Methylcobalamin Acts as a Synergistic Effector of Cadmium Chloride (Cdcl2 ) and Enhances Cadmium Induced Stress in Human A549 Cells through Methylation of MT2A Promoter
Methylcobalamin (MeB12) a vitamin B12 analogue is used in the treatment of several neurological disorders was used in this study to check for its role in cadmium toxicity. Cadmium is a highly toxic heavy metal can cause considerable damage to airway epithelium which can lead to many pulmonary disorders and even lung cancer. Human A549 cells were grown in medium supplemented with MeB12. for 48 hours, followed by 24 hours exposure to increasing concentrations of CdCl2 . MeB12 enhanced cadmium induced cells death. Isobolographic analysis revealed a synergistic effect of these two compounds. Studies on the revealed while MT1A promoter is already heavily methylated in A549 cell line, MT2A appeared to be methylated only in the presence of CdCl2 and MeB12. Our results suggest that the epigenetic modification of MT2A gene may be one of the contributing factors towards the enhanced cadmium sensitivity in A549 cells in the presence of MeB12.
Introduction
Methylcobalamin (MeB12) is a form of vitamin B12. It is the cofactor of the enzyme methionine synthase which catalyzes the regeneration of methionine from homocysteine. It transfers methyl group from methyl tetrahydrofolate to homocysteine, releasing 4 folate groups along with the formation of a methionine molecule [1].
Like every other vitamin, MeB12 is an antioxidant and is used for the treatment of neurological disorders like diabetic neuropathy [2], peripheral neuropathy [3] and its therapeutic effects have been reported in various other diseases like HIV [4] , cancer [5] etc. Studies on the protective effects of MeB12 against neurotoxicity in rat retinal cells, on administration of chronic MeB12 dosage (for a period of 48-96 hours) have shown positive results [3]. A double blind study on the effects of MeB12 on the compound muscle action potential amplitudes in ALS (Amyotrophic lateral sclerosis) patients showed that, at high dosages, it improved the neuronal functioning in those patients [6]. Physiological level of MeB12 in human body is equal to that of vitamin B12 and it is shown to be effective in treating diseases resulting from deficiency of Vitamin B12, like pernicious anaemia [7].
Heavy metals are potent toxicants. Biologically, cadmium not only has any constructive purpose, but also is detrimental to almost every organ in our body. It is toxic, non-essential and is classified as a human carcinogen by the National Toxicology Programme, National Institute of Health, US Department of health and human services [8, 9]. It is a stable metal element that occurs naturally in earth’s crust at an average concentration of 0.1 to 0.5 parts per million (ppm) and present almost everywhere in our environment-air, water and soil [10]. In humans, food is generally the main source of cadmium, for plants absorb cadmium from the soil. Smoking is another important source of cadmium exposure. Cigarette smoke contains about 1-2µg of cadmium per cigarette, about 10% of which is absorbed by the lung epithelium [11]. Its biological half-life in human body is between 17-30 years [12]. As a result smokers have comparatively higher level of cadmium in their blood than non-smokers [9, 11, 13].
Vitamins are known to be effective against metal toxicity [14, 15]. For e.g:- apart from chelation therapy, diets rich in vitamins and minerals are prescribed for people who are at risk of cadmium and lead toxicity [16]. There are similar reports available on the positive effects of vitamin B12 and MeB12 on the recovery of metal induced toxicity [17, 18, 19]. Reports show that MeB12 helps our body to eliminate the toxic heavy metal arsenic by methylating it along with selenium and methionine [20, 21].
Metallothioneins are metal binding cysteine rich membrane bound proteins. MT1 and MT2 are two major isoforms of this protein [22]. Expression of these isoforms is regulated by the transcription factor MTF1 (Metal- responsive transcription factor1) by binding to the metal response elements (MRE) in their promoter region [5, 22]. Studies have shown that both MT1 and MT2 expressions are induced in response to cadmium and these two isoforms play protective roles against cadmium induced cytotoxicity [5, 23]. On cadmium entry, activation of the transcription factor MTF1 and its subsequent translocation to the nucleus occurs. In the nucleus, MTF1 binds and activates the expression of genes having metal response elements (MRE) in their promoter region, like MT1 and MT2 leading to their expression and subsequent elimination of cadmium from the cell, by these two protein isoforms. A study in mice has shown that, disruption of these two genes makes them susceptible to cadmium [23].
Current study was intended to understand the effects of MeB12 on cadmium induced stress in human A549 cell line, a human non-small cell lung cancer cell line which was proved to be a suitable model for studying pulmonary toxicity [24, 25].
Materials and Methods
In Vitro Culture of A-549 Cells
The cell line was obtained from The National Centre for Cell Science (NCCS), Pune, India. A549 cells were maintained in Dulbecco’s Modified Eagle’s Medium (GIBCO) containing 10% Fetal Bovine Serum (SIGMA) supplemented with 0.1% antibiotic antimycotic solution (SIGMA) in 5% humidified CO2 condition at 370C.
Meb12 and Cdcl2 Treatment
A549 Cells at a seeding density of 2X104 cells/ml were grown in media containing 50 and 100 µg/ml MeB12 (SIGMA) while maintaining one set of cells in regular medium. After 48 hours, cells in the 3 experimental sets were exposed to different concentrations of CdCl2 (HIMEDIA) in the following order non treated control (no CdCl2), 2.5, 5 and 10µg/ml CdCl2. After 24 hours of treatment cells were harvested and subjected to various studies.
Cell Counting
After 24 hours of specific treatment, the total viable cell count was determined by trypan blue dye exclusion method using a Haemocytometer (Tiefe Depth Profondeur, Marienfeld, Germany).
Microscopic Analysis
Following the cadmium exposure for 24 hours, all the 3 experimental sets were observed under inverted optical microscope (Nikon eclipse TS 100) and the pictures were taken using a compact digital camera (Nikon COOLPIX L22) attached to the microscope.
Cytotoxicity Assay
For assessing the cytotoxic effect of cadmium, A549 cells were seeded at a density of 2x104 cells per well in a 24 well plate (both in regular and MeB12 supplemented media), allowed to grow for 48 hours and then exposed to different doses of CdCl2. After 24 hours of exposure, resazurin reagent (SIGMA) was added at a concentration of 10% in each well. Readings were taken spectrophotometrically at a wave length of 600nm and a reference wave length of 690nm.
Fluorescent Staining with Hoechst
Chromatin condensation was detected by nuclear staining with Hoechst 33342. Hoechst 33342 (1 µg/ml) was directly added to the media and stained nuclei were visualized under Nikon eclipse TS 100 fluorescence microscope at 40×magnification with an excitation wavelength of 355- 366 nm and an emission wavelength of 465-480 nm. Three independent replicates were used. In this way, apoptotic cells were stained bright blue because of their chromatin condensation, while normal cells were stained blue.
RNA Extraction and Reverse Transcriptase-PCR Analysis
Total cellular RNA was extracted using TRIzol® LS Reagent, as per the manufacturer’s (Invitrogen, USA) protocol. First strand cDNA was synthesised from RNA samples (2 µg) using the cDNA synthesis kit for Reverse Transcriptase-PCR (Tetro cDNA synthesis kit) from Bioline, USA, as per specifications provided in the kit. PCR amplification of different genes were carried out using their respective primers designed using NCBI Primer BLAST. For semi quantitative analysis of gene expression using reverse transcriptase PCR, in order to ensure that the amplicon is measured during the exponential phase of the reaction, equivalent amounts of cDNA was amplified over different number of PCR cycles for each gene. General PCR conditions were carried out for required cycles for each gene against their respective primers. Given below are the names of the genes along with their respective primer sequences and annealing temperatures- CDC 42 (500C) forward primer 5’ AGA GGA AAT ACG AGG GGT GGT 3’, reverse primer5’ CCT GAC TGG TCC CCA TGT TG 3’, RAC1 (600C) forward primer 5’GCC AAT GTT ATG GTA GAT GGA 3’ reverse primer 5’TTAAGA ACA CAT CTG TTT GCG 3’, RHOA (550C) forward primer 5’CCT TGT CTT GTG AAA CCT TG3’ reverse primer 5’ ACT GGT AGC AAG ATG ACT TC3’, HSP70 (500C) forward primer 5’ ACA CGA ATC CCT GCG GTA AA3’ reverse primer 5’AGC AGG CGA TAA GAT GGC AC3’, NFκB1 (500C) forward primer 5’ATC CAT ATT TGG GAA GGC CTG AA3’ reverse primer 5’GTA TGG GCC ATC TGT TGG CAG 3’, MT1 (550C) forward primer 5’ CCT GCA CCT GCA CTG GCT CCT G 3’ reverse primer 5’ TGG GCA CAC TTG GCA CAG CTC AT 3’, MT2 (500C) forward primer 5’ CAA AGG GGC GTC GGA CAA G 3’ reverse primer 5’ATA GCA AAC GGT CAC GGT CAG 3’, P53 (500C) forward primer 5’TGC TCA AGA CTG GCG CTA AA3’ reverse primer 5’CAA TCC AGG GAA GCG TGT CA3’, BCL2 (600C) forward primer 5’GAA CTG GGG GAG GAT TGT GG 3’ reverse primer 5’GGC AGG CAT GTT GAC TTCAC3’, BAX (700C) forward primer 5’GGC CCT TTT GCT TCA GGG TTT C3’ reverse primer 5’CAG TCG CTT CAG TGA CTC GG 3’, 18S rRNA (500C) forward primer 5’GTA ACC CGT TGA ACC CCA TT3’ reverse primer 5’ CCA TCC AAT CGG TAG TAG CG 3. PCR products were visualized on 1.2% agarose gel followed by ethidium bromide staining, using a gel documentation system (BIORAD, USA).
