GHRL Gene Polymorphisms in Early Onset Psoriasis: Molecular and In silico Analyses
Background: Psoriasis is an inflammatory skin disease. Ghrelin protein exhibits an anti-inflammatory effect by inhibiting the TNF-α/NF-κB pathway in psoriasis. The aim of this study was to investigate whether the GHRL gene rs27647 and rs34911341 polymorphisms are associated with susceptibility to generalized Early Onset Psoriasis (EOP). Material and Methods: The study group consisted of 80 EOP patients and 70 unrelated healthy controls. GHRL polymorphisms were determined using the polymerase chain reaction (PCR)-restriction fragment length polymorphism method. In addition, we determined other genes with which the GHRL gene interacts, and the effect on protein function and protein stability for rs34911341. We used web-based tools for all in silico analyses. Results: Our results have shown GHRL gene rs34911341 polymorphism GA genotype is associated with an increased risk of EOP (p=0.000). Also, there was a significant difference between the two groups with regard to the allele frequency of rs34911341 polymorphism (p=0.000). We have shown the prediction of Gene-gene interaction that GHRL is co-expressed with 18 genes, physical interaction with 16 genes and co-localization with 9 genes. We determined deleterious of rs34911341 in 7 tools and revealed that rs34911341 decreased protein stability significantly. Conclusion: The GHRL rs34911341 polymorphism genotype and allele frequencies of the EOP patients differ significantly from those of healthy controls. We obtained in silico analysis results that support molecular data for rs34 polymorphism in the GHRL gene. This may be responsible for explaining chronic inflammation in the pathogenesis of EOP.
Introduction
Psoriasis is a skin disease with pathological features in which chronic inflammation is seen [1]. Although it varies among ethnic groups, its prevalence has been reported as
2% and the exact cause is unknown, genetic predisposition seems to be the main cause [2, 3]. More than 20 gene loci for psoriasis have been identified. The locus with the strongest effect is the major Psoriasis susceptibility 1 (PSORS1) locus [4]. Although psoriasis typically appears on the skin, the inflammation is not limited to the skin. Cytokines released from different T cell subtypes such as interleukin-6 (IL-6), IL- 12, IL-18, tumour necrosis factor-alpha (TNF-α), interferon- gamma (IFN-γ) ensure the continuity of inflammation. Also, mast cells and neutrophils contribute to inflammation by secreting IL-17 [5, 6]. Released pro-inflammatory cytokines make inflammation systemic.
The Ghrelin gene (GHRL) is located on chromosomes 3p25-26 and Ghrelin is secreted by X/A cells in the stomach [7]. Ghrelin, a 28 amino acid peptide, inhibits pro-inflammatory signals through its action on T cells and monocytes [5, 8]. Ghrelin prevents inflammation by reducing the release of TNF-α, IL-6 and IL-1β inflammatory cytokines [9]. It is also known to inhibit nuclear factor kappa-B (NF- κB) and NLRP3 inflammatory signaling pathways [10]. Ghrelin exhibits an anti-inflammatory effect by suppressing the TNF-α / NF-kB pathway in psoriasis. Besides, In addition, the anti-inflammatory function mechanism of ghrelin has been determined. Ghrelin binds to GHSR1a and inhibits the release of inflammatory cytokines. Thus, it protects the body from inflammation [11, 12] (Figure 1).
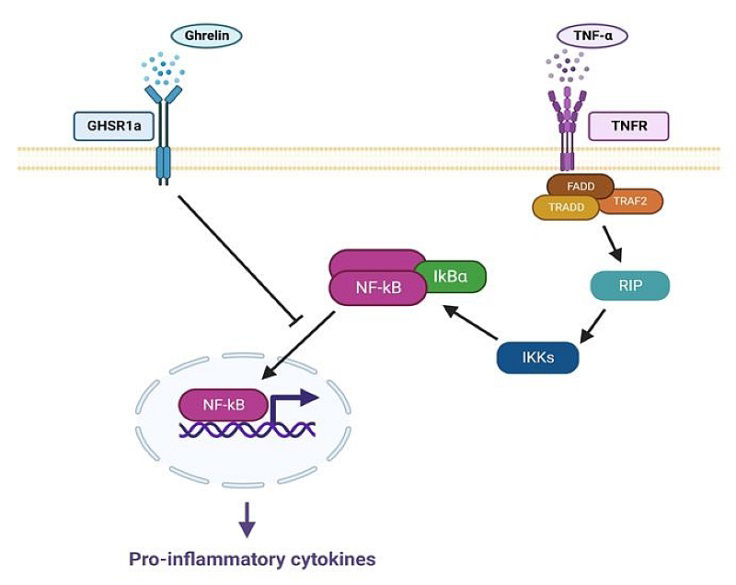
Figure 1: Mechanism of the effect of ghrelin for psoriasis. IKKs activated by TNF-α inhibit IκBα. When IκBα is inhibited, NF-κB is released and acts as a transcription factor, increasing the expression of inflammatory cytokines. Ghrelin binds to GHS R1a, inhibits NF-κB from reaching the nucleus, and inhibits the expression of pro-inflammatory molecules.
Missense variants directly affect proteins. The promoter and intron regions of the gene are not converted into proteins. However, since the promoter is an important region in transcription and intron regions are important in the splicing process, it affects the expression of the gene. Therefore, in addition to missense variants, variants in the non-coding region also directly affect gene expression. rs27647 is located in the promoter region of the GHRL gene, while rs34911341 (R51Q) is located in the exon. Previous studies reveal the relationship between rs27647 and rs34911341 polymorphisms in the GHRL gene with different diseases. Although the effect of the ghrelin hormone anti-inflammatory is known, to the best of our knowledge, no research has been conducted on GHRL gene polymorphism in patients with EOP. The aim of this study was to determine whether the GHRL gene polymorphisms (rs27647 and rs34911341) are associated with psoriasis risk and characteristics and we were to determine the effect of genetic variants on the protein by in silico approaches.
Material and Methods
Study Population
The study was approved by Mugla Sitki Kocman University Medical Faculty Ethics Committee (Ethics approval number 24/12/2020, 16/VI) was conducted according to the Declaration of Helsinki and informed consent was obtained from all participants. A total of 150 volunteers, consisting of 80 patients diagnosed with EOP and 70 healthy controls in Mugla province (Southwest of Turkey) were included in our study. Gender, age, family history, EOP period, body mass index (BMI), EOP severity, betatrophin, bisphenol A, urocortin and vitamin D levels of the patients were examined. The severity of EOP was determined using the Psoriasis Area Severity Index (PASI), pregnant and lactating women and individuals under the age of 18 were not included in our study. Anthropometric measurements of our sample group were recorded. The duration of psoriasis was determined according to the reports of the patients.
DNA Extraction and Genotyping
Venous blood samples (2 mL) were collected in Vacutainer tubes containing sodium/potassium EDTA. DNA was extracted with a Hibrigen Blood DNA Isolation Kit (MG-KDNA-02-250; Hibrigen Biotechnology R&D Industry and Trade Inc., Gebze, Kocaeli, Turkey). The rs27647 and rs34911341 SNPs in the GHRL gene were determined by the Polymerase chain reaction (PCR)-restriction fragment length polymorphism (RFLP) method. PCR was performed in a 25-µL volume with 100 ng of DNA 2X Taq Master Mix (MG-TAQMX-01-80; Hibrigen Biotechnology R&D Industry and Trade Inc., Gebze, Kocaeli, Turkey) using an automated thermal cycler (SimpliAmp Thermal Cycler, Thermo Fisher Scientific, USA).
The primer sequences and PCR annealing temperature are listed (Table 1). PCR products were digested with the appropriate restriction enzymes for rs27647 DraI also for rs34911341 SacI. Digestion conditions, restriction enzymes and restriction fragment sizes are shown (Table 2).
| Gene | Polymorphism | Primers | Temperature of annealing | PCR product sizes |
|---|---|---|---|---|
| GHRL | rs27647 rs34911341 | P1 P2 P3 P4 | 58 °C 64 °C | 200 bp 618 bp |
Table 1: Oligonucleotides, PCR conditions and PCR product sizes used for polymorphism of GHRL gene.
