Genetic Polymorphism of Endothelial Nitric Oxide Synthase in Sickle Cell Disease Patients in South-Eastern Nigeria
Background: Sickle cell disease (SCD) is an inherited blood disorder resulting from a point mutation of thymine (T) for adenine (A) at codon 6 of the gene for beta-globin chain of haemoglobin, leading to substitution of Valine for Glutamate at position 6 of the amino acid sequence, thereby shifting the isoelectric point of the protein. The single nucleotide polymorphism (SNP) (T-786C) in the 5’ promoter region which affects the expression of endothelial nitric oxide synthase (ENOS) gene is critical in determining the predisposition of humans to diseases including SCD. This work was therefore aimed at investigating the presence of SNP (T-786C) in the 5’ promoter region of ENOS gene in patients with SCD with respect to controls, visiting the University of Nigeria Teaching Hospital (UNTH) Ituku-Ozalla, Enugu State, south-Eastern Nigeria. Materials and Methods: The study recruited a total of 50 subjects including 20 SCD patients (SS), 15 sickle cell carriers (AS) and 15 healthy non-carriers (AA). DNA was extracted from venous blood samples; PCR amplification was done using specified primers to the region of polymorphism, and finally followed by HpaII restriction enzyme digestion. Results: The results obtained showed only TT genotype across all samples, and so could not establish any relationship between the SNP T-786C ENOS gene region and SCD. There was presence of the same genotype in both test and control groups. Conclusion: This research findings therefore suggest that T-786C ENOS gene polymorphism is not associated with SCD in south-Eastern Nigeria and thus cannot genetically influence the phenotypic outcome of the disease in the clime.
Introduction
Sickle cell disease (SCD) is a qualitative kind of haemoglobinopathy which affects the structure of the haemoglobin, the protein in red blood cells which delivers oxygen to all the cells of the body. This disease was discovered in 1910 [1, 2]. It is an inherited blood disorder resulting from a point mutation involving thymine (T) for adenine (A) (from a GAG to GTG) substitution at codon 6 of the gene for beta-globin chain of haemoglobin protein, leading to the substitution of Valine for Glutamate at position 6, thereby shifting the isoelectric point of the protein [3]. It is an autosomal recessive hereditary disease characterized by the presence of sickle-shaped red blood cells and accelerated haemolysis hence leading to anaemia. Individuals who have the S-gene in the homozygous state have sickle cell anaemia, while those who have it in combination with the normal A-gene (sickle cell carriers) usually do not show symptoms [4]. The sickled red blood cells are less flexible than normal ones, which results in micro-vascular occlusion leading to “crisis” that is characterized by episodes of severe pain, bone infarcts, leg ulcers, associated with increased susceptibility to secondary infections [4, 5]. The anaemia part of this disease is caused by the destruction of the red blood cells, because of their shape. Although the bone marrow attempts to compensate the shortfall in the number of red blood cells by creating new red blood cells, the production rate does not match the rate of destruction. Healthy red blood cells typically function for 90-120 days, but sickled red blood cells may only last for 10-20 days [6]. Signs of sickle cell anaemia usually begin in early childhood. The severity of symptoms of sickle cell anaemia varies from person to person [7].
According to global estimates, approximately 5% of the population has some type of haemoglobin variant, and more than 300,000 babies are born each year with haemoglobinopathies, with sickle cell anaemia being the most prevalent type [8, 9]. It is estimated that the prevalence of live births with the disease is 4.4% in the world, where the rates remain high on the continents of Africa, Southeast Asia and the Americas. The prevalence of sickle cell trait ranges from 10 to 45% in various parts of sub-Saharan Africa [10, 11, 12]. In Nigeria, carrier prevalence is about 20 to 30%. Sickle cell anaemia affects about 2 to 3% of the Nigerian population of more than 200 million [13].
The phenotypic diversity of SCD with different clinical outcomes seems to be modulated by polymorphisms in genes that are involved in inflammation, cell–cell interaction and modulators of oxidant injury and nitric oxide (NO) biology [14]. NO has properties that can impact every aspect of SCD, from decreasing platelet activation and adhesion receptor expression on the vascular endothelium, to decreasing vascular smooth muscle proliferation, limiting ischaemia- reperfusion injury, modulating endothelial proliferation, and regulating inflammation [15]. In vivo NO is synthesized during the enzymatic conversion of L-arginine to L-citrulline by three isoforms of nitric oxide synthase (NOS) enzyme, namely, neuronal NOS (nNOS or NOSI), inducible NOS (iNOS or NOSII), and endothelial NOS (ENOS or NOSIII) [16]. Endothelial (E) NOS, derived from vascular endothelium, is the most dominant form of these isoforms [17]. The level of NO in the body is linked to expression of ENOS gene [18]. The Single Nucleotide Polymorphism (SNP) (T-786C) in the 5’ promoter region affects the expression of ENOS gene. The T-786C allele binds the inhibitory transcription factor protein A1 resulting in a low mRNA level of ENOS and this reduces NO production and endothelial function [19]. Reduced endothelial NO bioavailability in SCD leads to activation of endothelial cell adhesion molecules besides platelet activation, which results into vascular occlusion and vasoconstriction [20]. Many published reports have suggested the involvement and otherwise of ENOS polymorphisms in the pathogenesis of sickle cell complications such as acute chest syndrome and painful vaso-occlusive crises [21, 22, 23, 24, 25, 26, 27]. There is a substantial interethnic diversity in the distribution of ENOS variants [22], and this difference could potentially clarify interethnic differences in nitric oxide bioavailability and potentially sickle cell pathophysiology [23]. Based on the diversity of these published reports on ENOS in relations with SCD in different climes, this work was therefore designed to investigate same relationship in south-Eastern Nigeria to ascertain whether or not ENOS polymorphism contributes to SCD pathophysiology in our environment.
Materials and Methods
Study Design
This study was a case control study carried out at the University of Nigeria Teaching Hospital (UNTH) Ituku- Ozalla, Nigeria from February 2020 to December 2020. The study recruited a total of 50 subjects which included 20 SCD patients (SS), 15 healthy sickle cell carriers (AS) and 15 healthy non-carriers (AA), with the last two groups serving as control groups.
Ethical Approval
The study was conducted in accordance with the Helsinki declaration, 2013 version. Ethical clearance certificate with approval number NHREC/05/01/2008B-FWA00002458- 1RB00002323 and reference number UNTH/CSA/329/ VOL.5 was issued by the UNTH Health Research Ethics Committee, dated 12/02/2020. Written informed consents were collected from patients willing to participate in the study. The privacy of patients was kept confidential and patients were free to withdraw from the study at any point in time.
Inclusion/Exclusion Criteria
SCD patients already diagnosed and being followed up at the UNTH Sickle Cell Clinic, who visited clinics on routine check-ups, were recruited for the study and their blood collected as test samples. Patients with severe malaria or critical health complications, and those who were admitted at the hospital were excluded from the study. Also, pregnant and breast-feeding women as well as HIV positive patients and children were excluded from the study. Blood samples for the controls were randomly collected from the UNTH Blood Bank where healthy donors’ blood samples were stored, and sent to Rite Care Hospital (Enugu, Nigeria) where their haemoglobin types were determined using Hemo Type SC screening kit.
Sample Collection
A total volume of 3ml of blood was collected from each patient and transferred into anticoagulant tube (EDTA), and stored at -80oC according to reported methods of storage [24].
DNA Extraction
Human genomic DNA was extracted from blood samples using the Zymo research DNA extraction kit according to the manufacturer’s recommendations. This was done at the Biotech Lab of Godfrey Okoye University, Emene and Enugu, Nigeria. Proteinase K solution was prepared by adding 1040μl to 20mg tube of proteinase K, and stored at -20oC. Two hundred (200) μl Biofluid and Cell Buffer, and 20μl proteinase K were added to 200μl of each of the blood samples in a microcentrifuge tube. The mixture was thoroughly vortexed for about 10minutes and then incubated at 55oC for 10minutes in a water bath. One volume (i.e. 420μl) of Genomic Binding Buffer was added to the mixture, and mixed thoroughly. The whole mixture was transferred to a Zymo-SpinTM IIC-XLR Column in a Collection Tube, centrifuged at about 12000xg for 1minute. The collection tube was then discarded with the flow content. Four hundred (400) μl of DNA Pre-Wash Buffer was added to the column in a new collection tube and centrifuged for another 1minute. The collection tube was emptied of the flow content. Seven hundred (700) μl of g-DNA Wash Buffer was added and centrifuged for 1minute. The collection tube was again emptied of its flow content. Two hundred (200) μl of g-DNA Wash Buffer was added for the final washing, and centrifuged for another 1minute. The collection tube was this time discarded with its flow content. The DNA was then eluted by adding 50μl of DNA Elution Buffer to the column in clean microcentrifuge tube, incubated at room temperature, and then centrifuged for 1minute. The column was finally discarded, and the DNA-containing microcentrifuges for all the samples were stored at -20oC.
PCR-RFLP Genotyping
The SNP (T-786C) in the 5’ promoter region of the ENOS gene was determined by Polymerase Chain Reaction analysis using forward primer 5’GAGTCTGGCCAACACAAATCC3’ and reverse primer 5’ACCTCTAGGGTCATGCAGGT3’ [25]. The PCR reaction mixture was prepared by adding 5μl genomic DNA to a total volume of 12.5μl ready master mix, 1μL of 10pmol/μl of each primer and 5.5μL of water to make up the volume to 25μl. The PCR mixtures were loaded on PCR machine. PCR amplification protocol was: Thirty cycles, consisting of 20seconds denaturation at 94°C, 10 seconds annealing at 58.5°C, and 40 seconds extension at 72°C and the final extension included a 3.5minutes extension at 72°C. The PCR fragment (658bp) was digested with 1 unit of Hpa II restriction enzyme at 37°C maintained on a water bath for 16 hours. The digested and undigested PCR products were estimated with 10μl 100bp DNA ladder in 2% agarose gels. According to Webcutter 2.0 program (rna.lundberg.gu.se/ cutter2) homozygous wild allele (TT) produced 2 fragments 374bp and 284bp; heterozygous mutant allele (TC) produced 3 fragments 374bp, 328bp, and 284bp; whereas homozygous mutant allele (CC) produced 2 PCR fragments 328bp and 284bp.
Data Analysis
Data was analyzed using Hardy-Weinberg equilibrium equation, p2 + 2pq + q2 where p and q are the two traits or polymorphisms being studied. Other statistical tools could not be used because the result produced only TT genotype.
Results
After human genomic DNA was extracted from 20 SS, 15 AA and 13 AS blood samples, the 658bp DNA segment of the gene for the samples as shown by 2% agarose gel are presented in the plates below:
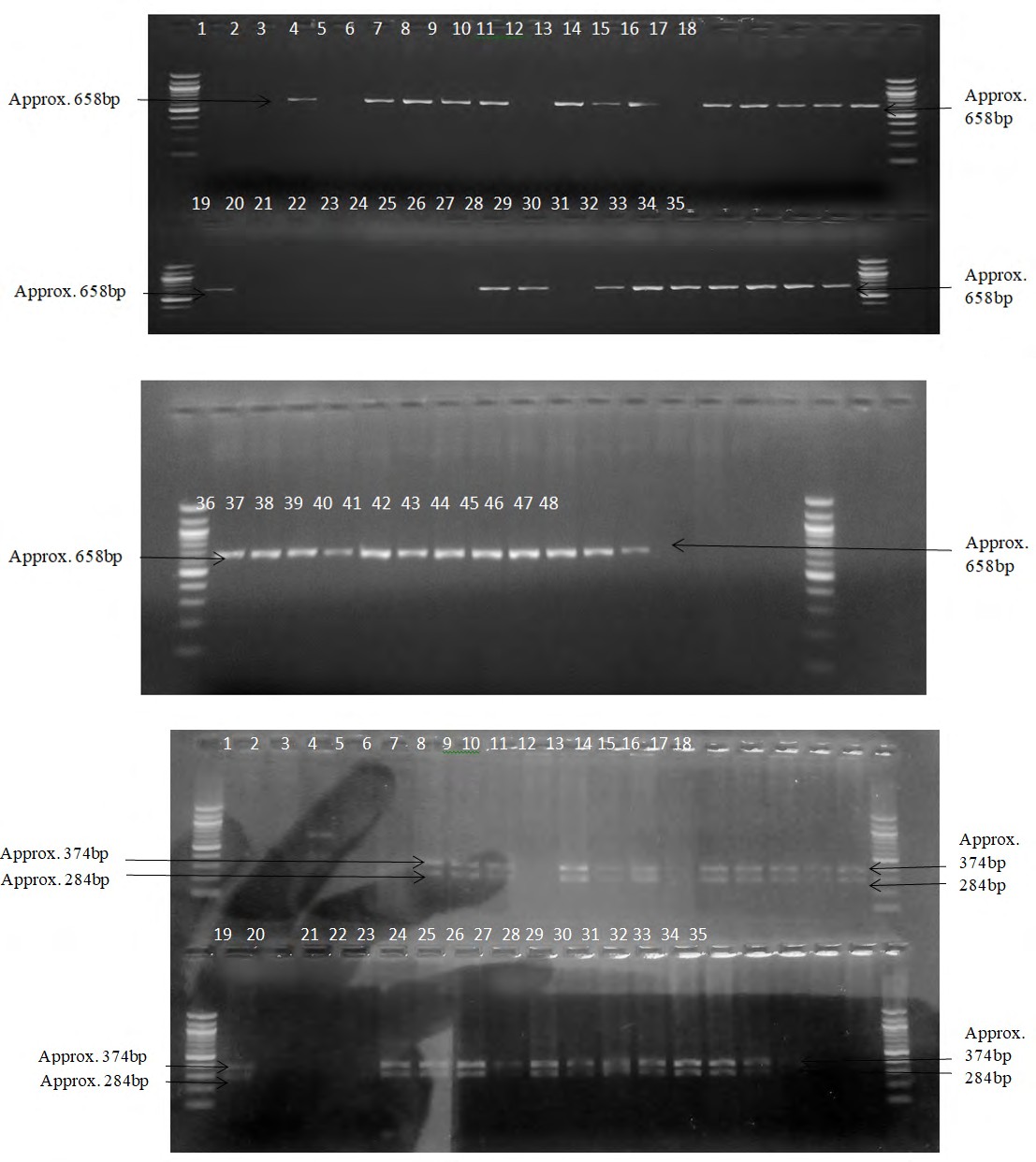
Plate 1: Amplified DNA region corresponding to 658bp band region on the gel. 1-20 represent those of SS DNA; 21-35 represent those of AA DNA.
Plate 2: Amplified DNA region corresponding to 658bp band region on the gel. 36-48 represent those of AS DNA.
The results for the restriction enzyme digestion are presented below in the plates below:
Plate 3: Amplified DNA digestion fragment by HpaII based on the presence/absence of the restriction sites. 1-20 represent those of SS DNA; 21-35 represent those of AA DNA. All show TT genotype.
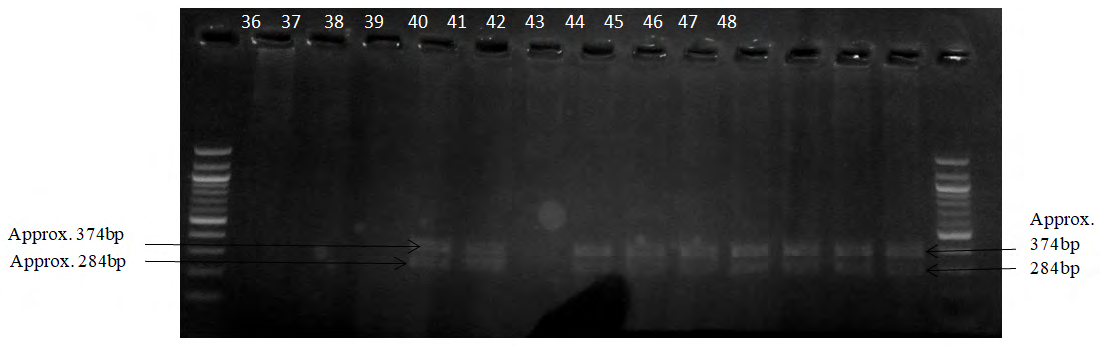
Plate 4: Amplified DNA digestion fragment by HpaII based on the presence/absence of the restriction sites. 36-48 represent those of AS DNA. All show TT genotype.
| Sample Group | TT (284bp/374bp) | TC (374bp/284bp/328bp) | CC (328bp/284bp) | Void |
|---|---|---|---|---|
| SS (n = 20) | 14 (1.0) | 0 | 0 | 6 |
| AA (n = 15) | 12 (1.0) | 0 | 0 | 3 |
| AS (n = 13) | 9 (1.0) | 0 | 0 | 4 |
Table 1: Genotype frequency of ENOS T-786C SNP.
Table 1 shows all the samples are TT genotype irrespective of the sample group. This can be deduced from the appropriate plates above. The presence of two bands around 284bp and 374bp regions as measured from the molecular ladder shows the presence of the TT homozygous genotype. Whereas three bands around 374bp, 284bp and 328bp regions show TC heterozygous genotype; and two bands around 328bp and 284bp imply the CC genotype.
Discussion
The SNP T-786C polymorphism in the 5’ promoter region of human ENOS genes has been implicated in the various disease states ranging from coronary heart and artery diseases [26, 27, 28, 29], sickle cell disease in other studies in different climes [25, 27] and deep vein thrombosis [28], hence the justification for these studies in our own environment. This work evaluated the significance of the T-786C polymorphism in the 5’ promoter region of human ENOS genes in sickle cell subjects using carriers and normal AA individuals as controls. This was aimed at finding the relevance of the single nucleotide polymorphism in ENOS gene in SCD. ENOS is a very important enzyme involved in inflammation, cell–cell interaction and modulation of oxidant injury. _Hpa_II is a type II restriction enzyme which has restriction site at 5’…C|CGG…3’ in both directions at position 786 of the gene 5’ promoter region. An earlier work in the Indian population [27] showed that there is an association between the SNP T-786C ENOS with SCD where they may be acting as genetic modifiers. However, the present work could not establish any relationship between the SNP in the T-786C ENOS gene region and SCD. There was presence of the same genotype in both the test and control groups, in agreement with the reported work in Sudan [25].
Conclusion
This study suggests that T-786C ENOS gene polymorphism is not associated with SCD in our environment. So, T-786C ENOS gene polymorphism cannot genetically influence the phenotypic outcome of the disease in the study population. However, there may be ethnic and geographical differences probably due to evolutionary diversity. Further investigations with larger sample size are needed to confirm or refute the finding in this study.
Recommendation
Larger sample size should be employed for further investigation of the association between this gene and SCD. It should also be investigated in connection with other oxidative free radical enzyme genes such as peroxidases, superoxide dismutase, catalases, glutathione reductase, glutathione peroxidases and glutathione S-transferases. This is because of their involvement in the antioxidant defense system of the body against highly reactive oxidative radicals being generated routinely in the healthy human body and much more in SCD patients. Inter- and intra-ethnic and geographical evolutionary factors should also be considered in further studies.
Declaration of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgment
We are grateful for the support, cooperation, and trust of the patients who released their blood samples for this research. We are also grateful to the medical doctors in the Haematology Clinic and other personnel of UNTH Ituku- Ozalla for their support and cooperation. We also appreciate the assistance of the staff of RiteCare Hospital, Enugu, Nigeria. We also appreciate the cooperation of all the staff of Biotech Lab, Godfrey Okoye University, Emene, Enugu, Nigeria.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
References
-
Konotey-Ahulu FID (1973) Effect of Environment on Sickle Cell Disease in West Africa; Epidemiological and Clinical Considerations. In: Abramson H, et al. (Eds.), Sickle cell disease, diagnosis, management, education and research. St. Louis: The CV. Mosby Company, USA 1973: 20-38.
-
Desai DV, Hiren D (2004) Sickle Cell Disease. History and Origin. The Internet Journal of Hematology 2: 47.
-
Stuart MJ, Nagel RL (2004) Sickle-cell disease. Lancet 364(9442): 1343-1360.
-
Lorenzi TF (2006) Manual de hematologia propedêutica e clínica. 4th (Edn.), Rio de Janeiro: Guanabara Koogan.
-
Jones P (2008) Sickle Cell Disease (Gene & Disease). 1st (Edn.), Chelsea House Publishers, New York.
-
Chirico EN, Pialoux V (2012) Role of oxidative stress in the pathogenesis of sickle cell disease. IUBMB Life 64(1): 72-80.
-
Kaur M, Dangi CBS, Singh M (2013) An overview on sickle cell disease profile. Asian Journal of Pharmaceutical and Clinical Research 6(1): 25-37.
-
WHO (2011) Sickle-cell disease and other haemoglobin disorders. Media Centre. WHO Technical Report Series Fact sheet. World Health Organization, pp: 308.
-
Modell B, Darlison M (2008) Global Epidemiology of haemoglobin Disorders and Derived Service Indicators. Bull World Health Organ 86(6): 480-487.
-
WHO (2013) Sickle cell disease prevention and control. Regional office for Africa, World Health Organization.
-
Serjeant GR, Serjeant BE (2001) The epidemiology of sickle cell disorder: a challenge for Africa. Archives of Ibadan Medicine 2(2): 46-52.
-
Okwi AL, Byarugaba W, Ndugwa CM, Parkes A, Ocaido M, et al. (2010) An up-date on the prevalence of sickle cell trait in Eastern and Western Uganda.BMC Blood Disord 10: 5.
-
Uzoegwu PN, Onwurah AE (2003) Prevalence of haemoglobinopathy and malaria diseases in the population of old Aguata Division, Anambra State, Nigeria. Biokemistri 15(2): 57-66.
-
Driss A, Asare KO, Hibbert JM, Gee BE, Adamkiewicz TV, et al. (2009) Sickle Cell Disease in the Post Genomic Era: A Monogenic Disease with a Polygenic Phenotype. Genomics Insight 2: 23-48.
-
Yetik-Anacak G, Catravas JD (2006) Nitric oxide and the endothelium: history and impact on cardiovascular disease. Vascul Pharmacol 45(5): 268-276.
-
Pritchard KA, Ou J, Ou Z, Shi Y, Franciosi JP, et al. (2004) Hypoxia induced acute lung injury in murine models of sickle cell disease. Am J Physiol Lung Cell Mol Physiol 286(4): L705-L714.
-
Rafikov R, Fonseca FV, Kumar S, Pardo D, Darragh S, et al. (2011) eNOS activation and NO function: Structural motifs responsible for the posttranslational control of endothelial nitric oxide synthase activity. J Endocrinol 210(3): 271-284.
-
Dillon GA, Vita JA (2000) Nitric oxide and endothelial dysfunction. In: Loscalzo J, et al. (Edn.), Nitric Oxide and the Cardiovascular System. Human Press Inc, Totowa, pp: 207-226.
-
Rossi GP, Taddei S, Virdis A, Cavallin M, Ghiadoni L, et al. (2003) The T-786C and Glu 298 Asp polymorphisms of the endothelial nitric oxide gene affect the forearm blood flow responses of Caucasian hypertensive patients. J Am Coll Cardiol 41(6): 938-945.
-
Mack AK, Kato GJ (2006) Sickle cell disease and nitric oxide: A paradigm shift? Int J Biochem Cell Biol 38(8): 1237-1243.
-
Hingorani AD (2001) Polymorphisms in endothelial nitric oxide synthase and atherogenesis: John French Lecture 2000. Atherosclerosis 154(3): 521-527.
-
Serrano NC, Diaz LA, Casas JP, Hingorani AD, Moreno-De- Luca D, et al. (2010) Frequency of eNOS polymorphisms in the Colombian general population. BMC Genet 11: 54.
-
Thakur TJ, Guindo A, Cullifer LR, Li Y, Imumorin IG, et al. (2014) Endothelin-1 but not Endothelial Nitric Oxide Synthase Gene Polymorphism is Associated with Sickle Cell Disease in Africa. Gene Regul Syst Bio 8: 119-126.
-
Bulla A, De Witt B, Ammerlaan W, Betsou F, Lescuyer P (2016) Blood DNA yield but not integrity or methylation is impacted after long-term storage. Biopreserv Biobank 14(1): 29-38.
-
Eltayeb NH, Salih MAM, Muddathir ARM (2015) Endothelial Nitric Oxide Synthase Gene Polymorphism (T-786 C) in Sudanese Patients with Sickle Cell Anaemia. American Journal of Medicine and Medical Sciences 5(5): 231-234.
-
Cattaruzza M, Guzik TJ, Słodowski W, Pelvan A, Becker J, et al. (2004) Shear Stress Insensitivity of Endothelial Nitric Oxide Synthase Expression as a Genetic Risk Factor for Coronary Heart Disease. Cir Res 95(8): 841-847.
-
Nishank SS, Singh MPSS, Yadav R, Gupta RB, Gadge VS, et al. (2013) Endothelial nitric oxide synthase gene polymorphism is associated with sickle cell disease patients in India. J Hum Genet 58(12): 775-779.
-
Akhter MS, Biswas A, Ranjan R, Sharma A, Kumar S, et al. (2010) The nitric oxide synthase 3 gene polymorphisms and their association with deep vein thrombosis in Asian Indian patients. Clin Chem Acta 411(9-10): 649-652.
-
Han Y, Xu W, Zhang W, Liu N, Ji Y (2010) T-786C Polymorphism in the Endothelial Nitric Oxide Synthase Gene is Associated with Increased Risk of Coronary Artery Disease in a Chinese Population. Pharmacology 85(4): 211-216.
- How to Identify and Overcome Barriers in Developing Blood Systems?
- Why Was Transfusion Medicine Not Recognized as a Clinical Discipline?
- Outcomes of Lenalidomide Relapsed/Refractory Patients
- Is Transfusion Always Necessary?
- The Logistics of Production and Use of Blood and Blood Components
- The Challenge for Component Therapies