Methylation Specific PCR (MSP)
The MSP primers for MT1A and MT2A genes were designed using the online software tool, MethPrimer. Genomic DNA was extracted from control and treated A549 cells using a Genomic DNA isolation Kit (Bioline, USA). Equal amount of genomic DNA from each sample was subjected to bisulfite modification using Nucleo-pore® DNA Methylation Kit as per instructor’s manual. Methylation-specific PCR for the MT1A and MT2A genes was carried out with primers specific for the methylated and unmethylated promoter DNA sequences of the two genes (sequences are shown in Table 1). The reaction mixture was incubated at 95°C for 5minutes, followed by 30 cycles of 94°C for 30 seconds, annealing temperature for 30 seconds, 72°C for 1minute seconds, and a final extension at 72°C for 5 minutes. PCR products were visualized on 2% TAE agarose gels, by ethidium bromide staining.
| Gene Name | Primer Sequence | |
|---|---|---|
| MT1A | Forward M primer | ATA GGG ATA GGT AAG GCG ATA GC |
| Reverse M primer | CGA TCG ACG TAA TAC AAA ACG TA | |
| Forward U primer | TGG ATA GGG ATA GGT AAG GTGA TAG T | |
| Reverse U primer | CCC A ATC AAC ATA ATA CAA AAC ATA | |
| MT2A | Forward M primer | GAG TCG TAA GTG ATT TAG CGC |
| Reverse M primer | TTC GCT AAA ACT TAA AAA AAA CGT A | |
| Forward U primer | TGG AGT TGT AAG TGA TTT AGT GTG G | |
| Reverse U primer | TTC ACT AAA ACT TAA AAA AAA CAT A |
Isobologram Analysis
To see the type of interaction displayed by the combination of CdCl2 and MeB12, a classic isobolograhic technique was used. When two chemicals are mixed together, the resulting interaction between them can be additive, synergistic and antagonistic. When their combined effect is equal to that of the sum of their individual potencies, then the interaction is termed additive, when it is greater the interaction is synergistic and when it is lesser, antagonistic [26, 27]. Isobologram is a graph in Cartesian coordinates where each axis represents the concentrations of a particular chemical (here MeB12 in X axis and CdCl2 in Y axis). Isobole is a line that joins dose pairs that produce a fixed response.
Here we have selected half the maximum response, so the intercepts are individual IC50 values of MeB12 and CdCl2. In brief, A54 cells in 24-well plates were exposed in triplicate to increasing concentration of each chemical or both the chemicals using a constant ratio (CdCl2: MeB12=1:2) combination design for 48 hours. Cell viability after 48 hours was measured by reassuring assay. From the experimental values a dose matrix was constructed in excel and entered in chalice analyzer to construct the isobologram. The line of additivity follows the isobole while the Cartesian coordinates denoting synergy will be below the line of additivity and that representing antagonism will be above it [28].
Statistical Analysis
Data are expressed as the mean ± standard deviation. Statistical analysis of the results was done using Microsoft excel. Two- way ANOVA with multiple comparisons was used to test whether differences in the degree of cell viability on exposure to CdCl2 and MeB12 were more than by chance. One-way ANOVA with post- hoc test was used within dosage points where interaction was observed. Probabilities of p< 0.05 were regarded as statistically significant. The evaluation of synergy, construction of dose matrix and isobologram was done using Chalice Analyzer.
Results
Effect of Cdcl2 on the Morphology and Viability of Meb12 Treated A549 Cells
In cells grown in MeB12 containing media as well as regular media, cell death showed positive linear relation with cadmium concentration. The remaining viable cells had different morphology compared to their respective controls in all the three sets (Figures 1A & 1B). Surprisingly cell death by CdCl2 was relatively more prominent in A549 cells grown in MeB12 media which increased with the increase in MeB12 concentration (Figures 1B & 1C). The IC50 (The half maximal inhibitory concentration) values of CdCl2 in MeB12 sets (8.5±0.543 and 7.8 ±0.657 µg/ml of CdCl2) compared to regular medium set (9.2 ±0.841 µg/ml of CdCl2) (Table 2), was lower (Figures 2 & 3) indicating higher cell death at lower CdCl2 concentration in those cells.
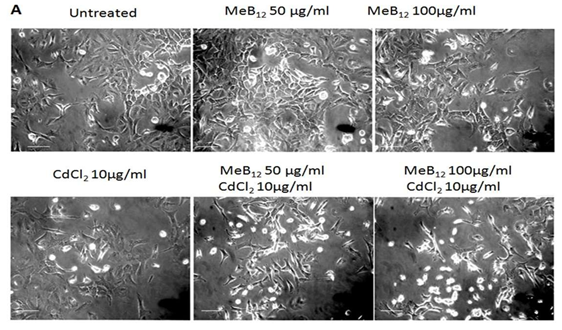
Figure 1A: Effect of Meb12 and Cadmium on A549 Cell Morphology and Viability.
Cells were grown in regular medium (DMEM +10% FBS) as well as media supplemented with two different concentrations of MeB12 (50 & 100 µg/ml) for 48 hours. All these cells were exposed to increasing concentrations of CdCl2 for 24 hours. After 24 hours cells were observed under microscope (Scale 20µm) and counted using hemocytometer after typan blue treatment. This is the representative picture (one of three different experiments). The data of three individual experiments are summarized in this Figure1B. (*) p< 0.05 in comparison with the respective control in each set, (#) p< 0.05 value in comparison with corresponding CdCl2 concentration in CdCl2 set.
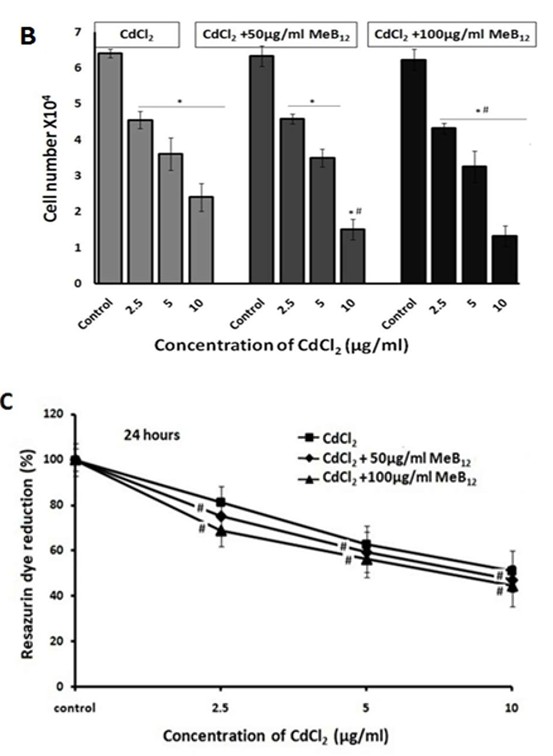
Figure 1B: Effect of Meb12 and Cadmium on A549 Cell Morphology and Viability.
Cells were grown in regular medium (DMEM +10% FBS) as well as media supplemented with two different concentrations of MeB12 (50 & 100 µg/ml) for 48 hours. All these cells were exposed to increasing concentrations of CdCl2 for 24 hours. After 24 hours cells were observed under microscope (Scale 20µm) and counted using hemocytometer after typan blue treatment. This is the representative picture (one of three different experiments). The data of three individual experiments are summarized in this Figure 1B. (*) p< 0.05 in comparison with the respective control in each set, (#) p< 0.05 value in comparison with corresponding CdCl2 concentration in CdCl2 set.
Figure 1C: Cell viability assay: After 24 hours of incubation at 37°C, the cell viability was measured using reassuring reagent and related to that of non-treated controls (set to 100%).
The data of three individual experiments are summarized in this figure. DATA was analyzed by two- way ANOVA followed by multiple comparisons. (#) p< 0.05 value in comparison with corresponding CdCl2 concentration in CdCl2 set.
| Experiment Set | CdCl 2 | CdCl +50µg/ml MeB 2 12 | CdCl +100 µg/ml MeB 2 12 |
|---|---|---|---|
| IC50 values (µg/ml of CdCl ) 2 | 9.2 ±0.841 | 8.5±0.543 | 7.8 ±0.657 |
Table 2: The half maximal inhibitory concentration (IC50) of CdCl2.
Data from three individual experiments are summarized in this figure and expressed as mean ±SD. (*) p< 0.05 in comparison with the respective control in each set, (#) p<0.05 value in comparison with corresponding CdCl2 concentration in CdCl2 set).
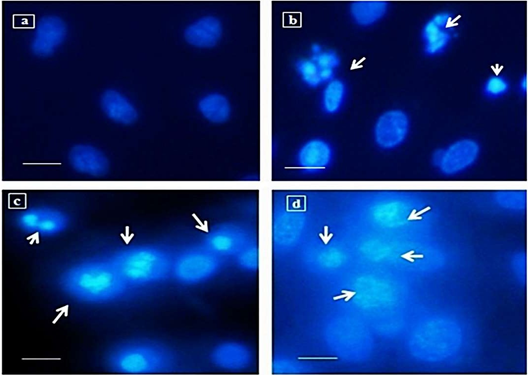
Assessment of Nuclear Damage by Hoechst Staining
Nuclear staining using Hoechst 33342 showed chromatin condensation and DNA damage in CdCl2 exposed A549 cells. The nuclear damage due to cadmium was higher in the A549 sets grown in MeB12 supplemented media compared to that grown in regular medium. In cells exposed to 10 µg/ml of CdCl2 most of the nucleus appeared fragmented with condensed chromatin. Figure 2 compares the extent of nuclear fragmentation and chromatin condensation on treatment with 10µg/ml of CdCl2 in A549 cells grown in regular medium and media supplemented with MeB12 compared to the control untreated cells.
Expression Studies of Apoptotic Genes
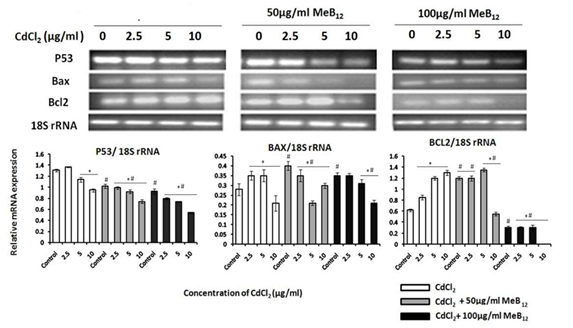
P53 mRNA expression was down regulated with increase in CdCl2 concentration in all the three experimental sets. Even though it appeared that P53 mRNA level increased in the CdCl2 concentration of 2.5µg/ml in A549 cells in regular media, this variation was not significant enough to say that it was up regulated. While in cells grown in MeB12 containing media, the expression level of P53 mRNA decreased with increase in CdCl2 concentration. The degree of down regulation of P53 increased not only with CdCl2 concentration but also with the rise in the concentration of MeB12 in the media (Figure 3). Anti-apoptotic gene BCL2 was over expressed with increasing cadmium concentration. With the combination of MeB12 and increasing cadmium dosage BCL2 expression level started declining. In the cells grown in 100µg/ml of MeB12 there was a high-level down regulation of BCL2 gene expression which further went down with increasing dosage of cadmium. Apoptotic regulator gene BAX was seen to be consistently expressed in response to cadmium stress, only in cells grown in highest concentration of MeB12 (Figure 3).
Expression of Metallothionein Genes
Subtypes of metallothionein gene, MT1A and MT2A showed slight but significant up regulation, with increasing concentration of cadmium. Interestingly in combination with MeB12 the expression of both these genes decreased in response to increasing cadmium dosage (Figure 4).
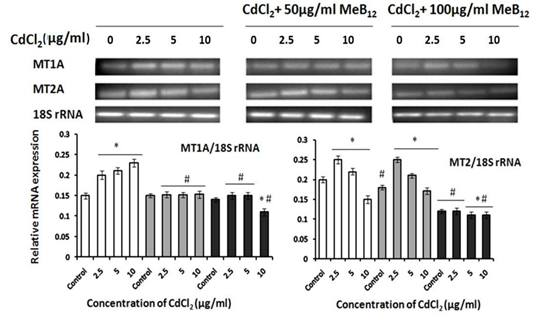
Data from three individual experiments are summarized in this figure and expressed as mean ±SD. (*) p< 0.05 in comparison with the respective control in each set, (#) p< 0.05 value in comparison with corresponding CdCl2 concentration in CdCl2 set).
Promoter Methylation Studies of MT1A and MT2A Genes
Analysis of promoter methylation of metallothionein genes MT1A and MT2A revealed that there is already a significant amount of methylation in MT1A promoter sequence in A549 cells (Figure 5). However, MT2A gene promoter appeared to be unmethylated. There was a slight degree of methylation in MT2A gene promoter in MeB12 treated A549 cells. Interestingly, this effect was further enhanced in the presence of cadmium. An increased level of methylation was observed in the promoter region of MT2A gene in the presence of CdCl2 and MeB12 (Figure 5).
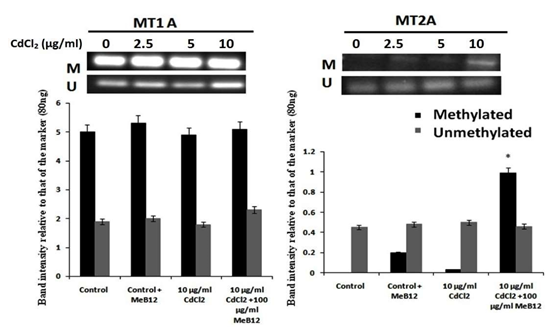
Assessment of Synergy Between Cadmium and Meb12
Isobolographic analysis showed that cadmium and MeB12 interact synergistically towards cell death in A549 cells (Figure 6). The IC50 value of CdCl2 alone was 9.2µg/ml, while that of MeB12 was 287µg/ml. But the same effect (IC50) was achieved by the combinations of these two chemicals in various dose pairs below their individual IC50 values.
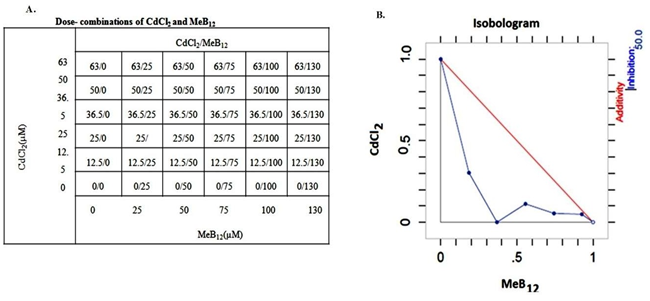
A. Dose combination table showing Percentage increase in cell death, with MeB12 dose on horizontal axis and CdCl2 dose on vertical axis. B. Isobologram showing dose pairs resulting in half the maximum cell death, IC50. Figure 6: Dose combination of CdCl2 and MeB12 and Isobologram: Synergistic interaction between CdCl2 and MeB12, as measured by the degree of cell death in A549 cells.