P1: 5’ AGAGAGGTTAAACGGACGGG 3’, P2: 5’ GAATGTACATCAGCCCCACG 3’ P3: 5’ CACAGCAACAAAGCTGCACC 3’, P4: 5’ AAGTCCAGCCAGAGCATGCC 3’ Table 1: Oligonucleotides, PCR conditions and PCR product sizes used for polymorphism of GHRL gene.
| Gene | Polymorphism | Restriction enzyme | Digestion conditions | Restriction fragment sizes |
|---|---|---|---|---|
| GHRL | rs27647 | DraI | 37°C, 3h | C allele: 200 bp |
| T allele: 150 bp | ||||
| rs34911341 | SacI | 37°C, 3h | A allele: 618 bp | |
| G allele: 455 bp |
Table 2: Oligonucleotides, PCR conditions and PCR product sizes used for polymorphism of GHRL gene.
Table2: Digestion conditions, restriction enzymes and restriction fragment sizes.
PCR products were separated using 2.5% agarose gels containing 0.5 mg/mL ethidium bromide (120V, 40-50 min). We used a 100-bp DNA ladder (Fermentas Vilnius, Lithuania) as the size standard for each gel lane. The gel was visualized under ultraviolet light using a gel electrophoresis visualizing system (Cleaver Scientific Ltd., Clear View UV Trans illuminator, Rugby, UK).
Statistical analysis
All of the frequency and statistical calculations were performed using SPSS, ver. 22.0 (IBM Corporation, Armonk, NY, USA). Chi-square analysis was used to compare frequencies of the GHRL genotypes and alleles between the patients in the EOP and control groups. Risk analyses were determined by logistic regression method. Odds ratios (OR) for genotypes were calculated and 95% confidence ranges (CI) were determined. Independent samples t-test was used to compare continuous variables. Results with a p-value of <0.05 were considered statistically significant. In silico Analyses In order to better understand the role of rs34911341, we evaluated the effect of amino acid alteration on protein stability and protein function.
Prediction of Deleterious missense SNP rs34911341
The tool PredictSNP1.0 (http://loschmidt.chemi. muni.cz/predictsnp1/) was used to show the deleterious effect of missense variant rs34911341 on the protein. Predict SNP 1.0 is a web-based tool that shows the results of 9 different SNP effect predictors such as Predict SNP, PolyPhen-1 (Polymorphism Phenotyping v1), PolyPhen-2 (Polymorphism Phenotyping v2), SIFT (Sorting Intolerant from Tolerant), MAPP (Multivariate Analysis of Protein Polymorphism), PhD-SNP (Predictor of Human Deleterious Single Nucleotide Polymorphisms), SNAP (Screening for Nonacceptable Polymorphisms), PANTHER (Protein Analysis Through Evolutionary Relationships), and SNP Analyser prediction tools [13, 14]. We determine the deleterious effect of rs34911341 on the protein was determined SIFT, PolyPhen-1, PolyPhen-2, MAPP, SNAP, PANTHER, and Predict SNP prediction tools.
SIFT evaluates whether the mutation is deleterious based on the physical characteristics and sequence homology of the amino acids. This web server uses multiple sequence alignment for each query. The tolerance index in the SIFT is low (<0.05), as the probability of tolerating the change for conserved amino acids is low [15, 16]. Polyphen-1 uses a set of expert empirical rules. Polyphen-2 uses multiple sequence alignment [17]. MAPP evaluates the physicochemical differences in each column in protein sequence alignment [18]. PhD-SNP uses a support vector machine (SMV), while SNAP uses a neural network [19, 20]. PANTHER determines the effect of nsSNP on protein based on evolutionary preservation specific to the position of amino acids [21]. PredictSNP calculates an estimate based on the confidence score of each tool.
Prediction of Change in Protein Stability
The change in protein stability was analysed with the MUpro tool (http://mupro.proteomics.ics.uci.edu/) [22]. This tool uses SMV and neural networks machine learning methods. This protein calculates a score between -1 and 1 as the confidence of the prediction. If the confidence score was <0, the mutation has decreased protein stability. But if the confidence score was >0, the mutation has increased protein stability.
Results
Associations of GHRL genotypes with EOP
DNA samples from 80 EOP patients and 70 healthy controls were analysed for GHRL gene rs27647 and rs34911341 polymorphisms. The patients’ demographic and clinical characteristics were represented (Table 3). The genotype and allele distributions for rs27647 and rs3491134 were shown in (Table 4).
- GHRL gene of its association with other genes in order to predict was used, GeneMANIA (https://genemania.org/)
- (accessed on 27 August 2022). The prediction of gene-gene interaction predictor GeneMANIA is based on the basis of pathways, co-localization, co-expression, protein domain
- Healthy controls n (%)
- Cases n (%) χ² p value
- OR (95 % CI)
- CC
- 20 (28.5)
- 12 (15)
- Reference
- Genotype rs27647
- CT
- 26 (37.1)
- 51 (63.7)
- 0.401
- 0.162 (0.041-0.370)
- TT
- 24 (34.3)
- 17 (21.3)
- 0.180 (0.055-0.541)
- Allele rs27647
- C
- 66 (47.15)
- 75 (46.8)
- 0.35
- T
- 74(52.85)
- 85(53.2)
- AA
- -
- 8 (10)
- Reference
- Genotype rs34911341
- GA
- 29 (41.52)
- 58 (72.5)
- 0
- 0.642 (0.262-3.392)
- GG
- 41 (58.48)
- 14 (17.5)
- 0.125 (0.041-0.304)
- Allele rs34911341
- A
- 29(20.7)
- 74(46.8)
- G
- 111(79.3)
- 86(54.2)
- 0
Table 3: Distributions of GHRL genotypes and allele frequencies in EOP patients.
| Clinical characteristics | Mean±SD | p value | |
|---|---|---|---|
| EOP (n=80) | Control (n=70) | ||
| Age | 33.51±10.2 | 34.62±11.3 | ˃0.05 |
| Gender, n(%) | |||
| Male | 46 | 38 | |
| Female | 34 | 32 | |
| Family story | |||
| Yes | 42 | ||
| No | 36 | ||
| Psoriatic nail | |||
| Yes | 56 | ||
| No | 24 | ||
| BMI | 23.7±11.2 | 23.4±9 | ˃0.05 |
| PASI˂3 | 30 | ||
| PASI (3-10) | 28 | ||
| PASI˃10 | 22 |
Table 4: Distributions of GHRL genotypes and allele frequencies in EOP patients.
All genotype frequencies observed in the control group were in Hardy–Weinberg equilibrium (p>0.05). The respective frequencies of the CC, CT and TT genotypes for the rs27647 GHRL polymorphism were 15%, 63.7%, and 21.3% in the control group and 28.5%, 37.1%, and 34.3% in the EOP group. Considering the genotype frequencies, the rs27647 polymorphism was not significant between the two groups (p=0.401) (Table 4).
The GA and GG genotype distributions of the rs34911341 GHRL polymorphism were 41.52%, and 58.48% for the controls group and was no AA genotype. The AA, GA and GG genotype distributions of rs34911341 GHRL polymorphism were 10%, 72.5% and 17.5% for the EOP patients. The GA genotype was higher than the GG genotype between the two groups. So, the GA genotype rs34911341 was the risk genotype for EOP (p=0.00) (Table 4). The allele frequency of rs27647 was not significantly different between the two groups. The A and G allele frequencies of rs34911341 were significantly different between the two groups. According to the results, the G allele was higher in the EOP patient group compared to the control group.
Prediction of Deleterious missense SNP rs34911341
The deleterious effect of rs34911341 (R51Q) has also been demonstrated in silico. Analyses were made with 8 different tools using PredictSNP 1.0. As a result, deleterious effects were detected in all tools except the PhD-SNP tool (Table 5).
| SNP | Tools | Confidence score |
|---|---|---|
| rs34911341 | Predict SNP | 0.76 (Deleterious) |
| PolyPhen-1 | 0.74 (Deleterious) | |
| PolyPhen-2 | 0.81 (Deleterious) | |
| SIFT | 0.79 (Deleterious) | |
| MAPP | 0.77 (Deleterious) | |
| PhD-SNP | 0.68 (Deleterious) | |
| SNAP | 0.72 (Deleterious) | |
| PANTHER | 0.66 (Deleterious) |
Table 5: The expected accuracy results of the rs34911341 of GHRL in PredictSNP1.0 and integrated tools.
Prediction of Change in Protein Stability
We analysed the rs34911341 (R51Q) predicted deleterious mutant with help of the MUpro web server. Confidence scores for the SVM and NN methods were calculated as -0.85 and -0.70 (kcal/mol), respectively. The rs34911341 (R51Q) polymorphism decreases protein stability. The results of our in silico analyses and the results of the molecular analysis for rs34911341 (R51Q) are compatible. rs34911341 (R51Q) is a missense variant and can have a serious effect on the phenotype.
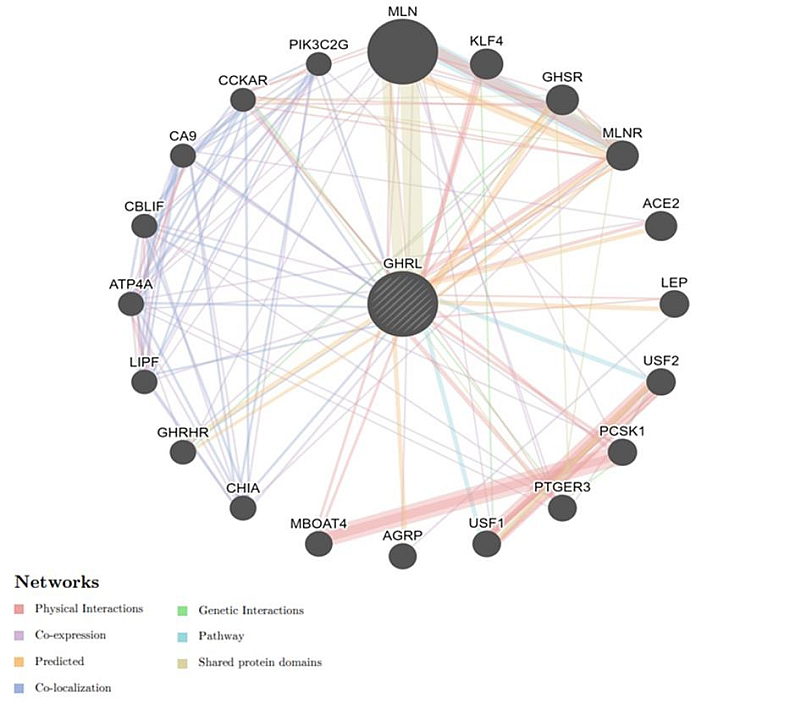
1.1 Prediction of Gene-Gene Interactions Our findings revealed that GHRL is co-expressed with 18 genes (MLN, GHSR, MLNR, ACE2, LEP, USF2, PCSK1, PTGER3, USF1, AGRP, CHIA, GHRHR, LIPF, ATP4A, CBLIF, CA9, CCKAR, PIK3C2G), shared a domain with only 7 genes (CCKAR, GHSR, MLN, MLNR, USF1, PTGER3, USF2), physical interaction with 16 genes (MLN, KLF4, GHSR, MLNR, ACE2, LEP, USF2, PCSK1, PTGER3, USF1, MBOAT4, LIPF, ATP4A, CBLIF, CA9, CCKAR), and co-localization with 9 genes (MLNR, CHIA, GHRHR, LIPF, ATP4A, CBLIF, CA9, CCKAR, PIK3C2G) (Figure 2).
Discussion
In our study, we examined for the first time the GHRL gene rs27647 and rs34911341 polymorphisms in EOP. According to the results obtained, the GA genotype of rs34911341 polymorphism increased the risk of EOP (p=0.00). The A allele is a risk factor for rs3491134 polymorphism. But we determined genotypes and allele frequency were no significant differences between the two groups for rs27647 polymorphism. In addition, in silico analysis was performed for the missense variant rs34911341 polymorphisms were predicted to be deleterious for 7 tools and predicted to reduce protein stability. This result is important for future studies with the GHRL gene. In addition, According to the estimates of GENEMANIA, the GHRL gene interacts with 20 different genes. Mutations or polymorphisms (rs27647 and rs34911341 for this research) in the GHRL gene can cause various diseases by affecting the functioning of other linked genes. Molecularly, the data we obtained for rs34911341 supports the results of in silico analysis.
Many studies of GHRL gene SNPs in different populations have been reported [25, 26, 27, 28, 29]. The Leu72Met (rs696217) polymorphism has been shown to be associated with Type 2 Diabetes Mellitus (T2DM) in the Chinese population. According to the study, individuals carrying the Met72 allele have a higher risk of T2MD [25]. In a study in a Mexican population, the 72Met allele was shown to have a protective effect on T2MD [30]. The association between the GHRL gene SNPs rs26802, rs10490816, and rs696217 and metabolic syndrome has been evaluated. It was concluded that SNPs do not pose a risk for metabolic syndrome [26]. SNPs rs27647, rs26802, rs696217 (Leu72Met) and rs4684677 were associated with lower systolic and diastolic blood pressure in the Finnish population [28, 31]. In another study on the Finnish population, the rs34911341 (R51Q) 51Gln allele was identified as a risk factor for hypertension and type 2 diabetes [29]. The association of the rs34911341 (R51Q) SNP with hypertension has also been shown in studies in the Caucasian population. The rs27647 and rs35683 SNPs in the GHRL gene were associated with a reduced risk of colorectal cancer, while the rs26312, rs26802 and rs27647 SNPs were associated with cerebral infarction [32]. Consequently, GHRL gene polymorphisms were associated with many diseases. In addition, the anti-inflammatory effect of the GHRL gene is known and its connection with psoriasis was reported [12, 33].
The TNF-α/NF-κB signalling pathway is the main mediator in the pathogenesis of psoriasis [1]. It was shown that TNF-α increases the expression of pro-inflammatory cytokines by the effect of reactive oxygen species [34]. GHRL provides inhibition of the TNF-α/NF-κB signalling pathway. GHRL modulation may be an important target in treating inflammatory skin diseases. The strong relation between rs34911341 polymorphism and EOP is a guide for future studies in elucidating the pathogenesis of EOP and increasing treatment possibilities. In silico analyses clearly demonstrate the disease potential of the GHRL gene. In addition, other genes that may be affected by possible mutations in the GHRL gene are important and different targets for EOP. This study has three main limitations; The first limitation is the small number of participants. We anticipated that recruitment might not meet targets. However, also because the condition is rare, the findings of this study offer new, potentially useful information for this patient population.
Conclusion
This is the first study to examine the association between GHRL gene polymorphisms and EOP. Our results show that the rs34911341 polymorphism of the GHRL might be associated with EOP. Although only the rs34911341 polymorphism was found to be significant, the molecular mechanisms underlying EOP need to be elucidated with larger sample sizes. In addition, we have revealed many genes related to the GHRL gene. The association of these genes with the pathogenesis of psoriasis could be investigated. The molecular data we obtained supports the in silico data. Signal that you are wrapping up your limitations: “Finally, we could not control for every possible lifestyle factor, and the observational nature of this design leaves the possibility of residual confounding.” We showed that the rs34911341 polymorphism, with which we obtained significant results, was pathogenic in 8 vehicles and reduced protein stability. The missense variant rs34911341 is a variant that should be investigated in detail for different diseases due to its pathogenic effects.
Conflict of interest
The authors declare there are no competing interests.
Author Contributions
Melih Furkan Turkmenoglu: Investigation, Writing- Original Draft, Visualization; Suzan Demir Pektas: Conceptualizaton, Resources, Investigation, Writing-Original Draft, Visualization; Aysegul Demirtas Bilgic: Investigation, Writing-Original Draft, Visualization; Cilem Ozdemir: Investigation, Writing-Original Draft, Visualization; Tuba Gokdogan Edgunlu: Conceptualizaton, Resources, Writing- Review and Editing, Supervision, Project Administration.
Funding Statement
This paper was supported by ‘Scientific and Technological Research Council of Turkey (TUBITAK) under Grant No: 1919B012003657
References
-
Pektas SD, Dogan G, Edgunlu TG, Karakas-Celik S, Ermis E, et al. (2018) The Role of Forkhead Box Class O3A and SIRT1 Gene Variants in Early-Onset Psoriasis. Indian J Dermatol 63(3): 208-214.
-
Rendon A, Schakel K (2019) Psoriasis Pathogenesis and Treatment. Int J Mol Sci 20(6): 1475.
-
Christophers E (2001) Psoriasis--epidemiology and clinical spectrum. Clin Exp Dermatol 26(4): 314-320.
-
Tawfik NZ, Abdallah HY, Hassan R, Hosny A, Ghanem DE, et al. (2022) PSORS1 Locus Genotyping Profile in Psoriasis: A Pilot Case-Control Study. Diagnostics (Basel) 12(5): 1035.
-
Ucak H, Demir B, Cicek D, Erden I, Aydinet S, et al. (2014) Metabolic changes and serum ghrelin level in patients with psoriasis. Dermatol Res Pract 2014: 175693.
-
Liang Y, Sarkar MK, Tsoi LC, Gudjonsson JE (2017) Psoriasis: a mixed autoimmune and autoinflammatory disease. Curr Opin Immunol 49: 1-8.
-
Kojima M, Kangawa K (2005) Ghrelin: structure and function. Physiol Rev 85(2): 495-522.
-
Gerkowicz A, Pietrzak A, Szepietowski JC, Radej S, Chodorowska G (2012) Biochemical markers of psoriasis as a metabolic disease. Folia Histochem Cytobiol 50(2): 155-170.
-
Ma Y, Zhang H, Guo W, Yu L (2022) Potential role of ghrelin in the regulation of inflammation. FASEB J 36(9): e22508.
-
Han K, Wang QY, Wang CX, Luan SY, Tian WP, et al. (2018) Ghrelin improves pilocarpine‑induced cerebral cortex inflammation in epileptic rats by inhibiting NF‑κB and TNF‑α. Mol Med Rep 18(4): 3563-3568.
-
Li WG, Gavrila D, Liu X, Wang L, Gunnlaugsson S, et al. (2004) Ghrelin inhibits proinflammatory responses and nuclear factor-kappaB activation in human endothelial cells. Circulation 109(18): 2221-2226.
-
Qu R, Chen X, Hu J, Fu Y, Peng J, et al. (2019) Ghrelin protects against contact dermatitis and psoriasiform skin inflammation by antagonizing TNF-α/NF-κB signaling pathways. Sci Rep 9(1): 1348.
-
Elkhattabi L, Morjane I, Charoute H, Amghar S, Bouafi H, et al. (2019) _In silico_ Analysis of Coding/Noncoding SNPs of Human RETN Gene and Characterization of Their Impact on Resistin Stability and Structure. J Diabetes Res 2019: 4951627.
-
Bendl J, Stourac J, Salanda O, Pavelk A, Wiebenet ED, et al. (2014) PredictSNP: robust and accurate consensus classifier for prediction of disease-related mutations. PLoS Comput Biol 10(1): e1003440.
-
Ng PC, Henikoff S (2003) SIFT: Predicting amino acid changes that affect protein function. Nucleic Acids Res 31(13): 3812-3814.
-
Das KC, Hossain MU, Moniruzzaman M, Salimullah M, Akhteruzzaman S (2022) High-Risk Polymorphisms Associated with the Molecular Function of Human HMGCR Gene Infer the Inhibition of Cholesterol Biosynthesis. Biomed Res Int 2022: 4558867.
-
Ramensky V, Bork P, Sunyaev S (2002) Human non- synonymous SNPs: server and survey. Nucleic Acids Res 30(17): 3894-3900.
-
Mah JT, Low ES, Lee E (2011) _In silico_ SNP analysis and bioinformatics tools: a review of the state of the art to aid drug discovery. Drug Discov Today 16(17-18): 800-809.
-
Capriotti E, Calabrese R, Casadio R (2006) Predicting the insurgence of human genetic diseases associated to single point protein mutations with support vector machines and evolutionary information. Bioinformatics 22(22): 2729-2734.
-
Bromberg Y, Rost B (2007) SNAP: predict effect of non- synonymous polymorphisms on function. Nucleic Acids Res 35(11): 3823-3835.
-
Tang H, Thomas PD (2016) PANTHER-PSEP: predicting disease-causing genetic variants using position-specific evolutionary preservation. Bioinformatics 32(14): 2230-2232.
-
Cheng J, Randall A, Baldi P (2006) Prediction of protein stability changes for single-site mutations using support vector machines. Proteins 62(4): 1125-1132.
-
Ghosh P, Ghosh S, Behera B, Behera JK, Bhattacharya M, et al. (2021) In-silico analysis of non-synonymous SNPs of human LDLR gene and their impact on familial hypercholesterolemia. Gene Rep 23: 101127.
-
Warde-Farley D, Donaldson SL, Comes O, Zuberi K, Badrawi R, et al. (2010) The GeneMANIA prediction server: biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res 38(Web Server issue): W214-W220.
-
Li YY, Lu XZ, Yang XX, Wang H, Geng H, et al. (2019) GHRL Gene Leu72Met Polymorphism and Type 2 Diabetes Mellitus: A Meta-Analysis Involving 8,194 Participants. Front Endocrinol (Lausanne) 10: 559.
-
You Y, Yu Y, Wu Y, Rao W, Zhang Y, et al. (2017) Association Study between Ghrelin Gene Polymorphism and Metabolic Syndrome in a Han Chinese Population. Clin Lab 63(1): 175-181.
-
Mager U, Kolehmainen M, Lindström J, Eriksson JG, Valle TT, et al. (2006) Association between ghrelin gene variations and blood pressure in subjects with impaired glucose tolerance. Am J Hypertens 19(9): 920-926.
-
Pöykkö S, Ukkola O, Kauma H, Savolainen MJ, Kesäniemi YA (2003) Ghrelin Arg51Gln mutation is a risk factor for Type 2 diabetes and hypertension in a random sample of middle-aged subjects. Diabetologia 46(4): 455-458.
-
Rivera-Leon EA, Llamas-Covarrubias MA, Sánchez- Enríquez S, Martínez-López E, González-Hita M, et al. (2020) Leu72Met polymorphism of GHRL gene decreases susceptibility to type 2 diabetes mellitus in a Mexican population. BMC Endocr Disord 20(1): 109.
-
Berthold HK, Giannakidou E, Krone W, Trégouët DA, Gouni-Berthold I (2010) Influence of ghrelin gene polymorphisms on hypertension and atherosclerotic disease. Hypertens Res 33(2): 155-160.
-
Campa D, Pardini B, Naccarati A, Vodickova L, Novotny J, et al. (2010) Polymorphisms of genes coding for ghrelin and its receptor in relation to colorectal cancer risk: a two-step gene-wide case-control study. BMC Gastroenterol 10: 112.
-
Chen L, Zhao H, Shen J, Ji X (2020) Association Between Ghrelin Gene Polymorphism and Cerebral Infarction. Med Sci Monit 26: e924539.
-
Ozdemir M, Yüksel M, Gökbel H, Okudan N, Mevlitoğlu I (2012) Serum leptin, adiponectin, resistin and ghrelin levels in psoriatic patients treated with cyclosporin. J Dermatol 39(5): 443-448.
-
Hébert HL, Bowes J, Smith RL, McHugh NJ, Barker JNWN, et al. (2014) Polymorphisms in IL-1B distinguish between psoriasis of early and late onset. J Invest Dermatol 134(5): 1459-1462.
- Epithelioid Granuloma; 3cases with Different Clinical Features
- Advancing Representation in Dermatology Clinical Trials: Ethical, Scientific, and Regulatory Imperatives for Inclusion Across all Fitzpatrick Skin Types
- A Case of Atopic Dermatitis with Concurrent Psoriasis Vulgaris: Successful Treatment with Upadacitinib
- Innovation Lifting Eyeshadow: A Synthesis of Makeup and Optical Illusion
- Distinguishing Superficial Actinic Porokeratosis from Actinic Keratosis with UVF Dermoscopy: A Case Report
- High Mobility Group Box 1 (HMGB1) in Cutaneous Inflammation: An Immune Modulator Bridging Cellular Stress, Ferroptosis and Danger Signaling