioural Processes 86(2): 222-229.
-
Bruni G, Lakhani P, Kokel D (2014) Discovering novel neuroactive drugs through high-throughput behavior- based chemical screening in the zebrafish. Frontiers in Pharmacology pp: 1-14.
-
Westerfield M (2000) The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio).
-
OECD (2013) Organization for Economic Co-Operation and Development.
-
Beliaeff B, Burgeot T (2002) Integrated Biomarker Response: A Useful Tool For Ecological Risk Assessment. Environmental Toxicology Chemistry 21(6): 1316-1322.
-
Haendel MA, Tilton F, Bailey GS, Tanguay RL (2004) Developmental toxicity of the dithiocarbamate pesticide sodium metam in Zebrafish. Toxicological Sciences 81(2): 390-400.
-
Hallare AV, Kosmehl T, Schulze T, Hollert H, Köhler HR, et al. (2005) Assessing contamination levels of Laguna Lake sediments (Philippines) using a contact assay with zebrafish (Danio rerio) embryos. Science of the Total Environment 347(1-3): 254-271.
-
Dennery PA (2007) Effects of oxidative stress on embryonic development. Birth Defects Research Part C - Embryo Today 81(3): 155-162.
-
Sun M, Ding R, Ma Y, Sun Q, Ren X, et al. (2021) Cardiovascular toxicity assessment of polyethylene nanoplastics on developing zebrafish embryos. Chemosphere 282: 131124.
-
Chhetri J, Dilek J, Davies N, Jacobson G, Dallmann R, et al. (2022) NQO1 protects against clioquinol toxicity. Front Pharmacol 13.
-
Henry SP, Giclas PC, Leeds J, Pangburn M, Auletta C, et al. (1997) Activation of the alternative pathway of complement by a phosphorothioate oligonucleotide: potential mechanism of action. J Pharmacol Exp Ther 281(2): 810-816.
-
Scherer PE, Manning-Kriegt UC, Jen6 P, Schatzt G, Biocenter MH (1992) Identification of a 45-kDa protein at the protein import site of the yeast mitochondrial inner membrane. Proc Natl Acad Sci USA 89(24): 11930- 11934.
- Evaluation of Skin Aging Preventive Effects of Cherry Blossom Petal Extracts Through Antioxidant and Anti-Glycation Activities
- Is Cell Death Responsible for False Positive Results of In Vivo Comet Assay?
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism