Exosomes Released from TRAIL Resistance Glioblastoma Cells Influence the Migration and Growth of Healthy Cells
Exosomes are the smallest vesicles that can cross the blood-brain barrier and transport proteins, nucleic acids, and other cell elements to cells. The role of exosomes in drug resistance and their effects on healthy cells have been researched topics in recent years. This study investigated the effect of exosomes released from TRAIL resistance U87 glioblastoma cells on healthy L929 fibroblast cells. U87 cells were incubated with TRAIL (5ng/mL) for three weeks to develop drug resistance and exosomes were isolated with ultracentrifuge. The obtained exosomes were incubated at a concentration of 1 μg/mL-100 μg/ mL on L929 cells for 48 hours, and their cytotoxic effect was examined with MTT assay. After incubation, CXCR4 Protein and gene expression levels of CXCL-12, PAK4, and 14-3-3β were also examined. This study showed that exosomes released from TRAIL-resistant cells increase CXCR4, CXCL-12, and PAK4 levels in healthy cells and do not cause changes in 14-3-3 β oncogenic protein. While 100 μg/mL exosome affected L929 cells, no change was seen at a lower concentration. Exosomes affect the migration potential of other cells due to drug resistance. It may contribute to developing different resistance mechanisms in the long term by changing the proliferation and migration abilities of healthy cells around the tumor cell.
Introduction
Extracellular vesicles (EVs) are biological nanoparticles that carry the heterogeneous phospholipid-bilayer membrane and contents of the cells from which they are produced, enabling intercellular communication [1, 2]. Due to their natural form, EVs are the most successful nanovesicles, both biocompatible and interacting with cells. Among these nanovesicles, they are the smallest (30-140 nm) exosomes, which can enter cells and even cross the blood-brain barrier [3]. Since exosomes are not immunogenic, they are not affected by phagocytic activity, can remain in circulation for a long time, and are protected from enzymatic degradation [4]. Exosomes are not toxic and provide more advantages over synthetic nanoparticles. Therefore, in recent years, exosomes have shown great potential for drug delivery or intracellular transport [5, 6]. Preclinical studies in exosome- mediated drug transport studies have come to the fore in recent years [7, 8].
Glioblastoma multiforme (GBM) is the most common and devastating brain tumor, unsuitable for surgical treatment and highly resistant to radiation and chemotherapy. GBM develops in middle-aged (45 years and older) people and accounts for 52% of brain tumors [9, 10]. GBM is fast growing and quickly recurs with an aggressive spread. Therefore, the potential for drug resistance in chemotherapy is high [11]. When it spreads aggressively, it spreads to the brain using tumor microtubules (TMs) and affects the connecting pathways to the brain. When TM-related structures are formed, the vascular systems allow the tumor to overgrow, and this spread of GBM may prevent its surgical removal. The main function of blood-brain-barriers (BBB) is to prevent therapeutic interventions and inhibit drug delivery and uptake [12]. If the BBB is damaged, the natural barrier function is lost and interrupted, increasing the potential risks of infection or open channels for pathogenicity [13]. Therefore, a delicate balance is important to ensure the integrity of the physical structures of BBBs without deterioration. In GBM treatment, the unique structure of the BBB selects and blocks many things from the microenvironment. In the tumor setting, BBBs cause treatment limitations and reduce drug bioavailability at sites. Nanoparticles designed to cross BBBs in GBM treatment should have high biocompatibility and low side effects [14]. Therefore, to prevent drug resistance in treatment, therapeutics’ roles in the blood-brain barrier and therapeutic and healthy cells other than cancer cells are at the forefront in GBM treatment [15, 16]. This study investigated the effect of exosomes released from resistant glioblastoma, which is known to cross the blood-brain barrier, on healthy cells.
Material and Method
Cell Culture and Drug resistance
U87 human glioblastoma cell line was cultured with a medium containing High Glucose-Modified Dulbecco’s Eagle Medium (DMEM)/F12 (Sigma, D8437) supplemented with 10% (v/v) fetal bovine serum (FBS, Sigma, H1270), 100 units/ml penicillin - 100 mg/ml streptomycin (Sigma, P0781), at 37°C in a humidified atmosphere containing 5% CO2 in 25 cm2 flasks (Corning, 430639). Then cells were incubated with human TRAIL recombinant protein (5 ng/mL, Sigma) for three weeks. During this time, the culture medium was replaced every three days until the cells reached about 80% confluence.
Exosome isolation
U87 glioblastoma cells that developed resistance were maintained in FBS and glucose-free medium for 48 hours [6]. Then the cell medium was collected and centrifuged at 2,000xg for 15 minutes. Next, the cell pellet was removed, the upper supernatant was taken into a new falcon tube and centrifuged at 12,000xg for 30 minutes, and the supernatant was collected [8]. Finally, the supernatant was centrifuged by ultracentrifuge at 90,000×g for 4h (Beckman Coulter, Optima L100). After centrifugation, the pellet was removed and suspended in PBS, and the protein concentration was measured using the BCA kit (Intron Biotechnology, 21071). Particle size, and distribution of exosomes was determined using Zeta Sizer (Nano ZS-90 Malvern Instruments, England).
MTT assay
L929 mouse fibroblast cells were cultured with a medium containing High Glucose DMEM, supplemented with 10% (v/v) fetal bovine serum, 100 units/ml penicillin - 100 mg/ml streptomycin, at 37°C in a humidified atmosphere containing 5% CO2. L929 cells were seeded into 96 well plates with 100 µL DMEM at 104 cells/well then U87 exosomes were placed on them at a concentration of 1 µg/mL, 10 µg/ mL, 100 µg/mL at 48 hours. After the incubation, cell media was changed with fresh DMEM. A total of 10 μL of the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) (ODC Research and Development Inc) labeling reagent was added to each well and incubated for 4 h in a humidified atmosphere at 37°C incubators with 5% CO2 in air [17]. An equal amount of DMSO was added to each well to solubilize formazan precipitate. Then absorbance was measured by a microplate reader at 570 nm.
Crystal violet staining
After the L929 cells were treated with U87 exosomes, the potential for colony formation in the cells was examined. Five hundred cells per well were seeded into 12 well plates, and crystal violet staining was performed seven days after exosome application. After incubation, cells were washed with PBS and fixed with acetic acid/methanol (1:3 ratio) for 30 minutes. After fixation, it was stained with 5% crystal violet solution for 30 minutes and washed with water. Then the images were examined under the microscope. Western blotting and QPCR assay After the L929 cells were treated with U87 exosomes, CXCR4, CXCL-12, PAK4, and 14-3-3 β protein expression levels were analyzed by western blotting [18]. 4-12% SDS PAGE gels were prepared, 30 µg of protein was loaded into each well, and the gel was run with an electric current. The proteins in the gel were transferred to the PVDF membrane using a semi-dry system. The membrane was blocked with 3% BSA for 1 hour. The primary antibody (1:1000: dilution of β-actin, CXCR4, CXCL-12, PAK4, 14-3-3 β from Santa Cruz Biotechnology and ODC Research and Development Inc) was then incubated overnight, and the secondary antibody (1:2000 dilution) was incubated at two hours. The bands were visualized on the imaging system by adding ECL. β-actin was used reference protein.
In addition, PAK4 and 14-3-3 β gene expression levels were analyzed by qPCR [19]. Total RNA from cells was isolated with the kit according to the manufacturer’s instructions (PureLink, Life Technologies) and transformed into cDNA using the kit. One hundred nanograms of cDNA were programmed at 95°C for 5 minutes, then 40 cycles at 95°C for 15 seconds, 60°C for 1 minute using Sybr Green PCR Master Mix (Applied Biosytems) in ABI StepOne Plus detection system. Primer sequences were used for expression of PAK4 (F: 5’GGGGACCCTCGTTCCTATCTA’3 and R: 5’GAGCTGCGTACAGTGGCAA’3) and 14-3- 3β (F: 5’TGGATAAGAGTGAGCTGGTACA’3 and R:
5’CGTGTCCCTGCTCTGTTACG’3). GAPDH was used reference gene and data was analyzed 2-ΔΔCt method.
Results
Cell viability of L929 cells
Cell morphologies of L929 cells were checked with U87 TRAIL resistance exosomes under an inverted microscope for 48 hours. When exosome was applied to L929 cells, there was not much change in cell viability at 1 µg/mL and 10 µg/ mL concentrations, while it increased cell viability at 100 µg/mL exosomes. This increase was found to be statistically significant (p˂0.001, Figure 1).

Migration effects of U87 exosomes on L929 cells
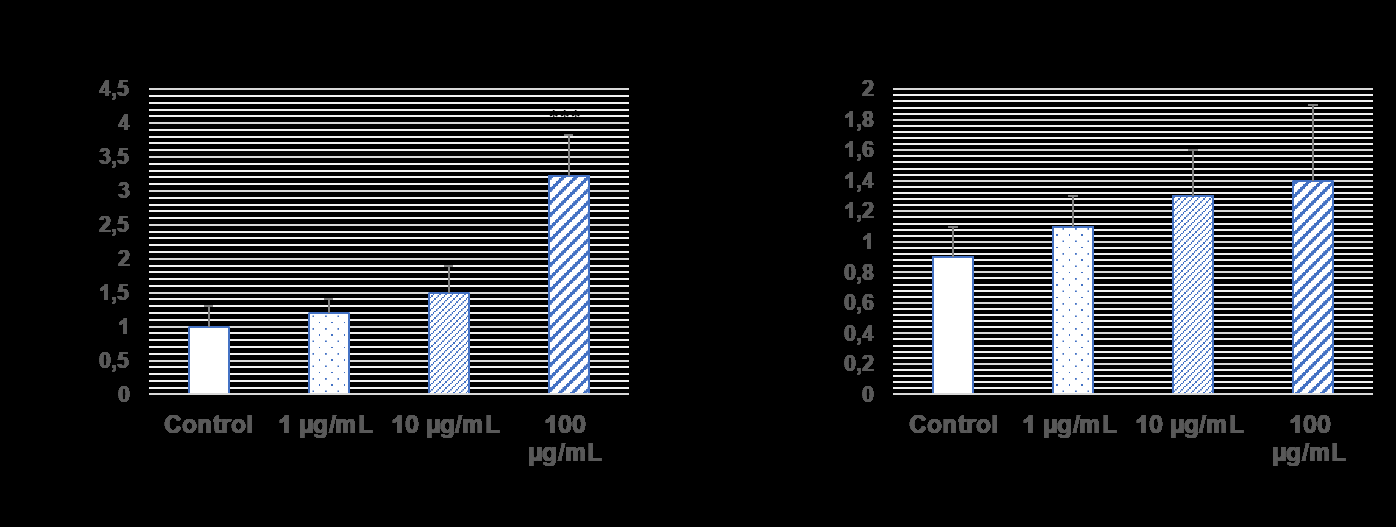
Western blotting and QPCR assay were used to examine the migration effects of TRAIL resistance U87 cells exosomes on L929 cells (Figures 2-3). CXCR4 and CXCL- 12 protein expression levels, which are members of the chemokine family that cause cancer to spread and invade, were significantly increased in 10 µg/mL and 100 µg/mL exosomes in L929 healthy cells (*p˂0.001). However, PAK4 protein expression levels were increased in L929 cells when only 100 µg/mL exosome was administered (*p˂0.001). On the other hand, 14-3-3 β protein expression levels, no significant change was observed in exosome administration.
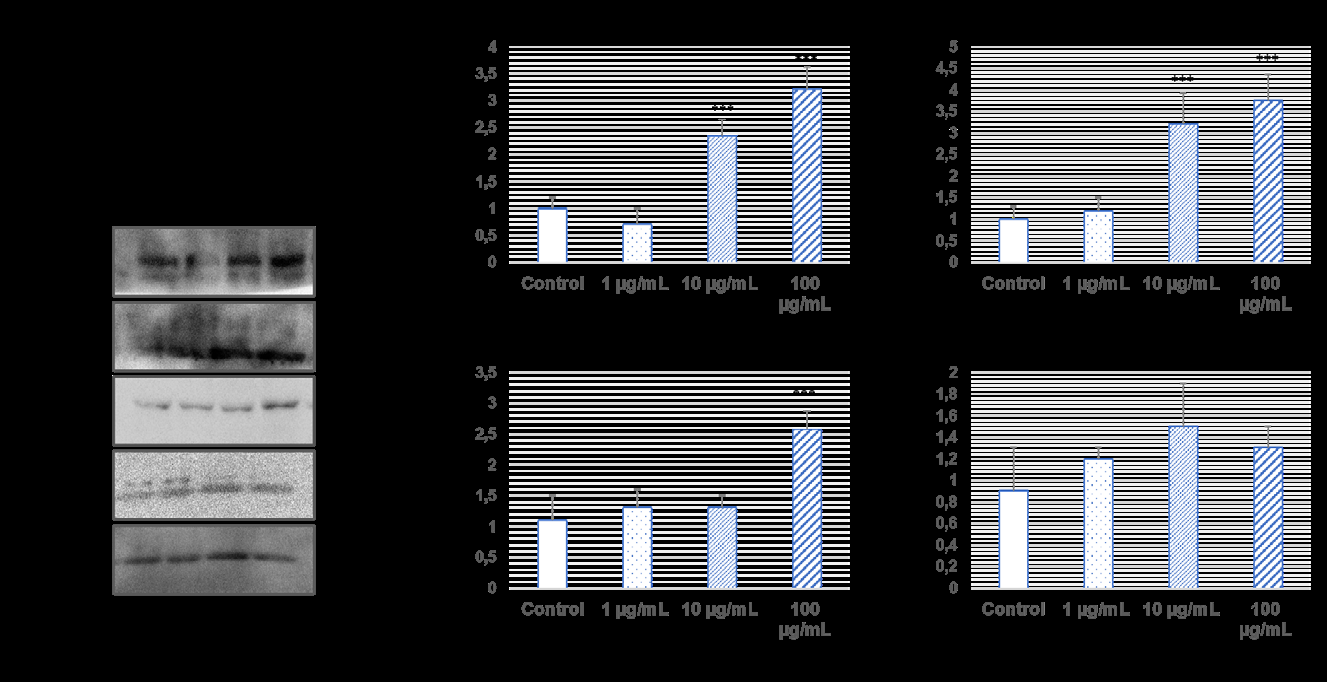
When the exosomes of 1 µg/ml and 10 µg/ml TRAIL resistance U87 cells were added to L929 healthy cells, no significant change was observed, and it increased gene expression at a concentration of 100 µg/ml (***p˂0.001). In addition, no significant changes were observed in 14-3-3 β gene expression levels in the administration of 1 µg/ml, 10 µg/ml, and 100 µg/ml exosomes.
Discussion
TRAIL, known as tumor necrosis factor-associated apoptosis-inducing ligand, is a member of the TNF superfamily. TRAIL shows cytotoxicity in cells and is a protein used in treating various cancers because it is applied systemically. TRAIL is a protein that triggers apoptosis in cells. It has been shown that resistance to TRAIL-mediated apoptosis can occur differently in cells that develop TRAIL resistance. TRAIL is a protein that triggers apoptosis in cells. In the primary mechanism of TRAIL, this protein, which binds to cell death receptors, triggers apoptosis in two different ways. First, when it binds to the DR4/DR5 receptors, FADD activates and changes the ratio of Bax and Bcl-2 from mitochondrial proteins and causes the cell to die with the activation of caspase-3 and caspase-9. Another way for TRAIL is to trigger cell death by activating TRADD and TRAF-2, increasing the expression of caspase-8 through NF-kB activation [20]. Moreover, it is reported that miRNAs contribute to this apoptosis pathway and play a role in the TRAIL signaling mechanism [21]. It has been shown that resistance to TRAIL-mediated apoptosis can occur differently in cells that develop TRAIL resistance TRAIL drug resistance is evaluated chiefly as dysfunctions of DR4 and DR5 death receptors due to mutations, loss of Bax and Bcl-2 function, decreased caspase activations, mitogen- activated protein (MAP) kinases defects, and the functioning of various nuclear factors [22, 23, 24]. It is known that TRAIL develops resistance in glioblastoma, and new studies show that different pathological conditions are exhibited due to the development of this resistance [25].
Preclinical studies have shown that TRAIL administration does not have a toxic effect on healthy cells but has a tumoricidal activity on GBM [26, 27]. In addition, some studies have reported that the combined use of TRAIL and bortezomib, which are used in GBM treatment, induced apoptosis in GBM cancer cells and patient-derived primary GBM stem cells [28]. On the other hand, it is thought that combination therapies or new nanocarriers developed by TRAIL can be resisted [29]. Not only the chemotherapeutics used but also the exosomes released from cancer cells contribute to the development of drug resistance in cancer. Exosomes are small vesicles found in blood, urine, saliva, and other body fluids that contain many biomolecules secreted by cells. Exosomes released in the microenvironment of cancer cells enter the circulation and have outstanding contributions to the development of drug resistance in heterogeneous nature of tumor [30]. A study showed that exosomes released by lung cancer cells, which developed resistance to cisplatin, activate miRNAs in other cells and induce drug resistance, which changes mTOR expression levels [31]. It has been reported that exosomes secreted by breast cancer cells resistant to azithromycin provide drug resistance by rearranging the cell cycle and genes associated with apoptosis [32]. It has been reported that exosomes derived from colon cancer cells resistant to cetuximab support drug resistance by increasing Akt phosphorylation levels in cells [19]. It has been shown that exosomes released from sorafenib-resistant liver cancer cells contribute to resistance by modulating the TGF-β signaling pathway via lncRNAs [33]. It has been shown that exosomes induce sunitinib resistance in kidney cancer cells [34]. In this study, we examined the effect of exosomes released from TRAIL resistance glioblastoma cells on healthy cells. We detected the effects of the contents of exosomes on CXCR4, CXCL-12, PAK4, and 14-3-3 β proteins that cause migration in cancer cells but have low expression in healthy cells. The PAK4 protein, a member of the six known p21- activated kinases (PAK) protein family, impairs DNA repair in glioblastoma and contributes to migration and resistance development [35]. 14-3-3 β is an oncogenic protein and has been reported to have the potential to be a new therapeutic target in glioblastoma. Because 14-3-3 β plays a role in tumor growth and progression [36]. Reports that CXCR4, a receptor of the chemokine family that contributes to the spread and proliferation of cancer cells, and its ligand, CXCL-12, are a novel therapeutic target in glioblastoma patients [37].
Our study shows that exosomes released from TRAIL- resistant cells increase CXCR4, CXCL-12, and PAK4 levels in healthy cells and do not cause changes in 14-3-3 β oncogenic protein. Released exosomes, miRNAs that develop due to drug resistance, and proteins that play a role in migration can cause changes in healthy cells by infiltrating various nucleic acids. Since exosomes can easily cross the blood-brain barrier, they can transfer signals related to drug resistance to other cells and cause the cells to lose their natural forms.
Conclusion
Finally, our study showed that exosomes released from drug-resistant cancer cells act on healthy cells and affect proteins that contribute to cancer metastasis and spread. Health cells respond like cancer cells, and the deterioration of their systems shows that high concentrations of exosomes cannot be eliminated inside the cell and may affect healthy cells in cancer patients.
Conflict of Interest
We have no conflict of interests to disclose and the manuscript has been read and approved by all named authors.
Acknowledgment
This work was supported by and Health Institutes of Turkey (TUSEB-4381) and Aydin Adnan Menderes University Research Grant (ADU-TPF-21050).
References
-
Chen C, Sun M, Wang J, Su L, Lin J, et al. (2021) Active cargo loading into extracellular vesicles: Highlights the heterogeneous encapsulation behaviour. J Extracell vesicles 10(13): e12163.
-
Kalluri R, LeBleu VS (2020) The biology, function, and biomedical applications of exosomes. Science 367(6478): eaau6977.
-
Wu P, Zhang B, Ocansey DKW, Xu W, Qian H (2021) Extracellular vesicles: A bright star of nanomedicine. Biomaterials 269: 20467.
-
Lu M, Xing H, Xun Z, Yang T, Zhao X, et al. (2018) Functionalized extracellular vesicles as advanced therapeutic nanodelivery systems. Eur J Pharm Sci 121: 34-46.
-
Schindler C, Collinson A, Matthews C, Pointon A, Jenkinson L, et al. (2019) Exosomal delivery of doxorubicin enables rapid cell entry and enhanced in vitro potency. PLoS One 14(3): e0214545.
-
Cenik M, Abas B, Kocabıyık B, Demirbolat G, Cevik O (2021) Development of a New Drug Delivery System from HELA-Derived Exosomes and the Effect of Docetaxel- Loaded Exosomes on Mitochondrial Apoptosis. J Pharm Innov 17: 931-939.
-
Walker S, Busatto S, Pham A, Tian M, Suh A, et al. (2019) Extracellular vesicle-based drug delivery systems for cancer treatment. Theranostic 9(26): 8001-8017.
-
Abas BI, Demirbolat GM, Cevik O (2022) Wharton jelly-derived mesenchymal stem cell exosomes induce apoptosis and suppress EMT signaling in cervical cancer cells as an effective drug carrier system of paclitaxel. PLoS One 17(9): e0274607.
-
Tamimi A, Juweid M, Vleeschouwer SD(2017) Epidemiology and Outcome of Glioblastoma.
-
Dymova MA, Kuligina E V, Richter VA (2021) Molecular Mechanisms of Drug Resistance in Glioblastoma. International Journal of Molecular Sciences 22(12): 6385.
-
Weil S, Osswald M, Solecki G, Grosch J, Jung E, et al. (2017) Tumor microtubes convey resistance to surgical lesions and chemotherapy in gliomas. Neuro Oncol. 19(10): 1316-1326.
-
Haumann R, Videira JC, Kaspers GJL, van Vuurden DG, Hulleman E (2020) Overview of Current Drug Delivery Methods Across the Blood–Brain Barrier for the Treatment of Primary Brain Tumors. CNS Drugs 34(11): 1121-1131.
-
Terasaki T, Pardridge WM (2000) Targeted drug delivery to the brain; (blood-brain barrier, efflux, endothelium, biological transport). J Drug Target 8(6): 353-355.
-
Elliott RO, He M (2021) Unlocking the Power of Exosomes for Crossing Biological Barriers in Drug Delivery. Pharmaceutics 13(1): 122.
-
Hersh SD, Wadajkar SA, Roberts BN, Perez GJ, Connolly PN, et al. (2016) Evolving Drug Delivery Strategies to Overcome the Blood Brain Barrier. Current Pharmaceutical Design 22(9): 1177-1193.
-
Nam L, Coll C, Erthal LCS, De la Torre C, Serrano D, et al. (2018) Drug Delivery Nanosystems for the Localized Treatment of Glioblastoma Multiforme. Materials 11(5): 779.
-
Çoruh I, Çevik Ö, Yelekçi K, Djikic T, Küçükgüzel ŞG (2018) Synthesis, anticancer activity, and molecular modeling of etodolac-thioether derivatives as potent methionine aminopeptidase (type II) inhibitors. Arch Pharm (Weinheim) 351(3-4): 1700195.
-
Cevik O, Acidereli H, Turut FA, Yildirim S, Acilan C (2020) Cabazitaxel exhibits more favorable molecular changes compared to other taxanes in androgen-independent prostate cancer cells. J Biochem Mol Toxicol 34(9): e22542.
-
Acidereli H, Turut FA, Cevik O (2021) Acetylation of interferon regulatory factor-5 suppresses androgen receptor and downregulates expression of Sox2. Cell Biochem Funct 39(5): 667-678.
-
Wang F, Lin J, Xu R (2014) The Molecular Mechanisms of TRAIL Resistance in Cancer Cells: Help in Designing New Drugs. Current Pharmaceutical Design 20(42): 6714- 6722.
-
Zhang X, Zhang X, Hu S, Zheng M, Zhang J, et al. (2017) Identification of miRNA-7 by genome-wide analysis as a critical sensitizer for TRAIL-induced apoptosis in glioblastoma cells. Nucleic Acids Res 45(10): 5930-5944.
-
Zhang L, Fang B (2005) Mechanisms of resistance to TRAIL-induced apoptosis in cancer. Cancer Gene Ther 12(3): 228-237.
-
Mitsiades CS, Treon SP, Mitsiades N, Shima Y, Richardson P, et al. (2001) TRAIL/Apo2L ligand selectively induces apoptosis and overcomes drug resistance in multiple myeloma: therapeutic applications. Blood 98(3): 795- 804.
-
Frese S, Pirnia F, Miescher D, Krajewski S, Borner MM, et al. (2003) PG490-mediated sensitization of lung cancer cells to Apo2L/TRAIL-induced apoptosis requires activation of ERK2. Oncogene 22(35): 5427-5435.
-
Deng L, Zhai X, Liang P, Cui H (2021) Overcoming TRAIL Resistance for Glioblastoma Treatment. Biomolecules 11(4): 572.
-
Fulda S, Wick W, Weller M, Debatin KM (2002) Smac agonists sensitize for Apo2L/TRAIL- or anticancer drug- induced apoptosis and induce regression of malignant glioma in vivo. Nat Med 8(8): 808-815.
-
Eisele G, Weller M (2013) Targeting apoptosis pathways in glioblastoma. Cancer Lett 332(2): 335-345.
-
Unterkircher T, Cristofanon S, Vellanki SHK, Nonnenmacher L, Karpel-Massler G, et al. (2011) Bortezomib Primes Glioblastoma, Including Glioblastoma Stem Cells, for TRAIL by Increasing tBid Stability and Mitochondrial Apoptosis. Clin Cancer Res 17(12): 4019-4030.
-
Panner A, Crane CA, Weng C, Feletti A, Fang S, et al. (2010) Ubiquitin-Specific Protease 8 Links the PTEN- Akt-AIP4 Pathway to the Control of FLIPS Stability and TRAIL Sensitivity in Glioblastoma Multiforme. Cancer Res 70(12): 5046-5053.
-
Zhong Y, Li H, Li P, Chen Y, Zhang M, et al. (2021) Exosomes: A New Pathway for Cancer Drug Resistance. Front Oncol 11: 743556.
-
Qin X, Yu S, Zhou L, Shi M, Hu Y, Xu X, et al. (2017) Cisplatin-resistant lung cancer cell-derived exosomes increase cisplatin resistance of recipient cells in exosomal miR-100-5p-dependent manner. International journal of nanomedicine 12: 3721-3733.
-
Yu D, Wu Y, Zhang X, Lv M, Chen W, et al. (2016) Exosomes from adriamycin-resistant breast cancer cells transmit drug resistance partly by delivering miR-222. Tumor Biol 37(3):3227-3235.
-
Takahashi K, Yan IK, Kogure T, Haga H, Patel T (2014) Extracellular vesicle-mediated transfer of long non- coding RNA ROR modulates chemosensitivity in human hepatocellular cancer. FEBS Open Bio 4(1): 458-467.
-
Qu L, Ding J, Chen C, Wu ZJ, Liu B, et al. (2016) Exosome- Transmitted lncARSR Promotes Sunitinib Resistance in Renal Cancer by Acting as a Competing Endogenous RNA. Cancer Cell 29(5): 653-668.
-
Blankenstein LJ, Cordes N, Kunz-Schughart LA, Vehlow A (2022) Targeting of p21-Activated Kinase 4 Radiosensitizes Glioblastoma Cells via Impaired DNA Repair. Cells 11(14): 2133.
-
Liu L, Liu Z, Wang H, Chen L, Ruan F, et al. (2017) 14-3-3β exerts glioma-promoting effects and is associated with malignant progression and poor prognosis in patients with glioma. Exp Ther Med 15(3): 2381-2387.
-
Ehtesham M, Mapara KY, Stevenson CB, Thompson RC (2009) CXCR4 mediates the proliferation of glioblastoma progenitor cells. Cancer Lett. 274(2): 305-312.
- The Muculent Bleb-Mucinous Cystic Neoplasm-Hepatobiliary Region
- Insulin Sensitizers as Anti-Aging Agents: Unveiling Synergies with Albumin, GLP-1RA, Klotho Protein, and Metformin in the Quest to Combat Aging
- Reprogramming of GLP-1 Response at Prediabetes for the Prevention of Type 2 Diabetes: The Role of Albumin and GLP-1 Receptor Agonists
- The Mingled Allies-Combined Hepatocellular Carcinoma and Cholangiocarcinoma
- Compilation and Embodiment-Leydig Cell Tumour Testis
- Glucolipotoxicity: A Novel Different Perspective on the Causes of Cancer