The Effects of Ganoderma lucidum Extract on Hematological Malignancy Cell Lines through Different Mechanisms
Hematological malignancies are one of the main reasons of death all over the world. Despite the introduction of various treatments for hematopoietic malignancies, they have a variety of side effects that eventually lead to disease recurrence. Resistance to treatment is the main challenge for patient recovery; therefore, to minimize the toxicity of the commerciallyavailable drugs, some alternatives are needed. Ganoderma lucidum is one of the most well-known medicinal fungus species recommended by Asian physicians and naturopaths to prevent and treat various diseases, including cancers, due to its bioactive and pharmacological components. The present review aimed at collecting recent findings on the molecular mechanism of action of this fungus in different hematological cancer cell lines and investigating how it exerts its anti-cancer activity in these cells.
Introduction
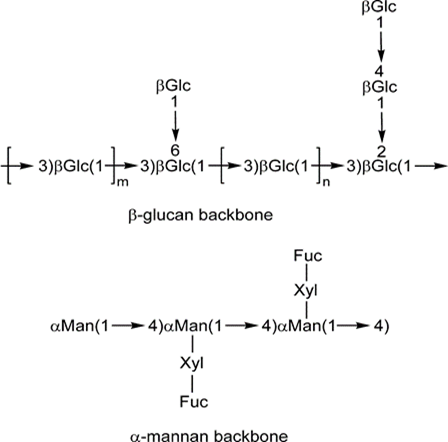
In recent years, people paid considerable attention to natural products, one of their most widely consumed as a folk remedy is Ganoderma lucidum (GL). GL, Reishi or Lingzhi is a basidiomycete, white-rot fungus belonging to the family Ganodermataceae from order Aphyllophorales [1]. It is used in Eastern Asian countries, particularly in traditional Chinese medicine (TCM), for many centuries to prevent and cure diverse human ailments [2], such as nephritis, allergy, bronchitis, asthma, arthritis, hepatitis, hyperglycemia, hypertension, gastric ulcer, hepatopathy, insomnia, and numerous cancers [3]; in addition, it is reported that GL has antioxidant, antiaging, antibacterial, and antiviral activities [4]. Furthermore, G. lucidum is commonly used as an herbal medicine rather than food [5]. Ganoderma lucidum is composed of more than 400 chemical components, including polysaccharides, triterpenoids, amino acids, phenols, sterols, and nucleosides, isolated from the mycelia, spores, and fruiting bodies [6]. Studies indicated that only polysaccharides and triterpenoids, presenting in the natural structure of the fungus, are the most prominently bioactive ingredients and its anti-tumor activity is related to such parts [7].
Polysaccharides, mainly derived from the fruiting bodies of G. lucidum, exert their anti-cancer activity indirectly by increasing host immune responses (Figure 1). Wang Sheng- Yuan, et al. observed that polysaccharides can stimulate the production of cytotoxic T- (CTL) and B-lymphocytes, natural killer (NK) cells, and macrophages, and ultimately increase the secretion of cytokines, such as interleukin (IL)-1B, IL- 6, tumor necrosis factor (TNF), and interferon-ϒ (IFN-ϒ) [8]. Moreover, evidence from previous studies suggests that G. lucidum polysaccharides (GLPs) exert their anticancer activities through immunomodulatory, anti-proliferative, pro-apoptotic, anti-metastatic, and anti-angiogenic effects [9, 10].
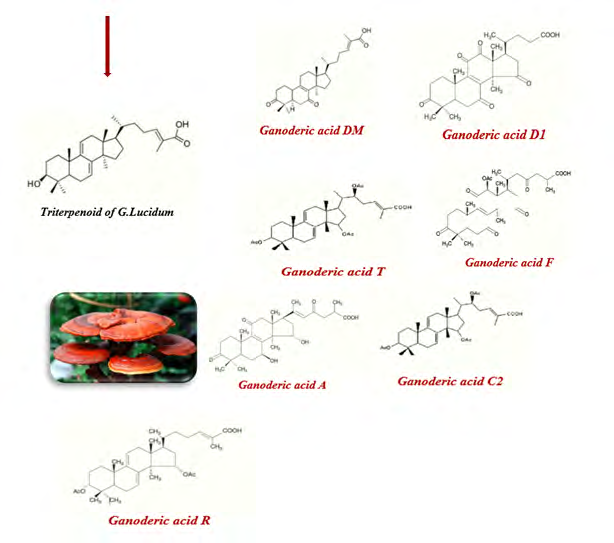
G. lucidum triterpenoids (GLTs), a subtype of trepan, consist of six isoprene units that may form linear or folded chains to form a ring-like structure. Triterpenes, mostly extracted from the spores of G. lucidum, play a pivotal role in directly inhibiting cancer cells. There are different types of triterpenes with various cytotoxic effects (Figure 2). Evidence suggests that some types of triterpenes have potent toxic effects at low concentrations on different human cancerous cell lines [11, 12, 13]. Ganoderic acid T is the most profuse known triterpene of G. lucidum with extraordinary anti-cancer activity in both in-vitro and in-vivo. In a study by Tang Wen, Ganoderic acid T from G. lucidum mycelia induced mitochondria-mediated apoptosis in lung cancer cells [14].
Ganoderic acid an enhanced induction of cisplatin cell death by increasing the sensitivity of HepG2 cells to cisplatin mainly via the signaling transducer and activator of transcription 3 suppression in a study by Yao Xiangyang [15]. Chen Liwu reported that ganoderic acid C2 led to aldose reductase inhibition in the HepG2 cell line [16]. Ganoderic acid D (GAD) is one of the major components in GLTs; in a recent report by Yue et al., it was observed that GAD causes G2/M cell cycle arrest and induces apoptosis in Hela human cervical cancer cells [17]. Xia Junbo showed that ganoderic acid DM induces the autophagic apoptosis in non-small lung cancer cells by suppressing the PI3K/AKT/mTOR activity [18].
Currently available data show that Lingzhi induces apoptosis and suppresses the growth and proliferation of cancerous cells through multiple molecular mechanisms; therefore, it can be concluded that this mushrroom has a potent cytotoxicity against different cancers, including leukemia. The current study aimed at investigating the mechanisms of cancer cell killing by GLPs and GLTs in hematopoietic cell lines.
Effect of Ganoderma lucidum on the THP-1 Cell Line (Acute Monocytic Leukemia)
A study investigating the effect of GLPs extract on acute monocytic leukemia concluded that GLP was capable of inducing death receptor ligands such as TNF-alpha and TRAIL to trigger signaling followed by oligomerization of death receptors, utilization of adaptor proteins, and stimulation of caspase cascade. Therefore, induction of apoptosis via death receptors after treating THP-1 cells with G. lucidum extract plays a pivotal role in the research [19].
Chan Wing Keung, et al. in a similar study, elucidated that GLPs have an immunomodulating role in THP-1 cells. GLPs possess the potential to induce the differentiation of THP-1 into DCs in the presence of cytokines, including IL-4 and GM- CSF; this event is associated with significant upregulation of antigen expression and costimulation molecules, and it should be noted that GLP products possibly sorely limit T-cell lymphocytes through downregulation [P2] of IL-10 production. it is observed that GLPs induce the differentiation of autologous blast cells into dendritic cells and decrease tumor cells burden [20].
Watanabe Kenji, et al. investigated the effect of triterpene-rich extract from G. lucidum AF on THP- 1 monocytic cells. Notably, accumulated data show that G. lucidum AF induces TNF-α production in both lipopolysaccharide-treated and non-treated monocytic THP- 1 cells and has a synergistic effect on THP1 cells treated with lipopolysaccharides (LPSs). Evidence suggested that G. lucidum AF increased phosphorylation of p38 MAPK induced by LPSs, which suppressed TNF-α production and decreased phosphorylation of c-Jun N-terminal kinase (JNK) MAPK, induced by LPSs that enhanced TNF-α production. Accordingly, G. lucidum isolated from triterpenes possibly stimulated immune responses, induced by LPSs via the modulation of p38 and JNK MAPKs activation in the THP-1 cell line [21].
Based on previous data published by the journal of the Evidence-Based Complementary and Alternative Medicine, exposure to GLPs strongly induces macrophage differentiation in THP-1 cell line via activation of caspase and p53. It is known that therapeutic differentiation is an important approach to the treatment of hematological cancers, including leukemia, which helps to eliminate the growth and proliferation of cancerous cells. As confirmed by various molecular experiments, caspase activation and up-regulation of p53 and p21 contribute to the macrophage differentiation process. The current study suggested that the treatment of human leukemia THP-1 cell enhances the expression of p53 and p21 and also the activation of the caspase pathway, which facilitate cellular differentiation and elevate the expression of macrophage differentiation markers in such leukemia cells [22].
Emerging evidence demonstrated that treatment with active lipids isolated from G. lucidum spores can drastically induce apoptosis in the THP-1 cell line through the suppression of ERK1/2 and AKT and activation of JNK1/2. Moreover, treatment of THP-1 cells with active lipids from G. lucidum led to upregulation of caspase 3, 8, and 9 in a dose- and time-dependent manner, which mediated apoptotic induction. Hence, according to reports, MAPK and PI3K signaling pathways play the most prominent role in the modulation of cell death by active lipids from G. lucidum in THP-1 monocytic cells [23].
Rathor Richa, et al. studied the impact of hydro-ethanolic extract of G. lucidum (HEGL) on inflammatory cytokines,and NF-kB activity and also its antioxidant property in monocytic THP-1 cells. Results of their study indicated that HEGL strongly suppressed the production of inflammatory mediators, including TNF-α, IFN-y, IL-1β, NO, and NF-kB in LPS-stimulated THP-1 cells; hence, this information proved the immunomodulatory activity of HEGL in LPS-treated THP- 1 cells [24].
The Effect of Ganoderma lucidum on Ramos and Daudi Cell Lines (Burkitt’s Lymphoma)
Recently, to determine the magnitude of apoptotic cell death, Ganoderic acid A (GAA) treatment was also examined on Ramos and Daudi cells; the accumulated data in this regard illustrated that the GLT had profound apoptotic characteristics through an intrinsic pathway correlated with mitochondrial dysfunction in such cells. The results of studies indicated that GAA treatment disturbed the mitochondrial membrane potential followed by enhanced cytochrome c cytosolic levels and cleavage caspases 3, 8, and 9. In addition, pro-apoptotic protein BIM and BAX up-regulated by GLT and downregulated anti-apoptotic protein bcl-2; hence, GAA in a dose- and time-dependent manner inhibited the cell proliferation and induced apoptosis [25].
In another study, G. lucidum extract was examined on a panel of 26 human cancer cell lines, including Daudi and Ramos, and reported its antitumor activity. This preliminary screening study displayed that the extract from G. lucidum induced apoptosis in these cell lines, but unfortunately, the underlying mechanism of the anti-cancer activity remained unknown in Daudi and Ramos cell lines (Table 1) [26].
| Type of G.lucidum component | Type of study | Type of cell line | Results | Potential mechanism of action | IC50 | Incuba- tion time | Reference |
|---|---|---|---|---|---|---|---|
| Polysaccharide fraction of G.lucidum | Laboratory | Acute monocytic leukemia (FAB M5) | Induce apoptosis in THP-1 cell line | Induces death receptor ligands such as TNF-a and TRAIL and activation of caspase cascades | 30 μg/ml | 48 h | Cheng Kun- Chieh, et al. [19] |
| Polysaccharide fraction of G.lucidum | Laboratory | Acute monocytic leukemia (FAB M5) | Reduce cancer cell burden in THP-1 cell line | Increase the cell proliferation and induce differentiation of THP-1 into DC and down-regulating of T-lymphocyte through by decrease IL-10 | 100 μg/ ml | 72 h | Chan Wing Keung, et al. [20] |
| Triterpenes rich-extract of G.lucidum AF | Laboratory | Acute monocytic leukemia (FAB M5) | Induce TNFa production in THP-1 cell line | Increase LPS-induced phosphorylation of p38 MAPK and inhibit LPS-induced phosphorylation of JNK MAPK | 50 μg/ml | 4 h | Watanabe Kenji, et al. [21] |
| Polysaccharide fraction of G.lucidum | Laboratory | Acute monocytic leukemia (FAB M5) | Induce macrophage differentiation in THP-1 cell line | activate caspase cascade and up-regulating of p53 and p21 and elevated of Macrophage differentiation markers | 30μg/ml | 4 h | Hsu Jia-Wei, et al. [22] |
| Active lipid of G.lucidum spores | Laboratory | Acute monocytic leukemia (FAB M5) | Induce apoptosis in THP-1 cell line | Inhibit ERK1/2, AKT, activate JNK1/2 signaling pathway and up-regulating of caspase 3, 8 and 9 | 1 mg/ml | 48 h | Wang Jia- He, et al. [23] |
| Hydro- ethanolic extract of G.lucidum (HEGL) | Laboratory | Acute monocytic leukemia (FAB M5) | Possesses anti-oxidant property in LPS-treated THP-1 cells | Inhibit production of TNF-α, IFN-y, IL-1β, NO and NF-kB | 100 μg/ml | 24 h | Rathor Richa, et al. [24] |
| Triterpenoid fraction of G.lucidum | Laboratory | Burkitt’s lymphoma (Ramos. Daudi) | Induce apoptosis in Ramos and Daudi cell lines | Enhance activation of caspase 3, 8 and 9, up-regulating of pro-apoptotic proteins BAX. BIM and down-regulating of anti-apoptotic proteins bcl-2 | 18.5-22 μM | 24 h | Radwan Faial FY, et al. [25] |
| Triterpenoid fraction of G.lucidum | Laboratory | Human acute lymphoblastic leukemia (FAM M2) | Induce apoptosis in HL-60 cell line | Up-regulating of bcl- 2, BAX translocation, mitochondrial cytochrome c release and caspase 3 activation | 210 μg/ ml | 6,24 h | Kim Kug Chan, et al. [27] |
| Triterpenoid fraction of G.lucidum | Laboratory | Human acute lymphoblastic leukemia (FAM M2) | Induce apoptosis in HL-60 cell line | Inhibit CDK1 phosphorylation and the dephosphyraltion of pRB and induce G1 phase arrest | 150 μg/ ml | 48h | Hsu Jia-Wei, et al. [22] |
| Triterpenoid fraction of G.lucidum | Laboratory | Human acute lymphoblastic leukemia (FAM M2) | Induce apoptosis in HL-60 cell line | Inhibit CDK1 phosphorylation and the dephosphyraltion of pRB and induce G1 phase arrest | 150 μg/ ml | 48h | Kim Kug Chan, et al. [28] |
| G.lucidum extract | Laboratory | Human acute lymphoblastic leukemia (FAM M2) | Reduce the proliferation activity in HL- 60 cell line | Decrease mitochondrial membrane potential and decrease G1 phase progression | 136.3 μg/ml | 48h | Liu Yue Wei, et al. [29] |
| ergosta‐7,22‐ diene‐2β,3α,9α‐ triol (EGDT) | Laboratory | Human acute lymphoblastic leukemia (FAM M2) | Induce apoptosis in HL-60 cell line | Increase DNA fragmentation and activation of caspase 3 | 12.7 μg/ ml | 24h | Lee Mi Kyoung, et al. [30] |
| Polysaccharide fraction of G.lucidum | Laboratory | Human acute lymphoblastic leukemia (FAM M2) | Induce apoptosis in HL-60 cell line | Activate p38 and JNK MAPK pathway and regulating of their downstream genes and proteins | 200-800 μg/ml | 20,48,72h | Yang Gouhua, et al. [31] |
| Polysaccharide fraction of G.lucidum | Laboratory | Human chronic myelocytic leukemia | Reduce resistance to Adriamycin in K562 cell line | Regulating of MDR-1 and MRP-1 transcription factors | 50 mg/L | 44h | Li Wei dong, et al. [32] |
| Ganoderma extract and spores oil | Laboratory | Human chronic myelocytic leukemia | Blocking the cell cycle in K562 cell line | inhibit the topoisomerase I and II activity and decrease of cyclin D1 | 0.39 mg/ ml | 24h | Chen Chun, et al. [33] |
| Fruiting bodies of G.lucidum | Laboratory | Human acute promyelocytic leukemia (FAB M3) | Induce apoptosis in NB4 cell line | Reduce of p53, AKT and ERK levels | 60 μg/ml | 19h | Calvino Eva, et al. [34] |
| Ergosterol peroxide (EP) | Laboratory | Human multiple myeloma | anti-tumor activity in U266 cell line | Inhibit STAT3 signaling pathway | 25 μM | 24h | Rhee Yun Hee, et al. [35] |
| Fruiting bodies of G.lucidum | Laboratory | Pre-B acute lymphoblastic leukemia | Causes apoptosis in Blin1 cell line | No cell cycle arrest was observed in G2/M phase | 38 μg/ml | 96h | Muller Claudia I, et al. [26] |
| Fruiting bodies of G.lucidum | Laboratory | Non-T, non-B acute lymphoblastic leukemia | Causes apoptosis in NALM-6 cell line | Arrest the cell cycle in G2/M phase | 30 μg/ml | 96h | Muller Claudia I, et al. [26] |
| Triterpenoid fraction of G.lucidum | Laboratory | Non-T, non-B acute lymphoblastic leukemia | Induce apoptosis in NALM-6 cell line | Regulating of caspase activity and BAX , BCL-2 and APAF1 expression | 40 mM | 24h | Radwan Faisal FY, et al. [25] |
| Triterpenoid fraction of G.lucidum | Laboratory | Non-T, non-B acute lymphoblastic leukemia | Induce apoptosis in NALM-6 cell line | Reduce the expression of miR- 17-5p and miR- 181b and induce apoptosis | 140 μg/ ml | 48h | Mortazavie Faezeh, et al. [36] |
| Diffuse histiocytic leukemia ( monocytic leukemia cell) | Laboratory | Causes apoptosis in U937 cell line | Increase protein expression of p21WAFI and p27 KIPI and No cell cycle arrest was observed in G2/M phase | 63 μg/ml | 96 h | Muller Claudia I, et al. [26] | |
| Fruiting bodies of G.lucidum | Laboratory | Multiple myeloma | Causes apoptosis in RPMI8226 cell line | Arrest the cell cycle in G2/M phase | 40 μg/ml | 96 h | Muller Claudia I, et al. [26] |
Table 1: Selected laboratory and pre-clinical studies of G.lucidum component.
Effect of Ganoderma lucidum on the HL-60 Cell Line (Acute Myeloblastic Leukemia)
Kug Chieh Chan Kim, et al. investigated the effect of G. lucidum extract on human promyelocytic leukemia cells (HL-60) and found that this fungus alone or in combination with Duchesnea chrysatha (GDE) increased the permeability of the mitochondrial membrane and induced the apoptosis in the HL60 cell line. Based on these data, the reduction in mitochondrial membrane potential was probably related to cytochrome c release, Bax expression, caspase3 activation, and BCL-2 regulation [27].
A study published in 2008 in Cancer Letters journal indicated that the simultaneous use of the combination of GL and DL with radiotherapy strongly induced mitochondrial damage and apoptosis. This combination triggered apoptosis through a mitochondrial-intrinsic pathway by increasing the activity of proapoptotic proteins, such as Bax, Smac/DIABLO, and caspase-3. Also, after exposure to Gl and DC, the HL-60 cells became more sensitive to γ-irradiation by changing the level and duration of the cell cycle arrest in G1 and G2/M phases through inhibition of pRB and CDK1/cyclin B1 checkpoint proteins phosphorylation, respectively [28].
It is recently found that ethanolic extracts of G. lucidum have anti-proliferative activity on acute promyelocytic leukemia cells. After treatment with Ganoderma extract, the HL-60 cells exhibit time-dependent changes in the cell cycle. The cell cycle was significantly arrested at the G2/ M transition [29].
It is reported that, among the chemical compounds in the fruit body of G. lucidum, EGDT (ergosta‐7, 22‐diene‐2β, 3α, 9α‐triol) has the highest cytotoxic effect on HL-60 cells. EGDT enhanced cleavage of procaspase-3, procaspase‐3, poly (ADP‐ ribose) polymerase (PARP), and DNA fragmentation that are associated with apoptosis process; though the apoptotic activity of EGDT was a dose‐dependent manner [30].
An investigation on the polysaccharide obtained from G. lucidum indicated its anti-cancer effects, both directly through signaling pathways and indirectly by regulating the immune system. These findings illustrated that phosphorylated MEK and ERK1/2 in the cells treated with GLP remarkably reduced in a dose- and time-dependent manner. GLP blocked the ERK/MAPK signaling pathway. On the other hand, the levels of IL-2, IL-6, IL-12, and TNF-a in the serum of the nude mice xenograft model were measured, and the results indicated that GLP increased their levels; thus, GLP can indirectly regulate the immune system [31].
The Effect of Ganoderma lucidum on The K562 Cell Line (Erythroid Chronic Myeloid Leukemia)
One of the reasons for cancer treatment failure is multidrug resistance (MDR). Overexpression of MDR1 (multidrug resistance), MRP1 (MDR-associated protein), and P-gp (P-glycoprotein) is observed in drug-resistance cell lines. In a previous study, in order to find a drug with higher MDR activity [P3] and lower toxicity, the effect of polysaccharides (Gl-PS) extracted from the fruit body of G. lucidum on the multidrug cell line K562/ADM was investigated. The findings showed that the expression of MRP1 and MDR1 decreased in exposure to GL-PS. Although Gl-PS had less toxic effects on normal cells, it reversed the resistance of K562/ADM to ADM [32].
A study on the effects of G. lucidum extract and spore oil on the molecular mechanisms, underlying their effects on the K562 cell line, specified that its anti-growth activity is due to blocking cell cycle at the transition between the G1 and S phases, decrease in cyclin D1 levels, and topoisomerase suppression. Topoisomerase enzyme is involved in the regulation of DNA supercoiling. Also, topoisomerase overexpression is observed in some hematologic malignancies. The suppression of topoisomerase I and II activities destructs dividing cancer cells in a dose dependent manner [33].
Effect of Ganoderma lucidum on NB4 Cell Line (Promyelocytic Leukemia)
Chen Chun, et al. discovered that NB4 human leukemia cells underwent intracellular changes after treatment with G. lucidum aqueous extract. Decrease in cell viability by inducing apoptosis, reduction of p53, Bcl-2 level, Erk, and pErk2 proteins synthesis, and increase in the Bax level were all due to the effect of Ganoderma on NB4 [34].
Effect of Ganoderma lucidum on U266 Cell Line (Multiple Myeloma)
One of the components of G. lucidum is ergosterol peroxide (EP), which has an anti-tumor effect on the U266 multiple myeloma cells. According to studies, the antitumor mechanism of EP on U266 is associated with its inhibitory effects on the signaling pathways of stat3. The stat3 is a cytoplasmic transcription factor involved in the regulation of genes encoding apoptosis inhibitors, such as BCL-xL, and BCL-2. EP remarkably suppresses JAK2 and Src activation, stat3 phosphorylation, and stat3 DNA-binding activity. It also induces SHP-1 protein expression, playing a pivotal role in inhibiting stat3 phosphorylation; all the processes were in a time- and dose-dependent manner [35].
Effect of Ganoderma lucidum on the Nalm-6 Cell Line (Non-T, Non-B Acute Lymphoblastic Leukemia)
Muller assessed the cytotoxic potential of G. lucidum extract against human cancer cell lines and reported that G. lucidum effectively inhibited the proliferation of Nalm-6 cells, which were one of the cell lines with the greatest sensitivity to this fungus. As explained, the fungal extract has an antitumor proliferation activity via both apoptotic pathway and cell cycle arrest on the G2/M phase, but unfortunately, information in the current study does not elucidate the exact mechanism by which the GLE induces apoptosis [26].
A study on Nalm-6 cells proved that Ganodric acid A reduces cell viability. Also, according to the findings, increased caspase expression and activity 3, 8, 9, and pro-apoptotic BIM, BAX proteins, decreased BCL-2 protein, high levels of APAF-1, and cytochrome c consequently led to apoptotic cell death in pre-B acute lymphocytic leukemia [25].
In an article published in 2022, the anticancer activity of Ganoderic acid an extract was evaluated on changes in the expression of miR-17-5p and miR-181b and the induction of apoptosis in the Nalam 6 cell line. According to the results of this study, Ganoderic acid A has the ability to greatly reduce the expression of miR-17-5p and miR-81b and induce cell apoptosis in the Nalm-6 leukemic cell line [36].
The effect of Ganoderma lucidum on the Blin-1 (pre-B acute lymphoblastic leukemia), Jurkat (T-cell acute lymphoblastic leukemia), U937 (diffuse histocytic leukemia), SUDHL6 (diffuse large B-cell lymphoma), and the ARH77 and RPMI8226 cell lines (multiple myeloma). A paucity of laboratory and pre-clinical studies evaluated anti-tumor activity of Reishi mushroom on the human cell lines, such as Blin-1, Jurkat, U937, AHR77, RPMI8226, and SUDHL6; the findings of their field suggested that G. lucidum extract induced apoptosis in these cell lines, but the results were incomplete from certain aspects; hence, further research is needed until exact mechanism of action is fully elucidated. Based on the available and relevant information, after the exposure of U937 cell line to different concentrations of G. lucidum extract for 48 and 72hours, the expression of apoptosis-related proteins, including p21 and p27, increased; even though there was no G2/M arrest after treatment with G. lucidum in the U937 cells in; additionally, it was observed that G. lucidum extract lightly enhanced cells in the RPML8226 in the G2/M phase [26].
Conclusion
Ganoderma lucidum is one of the famous medicinal fungi regarded as a promising anti-cancer immunotherapy agent due to its pharmacological properties and minimum side effects; since G. lucidum could be a drug of choice for the development of a novel class of anti-cancer drug. The present review evaluated the recent findings on the exact mechanism pathway in hematological cancer cell lines. Until now, extensive studies are performed on the anti-cancer activity of this magic mushroom; nevertheless, molecular and immune-supportive mechanisms in cancer cell lines are not fully elucidated accordingly. There is a scope for prospective research, especially since the mechanisms underlying immune modulation should be explored in detail, which can prove the efficiency and safety of this fungus; perhaps further studies provide new insight into the potential therapeutic applications of G. lucidum to combat cancer.
Acknowledgments
Authors extend their gratitude to the staff of the Diagnostic Laboratory Sciences and Technology Research Center of the School of Paramedical Sciences, Shiraz University of Medical Sciences, and Shiraz, Iran.
Conflict of Interest Statement
The authors declared no conflicts of interest. It was a review study and informed consent or ethical approval was not applicable.
Funding
There was no funding for the study. Table selected laboratory and pre-clinical studies of G. lucidum component.
References
-
Khoo SC, Ma NL, Peng WX, Ng KK, Goh MS, et al. (2022) Valorisation of biomass and diaper waste into a sustainable production of the medical mushroom Lingzhi Ganoderma lucidum. Chemosphere 286(1): 131477.
-
Luo H, Tan D, Peng B, Zhang S, Vong CT, et al. (2022) The Pharmacological Rationales and Molecular Mechanisms of Ganoderma lucidum Polysaccharides for the Therapeutic Applications of Multiple Diseases. Am J Chin Med 50(1): 53-90.
-
Blundell R, Camilleri E (2022) Ganoderma lucidum–the mushroom of immortality.
-
Cör Andrejč D, Knez Ž, Knez Marevci M (2022) Antioxidant, antibacterial, antitumor, antifungal, antiviral, anti-inflammatory, and nevro-protective activity of Ganoderma lucidum: An overview. Front Pharmacol 13: 934982.
-
El Sheikha AF (2022) Nutritional Profile and Health Benefits of Ganoderma lucidum Lingzhi, Reishi, or Mannentake as Functional Foods: Current Scenario and Future Perspectives. Foods 11(7): 1030.
-
Pathak I, Saxena M, Pandey SJIJOBS. A Review on Ganoderma lucidum: An Important Medicinal Mushroom.
-
Xu J, Shen R, Jiao Z, Chen W, Peng D, et al. (2022) Current Advancements in Antitumor Properties and Mechanisms of Medicinal Components in Edible Mushrooms. Nutrients 14(13): 2622.
-
Gharibzahedi SMT, Marti-Quijal FJ, Barba FJ, Altintas Z (2022) Current emerging trends in antitumor activities of polysaccharides extracted by microwave-and ultrasound-assisted methods. Int J Biol Macromol 202: 494-507.
-
Wang M, Yu F (2022) Research Progress on the Anticancer Activities and Mechanisms of Polysaccharides from Ganoderma. Front Pharmacol 13: 891171.
-
Hu Y, Lin Z, Fu H, Xia J, Xiong W, et al. (2022) Immunomodulatory effect of Ganoderma lucidum polysaccharide extract on peritoneal macrophage function of BALB/c mice. Cell Mol Biol 68(4): 31-34.
-
Kolniak-Ostek J, Oszmiański J, Szyjka A, Moreira H, Barg E, et al. (2022) Anticancer and Antioxidant Activities in Ganoderma lucidum Wild Mushrooms in Poland, as Well as Their Phenolic and Triterpenoid Compounds. Int J Mol Sci 23(16): 9359.
-
Ostek KJ, Oszmiański J, Szyjka A, Moreira H, Barg E (2022) Anticancer and Antioxidant Activities in Ganoderma lucidum Wild Mushrooms in Poland, as Well as Their Phenolic and Triterpenoid Compounds. Int J Mol Sci 23(16): 9359.
-
Cao L, Jin H, Liang Q, Yang H, Li S, et al. (2022) A new anti- tumor cytotoxic triterpene from Ganoderma lucidum. Nat Prod Res 36(16): 4125-4131.
-
Tang W, Liu JW, Zhao WM, Wei DZ, Zhong JJ, et al. (2006) Ganoderic acid T from Ganoderma lucidum mycelia induces mitochondria mediated apoptosis in lung cancer cells. Life Sci 80(3): 205-211.
-
Yao X, Li G, Xu H, Lü C (2012) Inhibition of the JAK- STAT3 signaling pathway by ganoderic acid A enhances chemosensitivity of HepG2 cells to cisplatin. Planta Med 78(16): 1740-1748.
-
Chen L, Du J, Dai Q, Zhang H, Pang W, et al. (2014) Prediction of anti-tumor chemical probes of a traditional Chinese medicine formula by HPLC fingerprinting combined with molecular docking. Eur J Med Chem 83: 294-306.
-
Yue QX, Cao ZW, Guan SH, Liu XH, Tao L, et al. (2008) Proteomics characterization of the cytotoxicity mechanism of ganoderic acid D and computer-automated estimation of the possible drug target network. Mol Cell Proteomics 7(5): 949-961.
-
Xia J, Dai L, Wang L, Zhu J (2020) Ganoderic acid DM induces autophagic apoptosis in non-small cell lung cancer cells by inhibiting the PI3K/Akt/mTOR activity. Chem Biol Interact 316: 108932.
-
Cheng KC, Huang HC, Chen JH, Hsu JW, Cheng HC, et al. (2007) Ganoderma lucidum Polysaccharides In Human Monocytic Leukemia Cells: From Gene Expression To Network Construction. BMC Genomics 8(1): 1-17.
-
Chan Wk, Cheung Cch, Law Hkw, Lau Yl, Chan Gcfjjoh (2008) Ganoderma lucidum Polysaccharides Can Induce Human Monocytic Leukemia Cells into Dendritic Cells with Immuno-Stimulatory Function. J Hematol Oncol 1(1): 1-12.
-
Watanabe K, Shuto T, Sato M, Onuki K, Mizunoe S, et al. (2011) Lucidenic Acids-Rich Extract from Antlered Form of Ganoderma lucidum Enhances Tnfα Induction in Thp- 1 Monocytic Cells Possibly Via Its Modulation of Map Kinases P38 and Jnk. Biochem Biophys Res Commun 408(1): 18-24.
-
Hsu JW, Huang HC, Chen ST, Wong CH, Juan H et al. (2011) Ganoderma lucidum Polysaccharides Induce Macrophage-Like Differentiation in Human Leukemia Thp-1 Cells Via Caspase and P53 Activation. Evid Based Complement Alternat Med 2011: 358717.
-
Wang JH, Zhou YJ, Zhang M, Kan L, Pjjoe H (2012) Active Lipids of Ganoderma lucidum Spores-Induced Apoptosis in Human Leukemia Thp-1 Cells Via Mapk and Pi3k Pathways. J Ethnopharmacol 139(2): 582-589.
-
Rathor R, Tulsawani R, Misra Kjijops, Research (2014) Hydro-Ethanolic Extract of Ganoderma lucidum (Hegl) Shows Anti-Inflammatory Activity on Thp1 Cytokines and Nf-[Kappa] B P65 Response. International Journal of Pharmaceutical Sciences and Research 5(6): 2337-2348.
-
Radwan Ff, Hossain A, God Jm, Leaphart N, Elvington M, et al. (2015) Reduction of Myeloid‐Derived Suppressor Cells and Lymphoma Growth by A Natural Triterpenoid. J Cell Biochem 116(1): 102-114.
-
Müller Ci, Kumagai T, O’kelly J, Seeram Np, Heber D, et al. (2006) Ganoderma lucidum Causes Apoptosis in Leukemia, Lymphoma and Multiple Myeloma Cells. Leuk Res 30(7): 841-848.
-
Kim Kc, Kim Js, Son Jk, Kim Igjcl (2007) Enhanced Induction of Mitochondrial Damage and Apoptosis in Human Leukemia Hl-60 Cells by the Ganoderma lucidum and Duchesnea Chrysantha Extracts. Cancer Lett 246(1- 2): 210-217.
-
Kim Kc, Jun Hj, Kim Js, Kim IG (2008) Enhancement of Radiation Response with Combined Ganoderma lucidum and Duchesnea Chrysantha Extracts in Human Leukemia Hl-60 Cells. Int J Mol Med 21(4): 489-498.
-
Liu YW, Gao JL, Guan J, Qian ZM, Feng K, et al. (2009) Evaluation of Antiproliferative Activities and Action Mechanisms of Extracts From Two Species of Ganoderma on Tumor Cell Lines. J Agric Food Chem 57(8): 3087- 3093.
-
Lee Mk, Hung Tm, Cuong Td, Na M, Kim Jc, et al. (2011) Ergosta‐7, 22‐Diene‐2β, 3α, 9α‐Triol From The Fruit Bodies of Ganoderma lucidum Induces Apoptosis in Human Myelocytic Hl‐60 Cells. Phytother Res 25(11): 1579-1585.
-
Yang G, Yang L, Zhuang Y, Qian X, Shen Yjjor (2016) Ganoderma lucidum Polysaccharide Exerts Anti-Tumor Activity Via Mapk Pathways in Hl-60 Acute Leukemia Cells. J Recept Signal Transduct Res 36(1): 6-13.
-
Li WD, Zhang BD, Wei R, Liu JH, Lin Z (2008) Reversal Effect of Ganoderma lucidum Polysaccharide on Multidrug Resistance in K562/Adm Cell Line. Acta Pharmacol Sin 29(5): 620-627.
-
Chen C, Li P, Li Y, Yao G, Xu Jhjol (2016) Antitumor Effects and Mechanisms of Ganoderma Extracts and Spores Oil. Oncol Lett 12(5): 3571-3578.
-
Calviño E, Manjón Jl, Sancho P, Tejedor Mc, Herráez A, et al. (2010) Ganoderma lucidum Induced Apoptosis in Nb4 Human Leukemia Cells: Involvement of Akt and Erk. J Ethnopharmacol 128(1): 71-78.
-
Rhee YH, Jeong SJ, Lee HJ, Lee HJ, Koh W, et al. (2012) Inhibition of Stat3 Signaling and Induction of Shp1 Mediate Antiangiogenic and Antitumor Activities of Ergosterol Peroxide in U266 Multiple Myeloma Cells. BMC Cancer 12(1): 1-11.
-
Mortazavie F, Taheri S, Tandel P, Zare F, Gjijoph T (2022) The Effect of Ganoderic Acid A on Mir-17-5p and Mir- 181b Expression Level and Apoptosis Induction in Human Leukemia Nalm-6 Cells. Iran J Ped Hematol Oncol 12(3): 152-163.
- How to Identify and Overcome Barriers in Developing Blood Systems?
- Why Was Transfusion Medicine Not Recognized as a Clinical Discipline?
- Outcomes of Lenalidomide Relapsed/Refractory Patients
- Is Transfusion Always Necessary?
- The Logistics of Production and Use of Blood and Blood Components
- The Challenge for Component Therapies