Discussion
MeB12 is reported to play protective role in different kinds of Revtovich [19, 29, 30]. Current study was undertaken to see the role of methylcobalamin in cadmium induced metal induced stress in A549cell line. Our studies showed that not only did this vitaminB12 analogue has a lethal in combination with cadmium, on A549 cell line but also changed the whole expression pattern of mRNAs of the genes that are involved in the cell signaling cascade during cadmium toxicity. There was a linear increase in cell death with increasing concentration of CdCl2 (Figures 1 & 2, Table 2). This effect was further enhanced by the presence of MeB12 in the media. The extent of nuclear damage by a particular concentration of CdCl2 (10µg/ml) was higher in cells grown in MeB12 containing media compared to those grown in regular medium (Figure 2). All these results show that a combination of these two was more lethal to the cells than either one alone. Similar effects were reported to be exhibited by Vitamin D in metal toxicity [31].
Level of P53 is known to rise drastically in a cell sustaining DNA damage [32]. The response towards increasing degree of toxic agents like UV or any chemicals involves its mRNA up regulation, but here in this study P53 mRNA was found to be down regulated with increase in CdCl2 concentration (Figure 3). The reason for this may be because of degradation of P53 mRNA in the toxic metal environment and this may not reflect in P53 protein expression at those toxic levels, which in many studies is found to be stabilized and continued to over express along with the increase in toxicity unlike their mRNA [32, 33, 34]. BCL2 is the main member of family of apoptosis regulator proteins. It is found to be anti- apoptotic and has been shown to inhibit caspase8 [35]. BAX gene product is a pro-apoptotic protein. The transcription of BAX gene is controlled by the transcription factor P53 and is involved in P53 mediated apoptosis [36].
Apoptotic gene expression profile shows that surviving cells in the monolayer of those grown in regular medium with CdCl2 express high level of anti-apoptotic gene BCL2 to resist programmed cell death. But this survival mechanism was lowered in when CdCl2 was in combination with MeB12 which is evident in cells exposed to 10 µg/ml CdCl2 in combination with 50 µg/ml MeB12 (Figure 3). High level of BCL2 is one of the factors determining cell survival [37]. Up regulation of BCL2 in viable attached cells shows their survival mechanism in the hostile condition which was exhibited by A549 cells in regular media when exposed to increasing cadmium concentration. Cells in 100µg/ml of MeB12 showed not only high degree of BCL2 down regulation but also comparatively higher level of BAX expression when exposed to CdCl2.
The two metallothionein isoforms; MT1A and MT2A showed an mRNA up regulation in response to cadmium in A549 cells in regular medium. While in cells grown in MeB12 the expression of these two isoforms were seen to be decreasing with the increasing concentration of cadmium (Figure 4). Further studies on promoter methylation revealed that MT2A gene promoter is indeed getting methylated in the presence of MeB12, and the level of methylation appeared to be enhanced in the presence of cadmium, providing more evidence on the reason behind the synergy between these two compounds (Figure 5).
Based on the experimental evidence from the current study and from various literature studies the possible mechanism behind synergistic effect of these two compounds on the cell stress is summarized below (Figure 7).
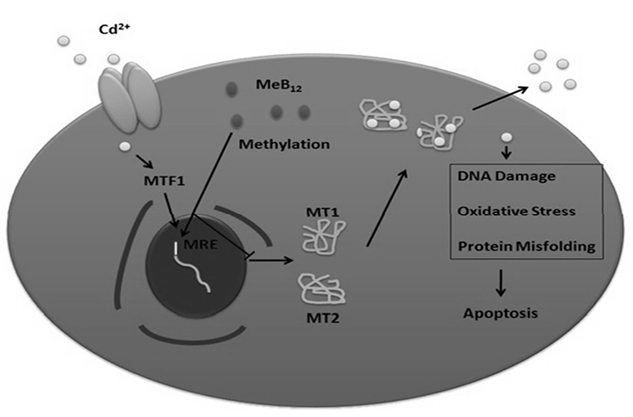
Cadmium enters the cell through protein channels [34], causes mis-folding of proteins [2], DNA damage, oxidative stress etc [26]; leading to the activation of P53 mediated apoptosis involving initiator caspase9 and effecter caspase. Cellular elimination of cadmium involves the activation of the transcription factor MTF1 on cadmium entry and its subsequent translocation to the nucleus [20]. In the nucleus, MTF1 will bind and activate the expression of genes having metal response elements (MRE) in their promoter region, like MT1 and MT2. MT1 and MT2 proteins bind to cadmium there by sequestering it and preventing it from binding to cellular proteins and organelles, alleviating the cellular damages caused by cadmium. One of the mechanisms of down regulation of metallothionein genes transcriptionally is the methylation of the MREs in their promoter region [9]. So being a methylating agent the possible mechanism of down regulation of metallothionein genes by MeB12, thereby sensitizing them to cadmium, may be through methylation of promoter region of these genes.
All these observations show that exposure to a combination of MeB12 and CdCl2 was more lethal to cells than exposure to either of them alone. To summarize MeB12 was found to have no protective role against cadmium induced cytotoxicity in A549 cells. More over along with cadmium it acted as a synergistic effecter of cell death in A549 cells. It also changed the apoptotic signaling mechanism involved during this particular metal toxicity. MeB12 seems to suppress stress response pathways making the cells more susceptible to metal induced cytotoxicity.
Studies on the effect of MeB12 on cadmium induced stress in A549 revealed an interesting property of this vitamin B12 analog- sensitizing the cells to cadmium. Verifying that this property is cancer cell specific and if this sensitization is possible for any cancer drug, might open a new idea for tumor specific cancer drug delivery using MeB12.
Funding
GR thanks Council of Scientific and Industrial Research (CSIR), Govt. of India for the fellowship grant (File No: 09/919(0013)/2011-EMR-I); and AS acknowledges Department of Biotechnology (DBT), Ministry of Science & Technology, Govt. of India for the research funding (grant No. 6242-P59/RGCB/PMD/DBT/ANSR/2015).
Conflict of Interests
Authors declare there is no conflict of interest in this work.
References
-
Drennan CL, Matthews RG, Ludwig ML (1994) Cobalamin- dependent methionine synthase: the structure of a methylcobalamin-binding fragment and implications for other B12-dependent enzymes. Current Opinion in Structural Biology 4(6): 919-929.
-
Yaqub BA, Siddique A, Sulimani R (1992) Effects of methylcobalamin on diabetic neuropathy. Clinical Neurology and Neurosurgery 94(2): 105-111.
-
Jian-Bo L, Cheng-Ya W, Jia-Wei C, Xiao-Lu L, Zhen-Qing F, et al. (2010) The preventive efficacy of methylcobalamin on rat peripheral neuropathy influenced by diabetes via neural IGF-1 levels. Nutritional Neuroscience 13(2): 79- 86.
-
Weinberg J, Sauls D, Misukonis M, Shugars D (1995) Inhibition of productive human immunodeficiency virus-1 infection by cobalamins. Blood 86(4): 1281- 1287.
-
Wimmer U, Wang Y, Georgiev O, Schaffner W (2005) Two major branches of anti- cadmium defense in the mouse: MTF-1/metallothioneins and glutathione. Nucleic Acids Research 33(18): 5715-5727.
-
Rosenfeld J, Ellis A (2008) Nutrition and Dietary Supplements in Motor Neuron Disease. Physical Medicine and Rehabilitation Clinics of North America 19(3): 573-589.
-
Izumi K, Fujise T, Inoue K, Mori H, Yamazaki K, et al. (2013) Mecobalamin improved pernicious anemia in an elderly individual with Hashimoto’s disease and diabetes mellitus. Japanese Journal of Geriatrics 50(4): 542-545.
-
World Health Organization (2006) Inorganic and organic lead compounds. IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. World Health Organization & International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans/ World Health Organization, International Agency for Research on Cancer 87: 1-471.
-
Sarkar A, Pilani B, Goa B, Ravindran G, Krishnamurthy V (2013) A brief review on the effect of cadmium toxicity: from cellular to organ level Studies on Anticoagulant Potential of Moon Jellyfish (Aurelia aurita) and Barrel Jellyfish (_Rhizostoma pulmo_) Tentacle Extracts 3.
-
Tran TA, Popova LP (2013) Functions and toxicity of cadmium in plants: Recent advances and future prospects. Turkish Journal of Botany 37: 1-13.
-
Friberg L, Elinder CG, Kjellström T (1992) WHO Task Group on Environmental Health Criteria for Cadmium. United Nations Environment Programme. International Labour Organisation., International Program on Chemical Safety. Cadmium. World Health Organization.
-
Goyer RA (1997) Toxic and Essential Metal Interactions. Annual Review of Nutrition 17(1): 37-50.
-
Anetor JI, Ajose F, Anetor GO, Iyanda AA, Babalola OO, et al. (2008) High cadmium / zinc ratio in cigarette smokers: potential implications as a biomarker of risk of prostate cancer. Nigerian Journal of Physiological Sciences 23(1-2): 41-49.
-
Ali S, Hussain S, Khan R, Mumtaz S, Ashraf N, et al. (2019) Renal toxicity of heavy metals (cadmium and mercury) and their amelioration with ascorbic acid in rabbits. Environmental Science and Pollution Research 26(4): 3909-3920.
-
Fleeson W, Jayawickreme E, Jones ABAP, Brown NA, Serfass DG, et al. (2017) Journal of Personality and Social Psychology 1(1): 1188-1197.
-
Zhai Q, Narbad A, Chen W (2015) Dietary strategies for the treatment of cadmium and lead toxicity. Nutrients 7(1): 552-571.
-
Kikuchi M, Kashii S, Honda Y, Tamura Y, Kaneda K, et al. (1997) Protective effects of methylcobalamin, a vitamin B12 analog, against glutamate-induced neurotoxicity in retinal cell culture. Investigative Ophthalmology & Visual Science 38(5): 848-854.
-
Mori K, Kaido M, Fujishiro K, Inoue N, Ide Y, et al. (1991) Preventive effects of methylcobalamin on the testicular damage induced by ethylene oxide. Archives of Toxicology 65: 396-401.
-
Revtovich AV, Lee R, Kirienko NV (2019) Interplay between mitochondria and diet mediates pathogen and stress resistance in _caenorhabditis elegans_. PLoS Genetics 15(3).
-
Patrick L (2003) Toxic metals and antioxidants: Part II. The role of antioxidants in arsenic and cadmium toxicity. Alternative Medicine Review : A Journal of Clinical Therapeutic 8(2): 106-128.
-
Zakharyan RA, Aposhian HV (1999) Arsenite methylation by methylvitamin B12 and glutathione does not require an enzyme. Toxicology and Applied Pharmacology 154(3): 287-291.
-
Yagle MK, Palmiter RD (1985) Coordinate regulation of mouse metallothionein I and II genes by heavy metals and glucocorticoids. Molecular and Cellular Biology 5(2): 291-294.
-
Masters BA, Kelly EJ, Quaife CJ, Brinster RL, Palmiter RD (1994) Targeted disruption of metallothionein I and II genes increases sensitivity to cadmium. Proceedings of the National Academy of Sciences of the United States of America 91(2): 584-588.
-
Foster KA, Oster CG, Mayer MM, Avery ML, Audus KL (1998) Characterization of the A549 cell line as a type II pulmonary epithelial cell model for drug metabolism. Experimental Cell Research 243(2): 359-366.
-
Stellavato A, Cammarota M, Miraglia N, Simonelli A, Giuliano M (2011) An alternative gas-phase _in vitro_ exposure system for toxicity testing: The interaction between nitrous oxide and A549 cells. ATLA Alternatives to Laboratory Animals 39(5): 449-459.
-
Tallarida RJ (2012) Quantitative Methods for Assessing Drug Synergism. Genes and Cancer2(11): 1003-1008.
-
Zhao J, Kelnar K, Bader AG (2014) In-depth analysis shows synergy between erlotinib and miR-34a. PLoS ONE 9(2).
-
Soo-il L (2010) Drug interaction: Focusing on response surface models. Korean Journal of Anesthesiology 58(5): 421-434.
-
Mori K, Kaido M, Fujishiro K, Inoue N, Ide Y, et al. (1991) Preventive effects of methylcobalamin on the testicular damage induced by ethylene oxide. Archives of Toxicology 65: 396-401.
-
Nishimoto S, Tanaka H, Okamoto M, Okada K, Murase T, et al. (2015) Methylcobalamin promotes the differentiation of Schwann cells and remyelination in lysophosphatidylcholine-induced demyelination of the rat sciatic nerve. Frontiers in Cellular Neuroscience pp: 9.
-
Moon J (1994) The role of vitamin d in toxic metal absorption: A review. Journal of the American College of Nutrition 13(6): 559-564.
-
Kastan MB, Onyekwere O, Sidransky D, Vogelstein B, Craig RW (1991) Participation of p53 Protein in the Cellular Response to DNA Damage. Cancer Research 51(23 Pt 1) 6304-6311.
-
Hockley SL, Arlt VM, Brewer D, Giddings I, Phillips DH (2006) Time- and concentration-dependent changes in gene expression induced by benzo(a)pyrene in two human cell lines, MCF-7 and HepG2. BMC Genomics.
-
Lakin ND, Jackson SP (1999) Regulation of p53 in response to DNA damage. Oncogene 18(53): 7644-7655.
-
Chang YC, Xu YH (2000) Expression of Bcl-2 inhibited Fas- mediated apoptosis in human hepatocellular carcinoma BEL-7404 cells. Cell Research 10(3): 233-242.
-
Chipuk JE, Kuwana T, Bouchier-Hayes L, Droin NM, Newmeyer DD, et al. (2004) Direct Activation of Bax by p53 Mediates Mitochondrial Membrane Permeabilization and Apoptosis. Science 303(5660): 1010-014.
-
Chang TC, Wentzel EA, Kent OA, Ramachandran K, Mullendore M, et al. (2007) Transactivation of miR- 34a by p53 Broadly Influences Gene Expression and Promotes Apoptosis. Molecular Cell 26(5): 745-752.
- Evaluation of Skin Aging Preventive Effects of Cherry Blossom Petal Extracts Through Antioxidant and Anti-Glycation Activities
- Is Cell Death Responsible for False Positive Results of In Vivo Comet Assay?
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism