Inhibition of Snake Venom Serine Proteases Activities by Specific Antibody
According to the World Health Organization, snake envenomation is a major neglected public health problem. In addition to deaths, these accidents cause other severe disabilities, such as amputations. Annually, around 2 million people worldwide are affected by snakebite, with Africa, Asia, and Latin America being the regions most affected. Snakes of the Bothrops genus are the main cause of snakebite accidents in Latin America. Serine proteases (SVSPs) are one of the main protein families that have been implicated in the alterations of the human hemostasis, which causes a tendency to increase thrombotic and hemorrhagic processes. Part of SVSPs, are thrombin-like enzymes (TLEs), which recognize and cleave human fibrinogen, but usually only releasing fibrinopeptide A or B and do not act on the coagulation Factor XIII. Consequently, these toxins contribute to coagulopathy by fibrinogen consumption, one of the major systemic hemostatic disturbances, frequently observed in snakebite victims. The bothropic antivenom is effective in reversing most of the systemic effects of envenomation when administered early in an adequate therapeutic dose. However, studies have demonstrated that, in some cases, antivenoms do not completely neutralize the action of SVSPs, which are co-responsible for systemic and local effects, such as coagulopathy and hemorrhage.In this context, a better understanding of SVSPs’ role in coagulopathy caused by envenomation, and developing new strategies to inhibit these toxins are important. To achieve this, we aimed in the current work to isolate an enriched pool of SVSPs from Bothrops jararaca and Bothrops atrox, to better understand the SVSPs’ interaction with murine monoclonal antibody (mAb) anti-SVSPs.
Introduction
Snake envenomation is a neglected public health problem, occurring more frequently in populations with limited access to health services [1]. Approximately 2 million people worldwide are affected by snakebite every year, with Africa, Asia, and Latin America being the regions most affected by it [2].
In Latin America, snakes of the Bothrops genus are the leading cause of snakebite accidents. Brazil is the country with the highest number of cases on the continent, registering approximately 19.882 snakebites and 66 deaths per year, with Bothrops snakes causing 90% of these accidents [3]. Peru has 4,500 documented cases per year, Venezuela has 2,500 to 3,000 cases, Colombia has 2,675 cases, Ecuador has 1,200 to 1,400 cases, and Argentina has 1,150 to 1,250 cases [4]. In Colombia and Costa Rica, Bothrops asper is the species responsible for the majority of accidents [5, 6].
Bothrops’ venom has a wide range of effects on the victim’s body. It exhibits proteolytic effects Nishida S, et al. [7] pro-coagulant properties Luciano PM, et al. [8] and induces hemorrhage Yamashita KM, et al. [8, 9] and pro- inflammatory responses [10]. These effects lead to various local manifestations, including pain, edema, bruising, blisters, necrosis, and gangrene Azevedo-Marques MM, et al. [11, 12] as well as systemic effects like gingival bleeding, epistaxis, hematuria, acute renal failure, hematemesis, hypotension, and shock [13, 14].
The overall composition of bothropic venom includes several components, such as metalloproteases (SVMPs), serine proteases (SVSPs), phospholipases A2 (PLA2), C-type lectins, bradykinin enhancers, L-amino acid oxidases, hyaluronidases, and venom endothelial growth factors [15, 16].
Phospholipases (PLA2s) are one of the most thoroughly studied enzyme proteins present in venoms [17]. These enzymes have important functions in several biological processes such as local and/or systemic myotoxicity Andria˜o- Escarso SH, et al. [18, 19] and inflammatory activity [20].
A rapid inflammatory process at the site of venom injection causes the increase of permeability of capillaries and venules by direct action of the components of the venom on microvasculature and the effect of endogenous mediators, lead to the formation of edema [21, 22]. Animals inoculated with Bothrops atrox venom showed an increase in Aspartate aminotransferase (AST) and alanine aminotransferase (ALT) levels, indicating tissue and liver abnormalities Talwer GP, et al. [23] as demonstrated by Gonçalves, et al. for B. jararaca in rats and Chaves, et al. [24, 25] for B. asper in mice.
The venom toxins also interact with the hemostatic system, causing disruptions in the endothelium, activation of the coagulation cascade and consequently enhancing the fibrinolytic activity, leading to increased hemorrhagic and procoagulant processes. SVMPs and SVSPs are the two main protein families responsible for the local and systemic effects. They play crucial roles in the proteolytic degradation of endothelial cell surface proteins, as well as in the induction of inflammatory, proteolytic, hemorrhagic, and procoagulant effects [26, 27].
SVMPs and SVSPs are the two main protein families responsible for the local and systemic effects. Unclotable blood is one of the most characteristic effects induced by Bothrops envenomation and this outcome is closely associated with the action of SVMPs. Snake venom metalloproteases can also cause proteolysis of basal lamina components in microvessels, resulting in the loss of vascular wall integrity and leading to blood extravasation into the skin [21, 28]. However, Bjarnason, et al. [29] and Perez, et al. [30] described the SVSPs as co- responsible for acting in a synergic manner with the SVMPs on the local hemorrhagic effect. In concordance to these authors, Santoro, et al. described the venom of the Bothrops jararaca snake as a rich mixture of enzymes and proteins that destabilize hemostasis in a multi- factorial manner [31].
Among SVSPs, there is a group of toxins able to recognize and cleave human fibrinogen, named thrombin-like enzymes. However, unlike human thrombin, these enzymes generally cleave either the α-chain or the β-chain of fibrinogen to give fibrinopeptide A or B, which results in the consumption of fibrinogen without forming stable fibrin. Moreover, these toxins usually do not activate factor XIII, and consequently an unstable fibrin network more susceptible to the action of the fibrinolytic system occurs. As a consequence, these toxins contribute to coagulopathy by consuming fibrinogen, which is a major systemic hemostatic disturbance commonly observed in snakebite victims [32, 33, 34]. Serine proteases also interfere with different aspects of human hemostasis, such as platelet aggregation and coagulation cascade [35].
The administration of bothropic antivenom in envenomed patients is recognized by the WHO as the recommended treatment for snakebite accidents [36, 37]. The antivenom is effective in reversing most of the systemic effects of envenomation when administered early in an adequate therapeutic dose [36].
Studies have shown that the bothropic antivenom may not fully reverse the local effects of the venom, even when used in large quantities, and the time elapsed between the accident and treatment can lead to temporary or permanent disability of the affected limb [38]. Furthermore, some papers have demonstrated that bothropic antivenom does not completely neutralize the action of SVSPs, present in the venoms [39].
Therefore, it is essential to gain a better understanding of SVSPs’ action, in coagulopathies caused by envenomation and try to develop new strategies to inhibit these toxins during the patient’s treatment. To achieve this, in this current work, we isolated enriched pools of SVSPs from Bothrops snakes and evaluated the interaction of these toxins in the presence of murine monoclonal (mAb) anti-SVSPs in experiments that simulated human coagulation.
Material and Methods
Reagents
Activated partial thromboplastin time reagent (aPTT clot), containing ellagic acid and synthetic phospholipids and Prothrombin time clot reagent (PT clot), containing tissue thromboplastin (rabbit brain extract) were purchased from BIOS Diagnóstica® (São Paulo, SP, Brazil). The column Benzamidine Sepharose 6B, was obtained from GE HealthCare. The chromogenic substrate S-2238 (H-D-Phe-Pip-Arg-pNa, 2HCl) was purchased from Chromogenix (Milano, Italy). The other reagents used are of the highest possible purity.
Venom and Monoclonal Antibody Antivenom
Bothrops jararaca venom (batch: 220007) was supplied by the Hyperimmune Plasmas Processing Section, Butantan Institute, São Paulo, Brazil. The Bothrops atrox venom (batch: ATX 21/003) was supplied by CETA Ltda, Morungaba, São Paulo, Brazil. The monoclonal antibody anti-serine protease (mAb-anti-SVSPs - (6AD2-G5) was previously purified by group.
Blood and Plasma Samples
Pool (162 mL) of citrated (0.32% final concentration) human plasma, was a donation of Colsan (Associação Beneficiente de Coleta de sangue) (Hemorrede SP/SUS) (Av Jandira, 1260), São Paulo, Brazil. Aliquots were stored at -80°C. The samples were collected immediately before use and maintained cooled. After obtaining the blood, the platelet poor plasma (PPP) was obtained by centrifugation at 25°C for 20 min at 2,500 x g. The samples were stored at - 20°C until use.
Affinity Chromatography
Venoms (Bothrops jaracara and Bothrops atrox) were fractionated by affinity chromatography on a Benzamidine Sepharose (HiTrap TM) column (0.7 x 2.5 cm, 1mL) (Amersham Pharmacia Biotech AB, Uppsala, Sweden), previously equilibrated with buffer A (Tris-HCl 0.05M, pH 7.6). Then, the elution was performed constant flow of 1mL/ min with buffer (Tris-HCl 0.05 M, pH 7.6 + NaCl 0.5 M) as eluent B and (Glycine-HCl 0.02 M, pH 3.2) as buffer C [40]. All peak profiles were monitored by their absorbance at A280 nm and A214 nm. The fractions were equilibrated in buffer A, desalted, and concentrated into an Amicon® System containing a 3kDa size exclusion filter (Amicon, Millipore, Germany).
Analysis of Fibrinogen Cleavage Inhibition by Anti-SVSP mAb
Sixteen micrograms of human fibrinogen (Sigma- Aldrich, MO, USA) was incubated with enriched pools of SVSPs (0.6 μg) and mAb anti-SVSPs 6AD2-G5 (4 μg, 8 μg and 16 μg) for 1h at 37°C in a wet bath under constant gentle agitation. Next, samples were submitted to a 10% SDS-PAGE under reducing conditions and the gels were stained with Coomassie Brilliant Blue R-250. The fibrinogenolytic activity was determined by the cleavage of α, β and/or γ chains of the fibrinogen.
Determination of the Inhibiting Activity of mAb Anti-SVSP In Hydrolysis of H-D- Phe-Pip-Arg- pNan·2HCl by SP-BjV and SP-BaV
The inhibition of SP-BjV and SP-BaV by mAb anti-SVSP was determined by the residual of enzymatic activity on the chromogenic substrate specific to thrombin-like enzymes, H-D- Val-Leu-Arg-pNan (S-2238 - Chromogenix). In typical experiments, 2µg of SP-BjV and 1 µg of SP- BaV were pre- incubated with increase quantities of mAb anti-SVSP (1, 2, 4, 8 and 16 µg) at 37°C and, after 10 minutes, 20 µL of the substrate (2mM) was added, in a final volume of 100µL, continuing incubation for 30 minutes at 37°C. The hydrolysis of the substrate was accompanied by photometric reading A405 nm of p-nitroaniline released in a SpectroMax® ABS Plus microplate reader, using software (SoftMax Pro Software 7.1.2). The experiments were carried out in triplicate.
Thromboelastometric Assays with Human Plasma Samples
Fibrin formation in platelet poor plasma (PPP) samples in presence or absence of mAb anti-SVSP (16 µg) were recorded in a computerized ROTEM® four-channel system (Pentapharm, Munich, Germany), according to the manufacturer’s instructions for intrinsic pathway (INTEM) thromboelastometry assays during 60 min (n = 3, each experimental group). For evaluation of the possible effect of mAb anti-SVSP as a serine protease inhibitor on the INTEM profile, mAb anti-SVSP (16µg) was incubated at the above described volume of 60 μL. The clotting time (CT, in seconds, represented by the start of the reaction to beginning of the clot formation) was analyzed.
Standardization of the Serine Protease Pool Mean Coagulant Dose (MCD)
Experimental groups (EG) (n=5 each) were assayed (in cups with final volume of 340 µL), according to the following protocol: (EG1): addition of 60 μL of saline solution at 0.9% to 260 μL human plasma before calcification with 20 µL of CaCl2 (0. 2M) (negative control); (EG2): 5 µL of the activator aPTT clot reagent (an activator of coagulation) plus 50 μL of saline solution at 0.9% to 260 μL human plasma before calcification with 20 µL of CaCl2 (0. 2M) (positive control) and (EG3) different doses of the serine protease pool (SVSPs) solubilized in the 60 µL of saline solution at 0.9% to 260 μL human plasma before calcification with 20 µL of CaCl2 (0. 2M) (assay test), for determination of its mean coagulant dose (MCD). The MCD of the serine protease pool was considered as that amount that shortens the CT parameter of the negative control group to an interval situated between the minimum and maximum coagulant response.
Statistical Analysis
The significance of the statistical difference between the experimental data obtained in the in vitro experiments was analyzed using the GraphPad Prism program (version 8.0 Prism, GraphPad). P values < 0.05 were considered statistically significant. The results were expressed as the mean ± standard error of the mean (SEM). For values of ROTEM, it was used the ANOVA, Newman-Keus post-test for analyzing the difference in relation to control values.
Results and Discussion
Isolation and Characterization of Bothrops Jararaca and B. atrox Venoms (BjV, BaV) Serine Proteases
The fractionation of B. jararaca and B. atrox venoms was performed by in two consecutive chromatographic process. To each venom, in the first, 10 mg of BjV and 15 mg of BaV was separately applied to Benzamidine Sepharose 6B affinity column, resulting in three fractions, denominated: F1, F2 and SP-BjV or SP-BaV (Figures 1A & 1B).
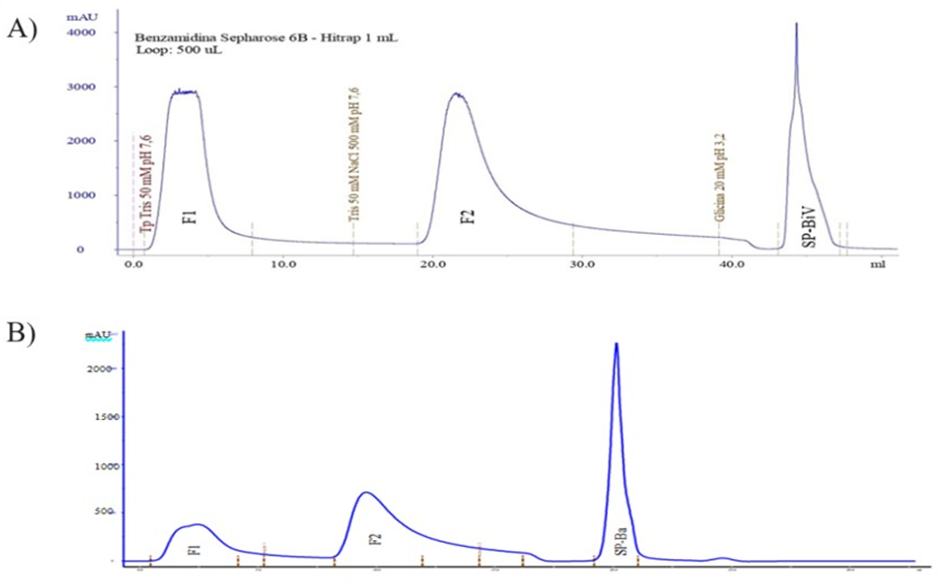
Source: author (2023). Figure 1: Affinity chromatography for BjV and BaV purification.
Figures 1A & 1B Chromatography of Bothrops jararaca and Bothrops atrox venom in Benzamidine Sepharose 6B affinity column. In both purification processes, the column was previously equilibrated with Tris buffer 50 mM pH 7.6, followed by elution with Tris 50 mM NaCl 500 mM pH 7.6 and Glycine 20 mM pH 3.2 under flow of 1.0 mL/min. Each chromatogram shows three fractions; F1; F2 and the fraction containing the SVSPs from each venom, denominated as SP- BjV and SP-BaV.
Analysis of Fibrinogen Cleavage by SVSPs
The coagulopathy by fibrinogen consumption is a major systemic hemostatic disturbance commonly observed in patients envenomed by Bothrops snakebites. During the envenomation there is the action of toxins, as thrombin- like enzymes (TLEs) that also recognize and cleave human fibrinogen [41]. Therefore, in this set of the experiments, we evaluated whether the murine mAb anti-SVSP 6AD2-G5 is able to inhibit the activity of bothropic serine proteases on human fibrinogen.
As observed in Figures 2A & 2B, SP-BaV and SP-BjV (0.6 μg for both) were able to efficiently cleave α and β chains of human fibrinogen. Despite the high proteolytic activity observed in electrophoresis, lower fibrinogen degradation and higher α and β chains preservation are apparently observed, especially for SP-BaV, when pretreated with increasing concentrations of the specific monoclonal antibody. This degradation pattern changes seen in Figure 2A is correlated and is in concordance with the kinetic results obtained using the thrombin-like enzymes’ chromogenic substrate (Figure 3) and by thromboelastometry analysis (Table 1), also showing a tendency for enzymatic inhibition, in the presence of the monoclonal antibody.
![Figure 2: ** Analysis of fibrinogen cleavage by SVSPs. Figures 2A & 2B 10% SDS-PAGE gel stained with Coomassie. (A) pool SP-BaV and (B) pool SP-BjV. 1: Low molecular weight standard; 2: Fibrinogen Control [16µg]; 3: SP-BaV (A) or SP-BjV (B) [0,6µg]; 4: mAb [16µg]; 5: Fibrinogen [16µg]+ P-BaV (A) or SP-BjV (B) [0,6µg]; 6: Fibrinogen [16µg] + SP-BaV (A) or SP-BjV (B) [0,6µg] + mAb [4µg]; 7: Fibrinogen [16µg]+ SP-BaV (A) or SP-BjV (B) [0,6µg] + mAb [8µg]; 8: Fibrinogen [16µg]+ SP-BaV (A) or SP-BjV (B) [0,6µg] + mAb [16µg]](/fulltextimages/11851/fig_2.png)
Source: author (2023). Figure 2: Analysis of fibrinogen cleavage by SVSPs. Figures 2A & 2B 10% SDS-PAGE gel stained with Coomassie. (A) pool SP-BaV and (B) pool SP-BjV. 1: Low molecular weight standard; 2: Fibrinogen Control [16µg]; 3: SP-BaV (A) or SP-BjV (B) [0,6µg]; 4: mAb [16µg]; 5: Fibrinogen [16µg]+ P-BaV (A) or SP-BjV (B) [0,6µg]; 6: Fibrinogen [16µg] + SP-BaV (A) or SP-BjV (B) [0,6µg] + mAb [4µg]; 7: Fibrinogen [16µg]+ SP-BaV (A) or SP-BjV (B) [0,6µg] + mAb [8µg]; 8: Fibrinogen [16µg]+ SP-BaV (A) or SP-BjV (B) [0,6µg] + mAb [16µg]
SVSPs Catalytic Activity and mAb Anti SVSPs Inhibitory Action
To better evaluate the inhibitory activity of this mAb on the catalytic activity of SP-BjV and SP-BaV fractions, the residual activity of these fractions was evaluated on the chromogenic substrate, H-D-Phe-Pip-Arg-pNA-2HCl (S- 2238), specific to thrombin-like enzymes, in the presence of different mAb concentrations. The results show no inhibition of the SP-BjV catalytic action by mAb anti-SVSP 5AD2-G5 (Figure 3A).
On the other hand, a significant inhibition in the SP- BaV activity by mAb anti-SVSP (52.3 %) was observed. However, even at higher mAb anti-SVSPs concentrations, there is no total inhibition of SP-BaV catalytic action (Figure 3B). Kuniyoshi et al., using selective substrates for SVSPs of Bothrops jararaca demonstrated that bothropic antivenom does not fully inhibit the catalytic action of these toxins [39].
Gutiérrez et al. demonstrated by antivenomics studies that B. asper serine proteases are also partially neutralized and have variable immunoreactivity [42]. In concordance, Patra et al., have shown that the enzymatic activities of mid and low molecular weight proteins such as, PLA2 and SVSP, were least neutralized by specific polyclonal antivenom. Interestingly, the SVSPs were well recognized by antivenom, suggesting that the catalytic sites of the enzymes may be poor immunogens [43]. These results reinforce the importance of molecules that inhibit the catalytic action of these enzymes.
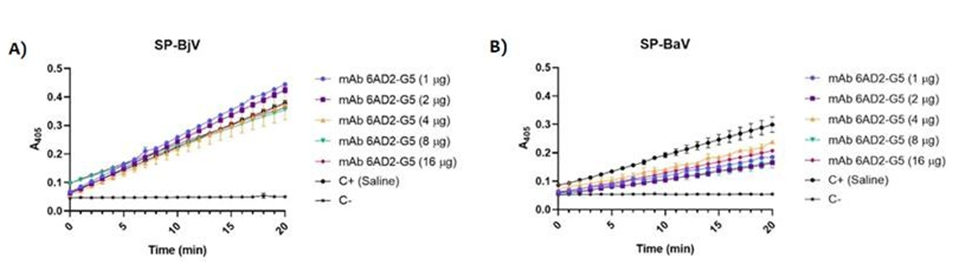
Figure 3 the inhibition percentage value was determined using a specific substrate for thrombin like enzymes, as described in Methods. The experiments were performed in triplicate and the standard deviations of inhibition percentage value did not exceed 10%. Thromboelastometric assays with human plasma samples. It is known that bothropic venoms cause hemostatic disturbances Santoro ML, et al. [31] and our group in the previous work demonstrated in vivo by decrease of the diameter of hemorrhagic halo that the addition of selective peptide inhibitors of SVSPs to the bothropic antivenom improves the efficacy of the treatment of local hemorrhage and the coagulopathy caused by Bothrops jararaca envenomation [44]. So, in this set of experiments, using human plasma samples, we aimed to analyze whether the mAb anti-SVSP 5AD2-G5 is able to restore the clotting time and, consequently, decrease the fibrinogen consumption.
So, in this set of experiments, using human plasma samples, we aimed to analyze whether the mAb anti-SVSP 5AD2-G5 is able to restore the clotting time and, consequently, decrease the fibrinogen consumption.
| Human Pooled Plasma | CT (s) | % inhibition in relation to MCD Control |
|---|---|---|
| Positive Control | 362±19 | --- |
| Negative Control | 1065±63 | --- |
| MCD - SP-BjV (2ug) | 516±15 | |
| MCD SP-BjV (2μg) + mAb (16μg) | 523±26 | No |
| MCD - SP-BaV (1μg) | 320±16 | |
| MCD - SP-BaV (2μg) + mAb (16μg) | 495±14 | 54,68% |
Table 1: Thromboelastometric parameters of INTEM profiles of human pooled plasma.
Abbreviations: INTEM (Intrinsic Pathway Thromboelastometry); CT: Clotting Time; MCD: Mean Coagulant dose Data are Expressed as mean ± SEM (n = 3). Table 1: Thromboelastometric parameters of INTEM profiles of human pooled plasma.
As observed in the Table 01, the mAb anti-SVSP 5AD2-G5, in concordance to previous results obtained by Petretski et al., the Bothrops jararaca fibrinogenolytic action was not inhibited by this mAb. However, in agreement with these authors and ours results from the kinetic assays substrate, a significant inhibition (around 50%) by mAb anti-SVSP was observed on SP-BaV activity on clotting time [45].
These results are very promising, especially in relation to the treatment of envenomation caused by B. atrox bites, that despite the efficacy of bothropic antivenom, quantitative differences in the effective doses are required, when compared to those used to neutralize venoms of the immunizing mixtures [46, 47, 48].
In conclusion, this specific monoclonal antibody (5AD2-G5) inhibits the catalytic action of the serine proteases from Bothrops atrox venom on human fibrinogen, decreasing the fibrinogen consumption during the envenomation, commonly observed in bothropic accidents. The development of fully recombinant anti-toxins monoclonal antibodies could help with the prognosis of these accidents, mostly by reducing the severe coagulopathy.
Conflicts of Interest
The authors declare that there is no conflict of interest.
Funding
We are grateful to the Instituto Butantan, CAPES, FAPESP and CNPq for supporting us in writing this article.
References
-
WHO (2023) Snakebite envenoming: an interactive data platform to support the 2030 targets.
-
Gutiérrez JM, Calvete JJ, Habib AG, Harrison RA, Williams DJ, et al. (2017) Snakebite envenoming. Nature reviews Dis Primer 3(1).
-
TabNet Win32 3.2 (2023) Accident by Venomous Animals- Notifications registered in the Notifiable Diseases Information System, Brazil.
-
Chavez García MV, Medina MS, Martillo STL, Cordova Cedeño EM (2020) Snakebite management. RECIMUNDO 4(1): 46-54.
-
Lizarazo J, Patiño R, Lizarazo D, Osorio G (2020) Hemorragia cerebral fatal después de una mordedura de serpiente Bothrops asper en la región del Catatumbo, Colombia. Biomédica 40(4): 609-615.
-
Coto Freund F, Barquero FM, Rocha Monge SM (2022) Ophidian accident: a focus on management at first level of care. Rev Medica Sinerg 7(2): e756.
-
Nishida S, Fujimura Y, Miura S, Ozaki Y, Usami Y, et al. (1994) Purification and characterization of bothrombin, a fibrinogen-clotting serine protease from the venom of Bothrops jararaca. Biochemistry 33(7): 1843-1849.
-
Luciano PM, Silva GEB, Azevedo-Marques MM (2009) Fatal Botrapch Accident Med. Ribeirão Preto 42(1): 61- 65.
-
Yamashita KM, Alves AF, Barbaro KC, Santoro ML (2014) Bothrops jararaca Venom Metalloproteinases are Essential for Coagulopathy and Increase Plasma Tissue Factor Levels during Envenomation. PLoS Negl Trop Dis 8(5): e2814.
-
Cybulsky MI, Chan MK, Movat HZ (1989) Acute Inflammation and Microthrombosis Induced by Endotoxin, Interleukin-1, and Tumor Necrosis Factor and their Implication in Gram-Negative Infection. Lab Invest 58(4): 365-378.
-
De Oliveira SS, Freitas-de-Sousa LA, Alves EC, de Lima Ferreira LC, da Silva IM, et al. (2017) Fatal stroke after Bothrops snakebite in the Amazonas state, Brazil: A case report. Toxicon 138: 102-106.
-
Azevedo-Marques MM, Cupo P, Hering SE (2003) Accidents caused by venomous animals: venomous snakes. Med Ribeirão Preto 36(2/4): 480-490.
-
Valente HM, Sakai F, Portes-Junior JA, Viana LG, Carneiro SM, et al. (2018) The Primary Duct of Bothrops jararaca Glandular Apparatus Secretes Toxins. Toxins 10(3): 121.
-
De Oliveira SS, de Souza Sampaio V, Alves EC, da Silva VC, de Lima JAA, et al. (2018) Snakebites in the Brazilian Amazon: Current Knowledge and Perspectives. Clinical Toxinology in Australia, Europe, and Americas pp: 73-99.
-
Galizio NDC, Serino-Silva C, Stuginski DR, Abreu PAE, Sant’Anna SS, et al. (2018) Compositional and functional investigation of individual and pooled venoms from long-term captive and recently wild-caught Bothrops jararaca snakes. J Proteomics 30(186): 56-70.
-
Nicolau CA, Prorock A, Bao Y, Neves-Ferreira AGDC, Valente HR, et al. (2018) Revisiting the Therapeutic Potential of Bothrops jararaca Venom: Screening for Novel Activities Using Connectivity Mapping. Toxins 10(2): 69.
-
Mackessy SP (2002) Biochemistry and pharmacology of colubrid snake venoms Journal of Toxicology: Toxin Rev 21(1-2): 43-83.
-
Andria˜o-Escarso SH, Soares AM, Rodrigues VM, Angulo Y, Díaz C, et al. (2000) Myotoxic phospholipases A2 in Bothrops snake venoms: effect of chemical modifications on the enzymatic and pharmacological properties of bothropstoxins from Bothrops jararacussu. Biochimie 82(8): 755-763.
-
Guti_errez JM, Alberto Ponce-Soto L, Marangoni S, Lomonte B (2008) Systemic and local myotoxicity induced by snake venom group II phospholipases A2: comparison between crotoxin, crotoxin B and a Lys49 PLA2 homologue. Toxicon 51(1): 80-92.
-
Teixeira C, Cury Y, Moreira V, Picolo G, Chaves F (2009) Inflammation induced by Bothrops asper venom. Toxicon 54(1): 67-76.
-
Gutiérrez JM, Lomonte B (1989) Local tissue damage induced by Bothrops snake venoms - A review. Mem Inst Butantan. 51(4): 211-223.
-
Farsky SH, Gonçalves LR, Cury Y (1999) Characterization of local tissue damage evoked by Bothrops jararaca venom in the rat connective tissue microcirculation: an intravital microscopic study. Toxicon 37(7): 1079-1083.
-
Talwer GP, Scrivastava LM (1989) Textbook of Biochemistry and Human Biology. 2nd (Edn.), Prentice Hall of Indian, New Delhi.
-
DA, Silva EV, Graça FA, Lira EC, Zanon NM, et al. (2008) In vivo effects of Bothrops jararaca venom on metabolic profile and on muscle protein metabolism in rats. Am J Trop Med Hyg 79(5): 771-778.
-
Chaves F, Gutiérrez JM, Brenes F (1992) Pathological and biochemical changes induced in mice after intramuscular injection of venom from newborn specimens of the snake Bothrops asper (Terciopelo). Toxicon 30(9): 1099-109.
-
Gutiérrez JM, Rucavado A, Escalante T, DíaZ C (2005) Hemorrhage induced by snake venom metalloproteinases: biochemical and biophysical mechanisms involved in microvessel damage. Toxicon 45(8): 997-1011.
-
Santoro ML, Sano-Martins SI (2004) Platelet dysfunction during Bothrops jararaca snake envenomation in rabbits. Thromb Haemost. 92(2): 369-383.
-
Gonçalves-Machado L, Pla D, Sanz L, Jorge BJR, Leitão- De-Araújo M, et al. (2016) Combined venomics, venom gland transcriptomics, bioactivities, and antivenomics of two Bothrops jararaca populations from geographic isolated regions within the Brazilian Atlantic rainforest. J Proteomics 135: 73-89.
-
Bjarnason JB, Fox JW (1994) Hemorrhagic metalloproteinases from snake venoms. Pharmacol Ther 62(3): 325-372.
-
Pérez VA, Saravia P, Rucavado A, Sant’Ana CD, Soares AM, et al. (2007) Local and systemic pathophysiological alterations induced by a serine proteinase from the venom of the snake Bothrops jararacussu. Toxicon 49(7): 1063-1069.
-
Santoro ML, Sano-Martins IS, Fan HW, Cardoso JLC, Theakston RDG, et al. (2008) Haematological evaluation of patients bitten by the jararaca, Bothrops jararaca, in Brazil. Toxicon 51(8): 1440-1448.
-
Castro HC, Zingali RB, Albuquerque MG, Pujol-Luz M, Rodrigues CR (2004) Snake venom thrombin-like enzymes: from reptilase to now. Cell Mol Life Sci 61(7- 8): 843-856.
-
Pradniwat P, Rojnuckarin P (2014) Snake venom thrombin-like enzymes. Toxin Reviews 33(1-2): 16-22.
-
Bhat SK, Joshi MB, Ullah A, Masood R, Biligiri SG, et al. (2016) Serine proteinases from Bothrops snake venom activates PI3K/Akt mediated angiogenesis. Toxicon 124: 63-72.
-
Kini RM (2005) Serine proteases affecting blood coagulation and fibrinolysis from snake venoms. Pathophysiol Haemost Thromb 34(4-5): 200-204.
-
Ministry of health (2023) Brazil.
-
Gutiérrez JM, Rucavado A, Escalante T, Herrera C, Fernández J, et al. (2018) Unresolved issues in the understanding of the pathogenesis of local tissue damage induced by snake venoms. Toxicon 148: 123-131.
-
Cardoso JL, Fan HW, Franca FO, Jorge MT, Leite RP, et al. (1993) Randomized comparative trial of three antivenoms in the treatment of envenoming by lance- headed vipers (Bothrops jararaca) in São Paulo, Brazil. Q J Med 86(5): 315-325.
-
Kuniyoshi AK, Rocha M, Carvalho DC, Juliano MA, Neto LZ, et al. (2012) Angiotensin-degrading serine peptidase: A new chymotrypsin-like activity in the venom of Bothrops jararaca partially blocked by the commercial antivenom. Toxicon 59(1): 124-131.
-
Menaldo DL, Bernardes CP, Santos-Filho NA, Moura LDA, Fuly AP, et al. (2012) Biochemical characterization and comparative analysis of two distinct serine proteases from Bothrops pirajai snake venom. Biochimie 94(12): 2545-2558.
-
Isbister GK (2010) Antivenom efficacy or effectiveness: The Australian experience. Toxicology 268(3): 148-154.
-
Gutiérrez JM, Chaves F, Bolaños R, Cerdas L, Rojas E, et al. (1981) Neutralization of the local effects of Bothrops asper venom by a polyvalent antivenom. Toxicon 19(4): 493-500.
-
Patra A, Kalita B, Chanda A, Mukherjee AK (2017) Proteomics and antivenomics of Echis carinatus carinatus venom: Correlation with pharmacological properties and pathophysiology of envenomation. Scientific Reports 7.
-
Silva GMD, Berto de Souza DH, Waitman KB, Ebram MC, Fessel MR, et al. (2021) Design, synthesis, and evaluation of Bothrops venom serine protease peptidic inhibitors. J Venom Anim Toxins Trop Dis 27: e20200066.
-
Petretski JH, Kanashiro M, Silva CP, Alves EW, Kipnis TL (2000) Two related thrombin-like enzymes present in Bothrops atrox venom. Braz J Med Biol Res 33(11): 1293-1300.
-
Furtado MFD, Maruyama M, Kamiguti AS, Antonio LC (1991) Comparative study of nine Bothrops snake venoms from adult female snakes and their offspring. Toxicon 29(2): 219-226.
-
Muniz EG, Maria WS, Estevão-Costa MI, Buhrnheim P, Chávez-Olórtegui C (2000) Neutralizing potency of horse antibothropic Brazilian antivenom against Bothrops snake venoms from the Amazonian rain forest. Toxicon 38(12): 1859-1863.
-
Sousa LF, Nicolau CA, Peixoto PS, Bernardoni JL, Oliveira SS, et al. (2013) Comparison of Phylogeny, Venom Composition and Neutralization by Antivenom in Diverse Species of Bothrops Complex. PLoS Neglected Tropical Diseases 7(9): e2442.
- Evaluation of Skin Aging Preventive Effects of Cherry Blossom Petal Extracts Through Antioxidant and Anti-Glycation Activities
- Is Cell Death Responsible for False Positive Results of In Vivo Comet Assay?
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism