A Comparison between Conventional Methods and Proton- Magnetic Resonance Spectroscopy (H1-MRS) for Estimation of Postmortem Interval in Six Different Species: A Systematic Literature Review
Determination of the postmortem interval in forensic investigations is an important mission to achieve justice in the world of crime. Conventional methods used to date allow the estimation of the time lapse since death depending on circumstantial evidence, livor and rigor mortis evaluation, and core body temperature identification. They are effective methods, but in some cases, they are not feasible enough to achieve accurate estimation, especially in the occurrence of putrefaction, as there are no reliable data on putrefied cadavers. This review was undertaken to show the efficiency of different methods for the determination of the postmortem interval of six-species cadavers, with an analysis focus on brain metabolic variables using proton-magnetic resonance spectroscopy (H1-MRS) in further work. The review considered all published work in PubMed, Google Scholar, and eight other search engines concerning the time lapse since death using conventional methods and H1-MRS. Additionally, some graphs were plotted to help in data analysis and synthesis. It revealed that H1-MRS can determine the postmortem interval in six different-species cadavers either in an early or late period of death and even after the occurrence of putrefaction. To this end, it is concluded that the identification of specific brain metabolites and estimation of their concentration changes by using H1-MRS may become a good way to identify time since death accurately and beyond the currently achievable range.
Introduction
Estimation of postmortem interval (PMI) has been very important in criminal investigations, as knowing the exact time of death helps in the collection of evidence, determines the investigation direction, and shares in the inclusion or exclusion of suspects [1]. The conventional methods usually used for the determination of PMI include the degree of livor mortis, rigor mortis extension, rate of body cooling, and signs of putrefaction. They are used worldwide and trusted by many pathologists and criminal courts until the third day postmortem. After that, they cannot be reliable due to the autolysis of tissues and the bacterial decomposition of the body [2]. Forensic specialists always prefer using these parameters rather than biochemical alterations in their legal investigations. The reliance on biochemical alterations is of concern due to the rapid changes in the chemical constituents of the body following death. When the cell membrane loses its selective permeability, the concentration gradient of metabolites is compromised, and the metabolites are degraded [3]. These alterations occur initially in the blood tissue after a few hours postmortem, then in CSF after 15- 20 hours PM, and in the vitreous humor of the eye after 120 hours PM [3]. Therefore, blood and CSF are unsuitable source materials for the estimation of PMI after only one day PM. In addition, there has been controversy over using vitreous humor as source material, and the issue is being studied to determine its suitability for the determination of PMI [4]. Moreover, the biochemical analysis of various tissues, such as the liver or muscles, is ineffective due to the heterogeneity of organ metabolism caused by individual variations, the effect of disease conditions, or the administration of treatments prior to death [5].
Conversely, the biochemical analysis of the brain tissue would be most suitable for the estimation of PMI, as the brain is neither subject to individual variations nor bacterial putrefaction-based rapid metabolic changes postmortem, protected by the skull from environmental influences, and away from the intestine and bacterial invasion [5]. Additionally, the presence of the blood‒brain barrier helps reduce the metabolic alterations caused by treatments taken prior to death. When scientists recognised these facts about the brain several years ago, they could propose a relative method for the estimation of PMI in a period of more than three days PM. They wanted to extend the range achieved by the conventional methods, overcome their disadvantages, and increase the accuracy of the estimation. The experiment was carried out for 4-20 days PM on the basis of glutamic acid reactions. However, the procedure did not give rise to real practical relevance due to the long-time consumption and the requirement for lab-intensive chemical analysis [6, 7, 8].
In recent years, along with imaging system evolution, proton-magnetic resonance spectroscopy (H1-MRS) has been developed as an ionising radiation-free analytical technique. It was found to provide a non-invasive method for studying metabolic changes in brain affections. This technique has been used to improve tissue characterisation by MRI through the estimation of certain brain metabolite concentrations, in addition to the two-dimensional or three-dimensional image created by MRI [1].
Therefore, researchers have extrapolated that H1-MRS could be a promising tool to estimate PMI accurately and for a long time after death.
The fact that the metabolic processes do not cease immediately after death allows the investigators to classify the postmortem metabolites into three categories (Cat. I) the original basis set, (Cat. II) Basis extension (1), and (Cat. III) Basis extension (2). Cat. I includes N-acetyl-aspartate (NAA), N-acetyl-aspartyl-glutamate (NAAG), aspartate, lactate, trimethylammonium compounds (TMA), alanine, creatine, gamma-aminobutyric acid (GABA), myo-inositol, syllo- inositol, glutamine, glutamate, taurine, hypoxanthine (HX), and glucose. Cat. II includes acetate, ethanol, glutathione, glycerol, glycine, choline, phosphoethanolamine, valine, lipids, and lipid-like methyl groups. In addition to Cat. III, which includes free TMA, butyrate, propionate, isobutyrate, and succinate. Metabolites of Cat. I and Cat. II had substantially changed within 48 hours PM, while metabolites of Cat. III appeared prominently after 48 hours PM [9]. Therefore, many studies have been applied to different species, such as canines, bovines, porcines, rabbits, sheep, and humans, using H1-MRS to calculate PMI through a statistical analysis of various brain metabolite concentration changes at different intervals after death [10].
In the current work, we reviewed a comparison between conventional methods and H1-MRS for the estimation of PMI in six different species. We also showed the similarities and differences in the brain metabolite concentrations among the species. Limitations of H1-MRS studies and suggestions for improving the results in future experiments were also mentioned.
Methods
Search Strategy
We conducted an electronic search in ten electronic search engines: PubMed, Google Scholar, Scopus, Web of Science, Virtual Health Library (VHL), Popline, International
Scientific Indexing (ISI), WHO Global Health Library, New York Academy of Medicine Grey Literature Report (NYAM), and System for Information on Grey Literature in Europe (SIGLE). They were searched for relevant studies published before January 2023, as shown in Figure 1. We used the search terms “postmortem interval”, “Livor mortis”, “Rigor mortis”, “Body cooling”, “Putrefaction Signs”, and “proton-magnetic resonance spectroscopy”, and then the search string was modified to be suitable for each database. Reference lists of some published papers were examined for more sources not found in the database searches.
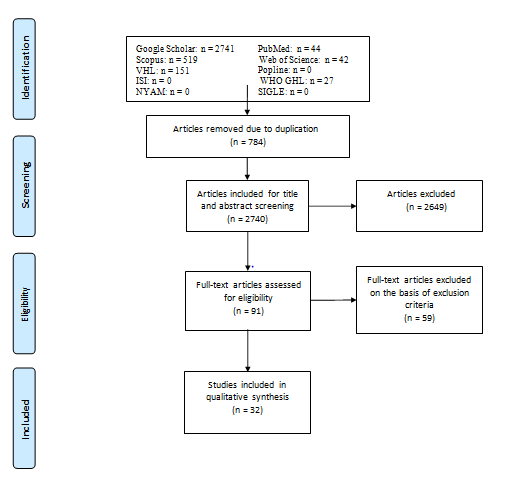
Study Inclusion Criteria and Study Selection
Only original articles (published before January 2023) concerning the use of conventional methods or H1-MRS in the determination of PMI in humans and different animal species were included in this systematic review. There was no restriction on a certain language, publication year, or country, but we limited the included studies to those trials that were applied away from the effect of ambient temperature on brain metabolite concentrations. Letters, editors’ correspondences, news, and studies whose data could not be reliably retrieved or extracted were excluded. A preliminary assessment of the titles and abstracts was performed to identify relevant articles. After that, full texts were screened, and the eligible texts were used in the data synthesis.
Data Extraction
The following data were extracted from the included studies: the first author’s name, year of publication, year of carrying out of the experiment, study design, country and city of origin, characteristics of human and animal models (species, age, gender, weight), cause of death, number of included subjects in each experiment, name of metabolites produced in each species, duration of spectroscopy measurement, and time interval from death until the occurrence of brain decomposition in all species. Some graphs were plotted to be used in data synthesis. In addition, the identification and elimination of overlapping datasets were performed under extensive care, especially for multiple studies containing the same species.
Data Analysis
Meta-analysis was inapplicable due to a lack of studies on the same species with the same circumstantial evidence and the same environmental conditions.
Quality Assessment
A total of 11 studies were identified to be assessed by using the Assessment of Diagnostic Accuracy Studies-2 (QUADAS-2)” tool, which is highly recommended by the Cochrane Collaboration for diagnostic test accuracy (DTA). QUADAS-2 addressed “risk of bias” and “applicability concerns” through 4 domains: patient selection, index test, reference standard, and flow and timing, as shown in Table 1. The quality of the included studies was evaluated as medium.
Notably, most studies did not compare the results of H1- MRS with the results of the conventional methods for PMI calculation, so “reference standard” and “flow and timing” were categorised as “unclear” risk of bias and applicability concerns.
| Study | Risk Of Bias | Applicability Concerns | |||||
|---|---|---|---|---|---|---|---|
| Patient Selection | Index Test | Reference Standard | Flow and Timing | Patient Selection | Index Test | Reference Standard | |
| Choe, et al. [11] | Low | High | Unclear | unclear | Low | High | Unclear |
| Michaelis, et al. [12] | Low | High | Unclear | unclear | Low | High | Unclear |
| Banaschak, et al. [13] | Low | High | Unclear | unclear | Low | Low | Unclear |
| Yang, et al. [14] | Low | High | Unclear | unclear | Low | Low | Unclear |
| Yang, et al. [15] | Low | High | Unclear | unclear | Low | Low | Unclear |
| Scheurer, et al. [1], (Sheep) | Low | High | Unclear | unclear | Low | Low | Unclear |
| Ith, et al. [16] (Sheep) | Low | High | unclear | unclear | High | Low | Unclear |
| Ith, et al. [5](Sheep) | Low | High | unclear | unclear | Low | Low | Unclear |
| Ith, et al. [9], (Human) | High | High | unclear | unclear | Low | Low | Unclear |
| Ith, et al. [16], (Sheep) | Low | High | unclear | unclear | Low | Low | Unclear |
| Ith, et al. [16], (Human) | Low | High | Low | Low | Low | Low | Low |
| Scheurer, et al. [10] | Low | High | unclear | unclear | Low | Low | Unclear |
| Musshoff et al. [17] | Low | High | unclear | unclear | Low | Low | Unclear |
Table 1: Quality Assessment of Studies Using the QUADAS-2 Tool.
Results
This section offers a review and analysis of results concerning the identification of PMI by using conventional methods and H1-MRS in canines, bovines, porcines, rabbits, sheep, and humans.
Canine
For almost more than a century, pathologists have depended on many conventional methods, such as alterations in body temperature, lividity, rigor mortis, and signs of putrefaction, to identify the time elapsed after death in different animals [11]. Some people used to own dogs in their houses, which is why animal cadavers might be found beside their dead owners in crimes. Therefore, studying the postmortem changes in canine cadavers has been a point of interest to help pathologists determine the PMI of dogs and consequently the PMI of their owners [12]. Based on that, PMI was determined as follows: during the 1st day PM, the rectal temperature of the canine was above the ambient temperature, the cornea dried out, rigor mortis started to develop in most muscle groups, and blue to green skin discolouration developed in the caudal abdomen and flank [13].
On the 3rd day PM, the rectal temperature equaled the ambient temperature, the upper eyes were shrunken, the rigidity of some joints was lost and skin discolouration of the abdomen and flank became deeper. On the 7th day PM, the eyes collapsed, rigor mortis was still present in the jaw and hind limbs, and signs of putrefaction showed great development. Finally, by three weeks PM, the cornea and adjacent membranes were colonised by molds and fungi, green, blue, and brown skin discolouration was so marked, and the epidermal layer was easily detached [13].
On another note, in 1995, Choe et al. investigated the alterations in brain metabolite concentrations in dogs before and after death until two days PM [14]. Dogs were treated with anaesthetic agents and then killed by intracardiac injection of 20 ml 10% KCl. The results revealed the presence of four premortem metabolites that were considerably high and characterised for the premortem period (livability) in dogs. They were N-acetyl-aspartate (NAA), creatine (Cr), choline (Cho) and inositol (Ino). There was no resonance signal of lactate before death.
Noticeably, just 30 minutes after death, a significant lactate signal appeared, giving a lactate/Cr ratio (w/w) from 2 to 2.86 mmol/kg, and within three hours PM, it increased to a range of 2.53 - 3.37 mmol/kg. After these early postmortem hours, the lactate/Cr ratio started to decrease gradually until reaching 2.02 - 2.94 mmol/kg at 24 hours PM (Table 2). In addition, the inositol/Cr ratio also increased to a range of 1.16 - 1.86 mmol/kg at three hours PM, but it kept increasing until reaching 1.43 - 3.11 mmol/kg at the end of the 1st day PM.
Considering the neuronal marker NAA, there was a gradual decrease in its concentration until it almost disappeared on the 3rd day PM. This might be attributed to its responsibility for the integrity of neuronal tissue, which was lost when ischemia occurred after death and continued to develop until the 3rd day PM. NAA and creatine concentrations were decreasing at the same time. This might explain why the NAA/Cr ratio was approximately constant and did not decrease substantially (Table 2). At 48 hours PM, autolysis occurred very rapidly in the neurons of the ventral thalamic nucleus and then in the small cells of the striatum, leading to the inability of H1-MRS to identify the signals of metabolites. Notably, slower decomposition occurred in the large pyramidal cells of the cortical layer of the parietal lobe, which were more durable [14]. Therefore, researchers turned to positioning the spectroscopy volume of interest in the brain cortex instead of the thalamus in their later studies [1, 5, 9, 10, 15, 16, 17].
| Metabolites’ ratios | Time interval | ||
|---|---|---|---|
| Premortem | 3 hPM | 24 hPM | |
| Lactate/Cr | Zero | 2.53 – 3.37 | 2.02 – 2.94 |
| Inositol/Cr | 0.69 – 1.21 | 1.16 – 1.86 | 1.43 – 3.11 |
| NAA/Cr | 0.97 – 1.37 | 1.12 – 1.7 | 0.86 – 1.22 |
Table 2: Statistical Evaluation of Metabolite Ratios in the Dog Brain by Using H1-MRS (P<0.05) [14].
(n =10). * (hPM) refers to hour/s postmortem. * (Cr) refers to creatine; (NAA) refers to N-acetyl aspartate. Table 2: Statistical Evaluation of Metabolite Ratios in the Dog Brain by Using H1-MRS (P<0.05) [14].
Bovine
There are no published data on the estimation of PMI by conventional methods in Bovine, as veterinary forensic studies on ruminants are lacking. However, it is known that pathologists rely on their experience to estimate it subjectively [13].
Therefore, we concluded the results of nuclear magnetic resonance (NMR) spectroscopy only. Bovine brain homogenate was used to represent each animal. Portions of approximately 4.0 g cortex were homogenised and stored in a refrigerator at 4°C until the end of proton NMR measurement at 195 hours PM [18]. Under refrigeration, the concentrations of lactate, creatine, myo-inositol, scyllo- inositol, glutamate, and glutamine remained constant from three hours PM until the end of the experiment, with ranges of 26.6 ± 2.2, 9.4 ± 0.6, 8.1 ± 0.7, 0.58 ± 0.04, 6.9 ± 0.4, and 2.4 ± 0.2 mmol/kg, respectively [18].
The high concentrations of lactate, creatine, and myo- inositol in the brain homogenate were indicative signs of death at three hours PM. In addition, the concentration of GABA, an inhibitory neurotransmitter in the brain tissue, at three hours PM ranged from 1.7 - 2.1 mmol/kg. At 8.5 hours PM, the concentration increased dramatically to 2.3 - 3.3 mmol/ kg, indicating that GABA is a characteristic brain metabolite for bovine brain homogenate at 8.5 hours PM. Additionally, there was a gradual increase in GABA concentration from the onset of death along the experimental time, reaching an average of 1.9 mmol/kg at three hours PM and 6.2 mmol/kg at 195 hours PM [18].
The decomposition of glycerophosphocholine (Gpc) into glycerol and choline after death resulted in an increase in the concentrations of glycerol and choline during the 195 hours of the experiment [18]. For example, at 22 hours PM, the concentration of glycerol ranged from 0.84 - 0.94 mmol/ kg, and at 31 hours PM, it increased to 0.95 - 1.15 mmol/ kg. Glycerol was a characteristic brain metabolite for bovine brain homogenate at that time range (22-31 hours PM).
At 147 hours PM– almost six days PM– the concentrations of choline and glycerol were identified as 1.35 - 1.87 mmol/ kg and 1.91 - 2.13 mmol/kg, respectively (Table 3).
At 195 hours PM, the three characteristic metabolites, namely, alanine, choline, and glycerol, were determined at concentrations of 3.2 - 3.4, 2.02 - 2.30, and 2.52 - 2.56 mmol/ kg, respectively. These specific concentrations appeared to confirm the beginning of the 9th day PM for the bovine bodies. Although the decrease in NAA concentration was insignificant during the first two days PM, it started to decrease gradually, reaching an average ranging from 3.1 - 3.7 mmol/kg on the 9th day PM. Notably, NAA is decomposed into its primary components acetate and aspartate, whose concentrations accordingly increased along the 195 hours PM [18].
| Metabolite | PMI | |||||
|---|---|---|---|---|---|---|
| 3 hPM | 8.5 hPM | 22 hPM | 31 hPM | < 6 days PM (147 hPM) | < 8 days PM (195 hPM) | |
| GABA | 1.7 – 2.1 | 2.3 – 3.3 | 3.3 – 4.5 | 3.8 – 4.8 | 4.8 – 6.2 | 5.5 – 6.9 |
| Glycerol | 0.43 – 0.71 | 0.64 – 0.76 | 0.84 – 0.94 | 0.95 – 1.15 | 1.91 – 2.13 | 2.52 – 2.56 |
| Choline | 0.47 – 0.53 | 0.48 – 0.64 | 0.54 – 0.94 | 0.78 – 1.1 | 1.35 – 1.87 | 2.02 – 2.3 |
| NAA | 7.5 – 8.7 | 7.1 – 9.5 | 6.9 – 8.5 | 7.7 – 8.1 | 3.7 – 5.7 | 3.1 – 3.7 |
Table 3: Metabolite Concentrations ((mmol/kg)) in Bovine Brain Homogenate (P<0.05) [18].
(n =24). * (hPM) refers to hour/s postmortem. Table 3: Metabolite Concentrations ((mmol/kg)) in Bovine Brain Homogenate (P<0.05) [18].
Porcine
Porcine is characterised by light rosy skin and less hair than other animals, so postmortem changes appear obviously on their skin after death. During the first 12 hours PM, lividity was prominent in the pig carcass with hypostatic pink skin discolouration, in addition to a beginning of rigor mortis in the muscle group. At the end of the 1st day PM, lividity was darker colored over the head, neck, and trunk, and rigor mortis extended to include the whole body muscles. Purple to black discolouration was observed in the lower abdominal region on the 2nd day PM. The marbling of the trunk region and early onset of bloating were marked signs of the beginning of the decomposition stage on the 3rd day PM. Then, blisters filled with maggots appeared on the trunk region, and the skin started to slip. Towards the end of this stage, bloating developed in the neck, hair was lost, and brown discolouration developed around the edges. These changes were observed in spring/summer in South Africa, and there is no doubt that the rate of corpse decomposition is faster in summer and spring than in winter and autumn [19].
Magnetic resonance spectroscopy was also applied to porcine in a study by Banaschak, et al [13].
Heads of young pigs were obtained from an abattoir, processed, and stored at 21 ± 1°C for three weeks.
The analysis of resonance spectra of pig brain metabolites revealed the presence of alanine, acetate, and choline signals throughout the experiment. In the first four days PM, prominent concentrations of NAA, creatine, and lactate were determined, while succinate and free trimethylammonium (f TMA) began to appear at the beginning of the 5th day PM. The disappearance of NAA at approximately five days and a half PM (~130 hours PM) and creatine at seven days PM (~170 hours PM) marked their postmortem times. Interestingly, lactate was the most durable metabolite, which disappeared almost at the first third of the 9th day PM (~200 hours PM) [15].
Rabbits
The decomposition rate and insect succession of rabbit carcasses occur simply and more rapidly than those of large- weighted carcasses [20]. Corpse decomposition can be divided into five stages (fresh, bloat, active decay, advanced decay, and dry) [21, 22]. The fresh stage lasted approximately 12 hours for outdoor-placed carcasses and 36 hours for indoor-placed carcasses. Then, the bloat stage followed the fresh stage until the 1st and 2nd day PM. High decomposition of outdoor and indoor-placed carcasses marked the active decay stage on the 3rd day PM, and skeletonisation of both carcasses marked the advanced decay stage on the 5th day PM. Finally, the dry stage developed on the 19th day PM for outdoor-placed carcasses and on the 16th day PM for indoor- placed carcasses [22]. Notably, calliphorid flies were the first forensic insects to arrive and breed on rabbit carcasses in all seasons of the year [23].
In addition, regarding the application of the analytical technique H1-MRS on the brains of rabbits in the postmortem period, it showed a substantial decrease in the relative concentrations of NAA, choline, and creatine within 24 hours PM [24, 25]. PMI was estimated through two independent equations using the NAA/Cr and Ch/Cr ratios as independent variables, and the average deviations of the two estimated PMIs were 2.1 - 37.9 min and 1.69 - 40.87 min, respectively [24]. This indicated that NAA/Cr and Ch/Cr were significant metabolic ratios for the estimation of PMI in rabbits.
Sheep
As mentioned before, forensic studies on ruminants are rare. Therefore, we concluded the available published data on sheep as follows: Lucilia cuprina was the first forensic fly that arrived and breaded on sheep cadavers upon death. Minutes- hours PM, it was attracted to sheep cadavers and started a colonisation cycle (egg- maggot- pupa- adult). It is known as a “sheep green bottle” and is only found on corpses located in a drier climate, as it is a very specific insect. Pathologists have used it as a tool for the estimation of PMI by classifying the age of its developmental stages [26].
On another note, several studies were conducted on sheep models for PMI calculation through alterations in their brain metabolite concentrations. Sheep were slaughtered, and heads with attached spinal cords were sealed by plasticine immediately after decapitation to prevent any bacterial invasion or loss of CSF, placed in plastic containers, and stored at room temperature (21 ± 3°C) for investigation [1, 5, 9, 10, 16].
In the premortem period, the analysis of results revealed the presence of propionate and butyrate signals at concentrations of 1.7 mmol/kg and 3.1 mmol/kg, respectively, but no signals of isobutyrate and f TMA [9]. At seven hours PM, the propionate signal disappeared, the butyrate concentration decreased to almost 2.7 mmol/kg, and isobutyrate and f TMA signals remained absent (Table 3). Nevertheless, there were clear signals of creatine and choline compounds in that time range [9, 16]. NAA had a predominant signal with an average concentration of 5-6 mmol/kg a few hours PM. However, it decreased gradually to
3 - 5 mmol/kg at the end of the 1st day PM and to 1 - 3 mmol/ kg on the 2nd day PM [3, 4, 10].
On the 3rd day PM, the propionate concentration reached approximately 3.6 mmol/kg and continued to increase significantly (p < 0.05) until the end of the experiment. fTMA had a prominent signal with a concentration ranging from 6 - 40 mmol/kg from the 4th day to the 7th day PM [9]. In parallel, when the concentrations of alanine, acetate, and lactate increased, the concentrations of NAA, total creatine, and choline decreased over time [16].
On the 9th day PM, there were substantial signals of alanine, acetate, and succinate, while NAA, total creatine, and lactate almost disappeared. On the 15th day PM, there was a noticeable increase in the concentrations of propionate, butyrate, fTMA, and isobutyrate, in addition to acetate, which broke a record, reaching a concentration greater than 200 mmol/kg [16].
| Metabolite | PMI | |||
|---|---|---|---|---|
| 7 – 8 hPM | 2nd - 3rd-day PM | 4th – 5th-day PM | 12th – 15th-day PM | |
| Propionate | Zero | 0.2 – 3.6 | 4.1 – 9.7 | 12.7 – 31.3 |
| Butyrate | 2.4 - 3 | 2.4 – 3.6 | 0.7 – 4.9 | 26 - 62 |
| Free TMA | Zero | Zero | 24-Feb | 26 – 38 |
Table 4: Metabolite concentrations (mmol/kg) estimated by NMR in the sheep brain [9].
*(Total n = 9). * (hPM) refers to hour/s postmortem. Table 4: Metabolite concentrations (mmol/kg) estimated by NMR in the sheep brain [9].
Human
Using conventional methods for the identification of PMI is very common and objective in humans. As livor mortis determined PMI from half an hour to approximately eight hours after death according to its degree [4]. Additionally, algor mortis developed from half to an hour postmortem and continued until approximately 10-12 hPM when the body temperature equaled the environmental temperature. In a parallel way, rigor mortis started within almost two hPM and developed from head to feet in 12 hours, stunned for an additional 12 hours, and then disappeared from feet to head from 24 hPM to 36 hPM [4].
From a forensic entomologic perspective, blowflies are deposited on body orifices at highly variable times depending on the conditions of the cadaver and the environment. Overall, the life cycle of blowflies to become adults is approximately 18 to 24 days, divided into life stages referring to different periods of death [23].
In addition, considering the application of H1-MRS on human subjects, it had some limitations, including number, measurement, and ethical considerations. In spite of the lack of trials and cadavers, researchers could drive promising results for the calculation of PMI using H1-MRS. Human subjects used in these studies were those reported dead in closed areas at room temperature, transported to the Institute of Legal Medicine, and stored at 4°C until scanned [1, 9].
The spectroscopic examination and analysis took almost a couple of hours, which was much shorter than any other conventional criminological investigation [1]. In the early stage, there was a decrease in NAA and NAAG, an obvious increase in lactate and acetate [17], and detectable alterations in the metabolites of Cat. II [9]. Additionally, it was concluded that the overall absence of isobutyrate, fTMA, and propionate signals referred to death from five to six hPM (table 4) [10]. Isobutyrate and fTMA signals appeared after death, as they were not found in the live human brain [9]. The isobutyrate signal started to appear two days PM, and the fTMA signal appeared on the 3rd day PM [10]. After the first three days, creatine showed a decrease followed by disappearance, while acetate and fTMA showed a dramatic increase [17].
| Metabolite | PMI | |||
|---|---|---|---|---|
| 5 hPM | 3rd day PM | 4th day PM | 6th day PM | |
| Propionate | Zero | 2.1 | 8 | 12.3 |
| Free TMA | Zero | Zero | 7.1 | 15.5 |
Table 5: Metabolite Concentrations (mmol/kg) Estimated by H1-MRS in the Human Brain [9].
(n =4). *(p < 0.05; Z-test, one-tail; calculated from group deviation when n > 1). Table 5: Metabolite Concentrations (mmol/kg) Estimated by H1-MRS in the Human Brain [9].
Discussion
Conventional methods such as the assessment of livor and rigor mortis, core body temperature, and putrefaction signs are considered objective parameters for the estimation of PMI in many species worldwide. However, in two-day aged cadavers or more, it would be a difficult mission for these methods to be feasible and would identify PMI with a long time interval and a low accuracy extent [1, 9]. Using H1-MRS, regardless of the time elapsed after death or occurrence of putrefaction, has improved a way towards solving this problem [1]. It is an easily applicable technique that can run on the same magnetic resonance imaging (MRI) system, penetrate the skull, determine brain metabolites, and analyse their relative concentrations without any invasion or autopsy [1]. In addition, through spectroscopy, we could perform a statistical analysis of metabolite concentration changes, giving an accurate confidence interval, unlike conventional methods that cannot include such statistical functions [1, 10]. Spectroscopy could also determine the metabolite concentrations in other organs, such as muscles and bovine brain homogenate, extending the PMI calculation to be determined until 15 days PM beyond the achievable range calculated by conventional methods [5]. Moreover, high-resolution angle spinning spectroscopy could provide an interesting method for the examination of excised tissue because it only requires a small amount of unprocessed tissue [1]. Finally, H1-MRS increased the accuracy extent of PMI estimation to hours instead of the long weekly PMIs determined by the conventional methods in putrefied cadavers [1].
Consequently, the application of H1-MRS alone or add- on conventional methods would be a “gold standard” for the estimation of PMI in many species.
In light of previous studies using H1-MRS, we analysed their results and concluded that the neuronal markers N-acetyl-aspartate (NAA) and N-acetyl-aspartyl-glutamate (NAAG) decreased rapidly after death, and they might be considered indicators for the determination of PMI until 70 hours PM in sheep and humans [4, 10] and until 130 hours PM in porcines [15]. Additionally, the live brain of animals produces low concentrations of butyrate and propionate [9], and within 48 hours PM, these metabolites generally increase until 400 hours PM, as they arise from bacterial metabolism [9].
In humans and sheep, one of the most interesting findings was the two brain metabolites known as isobutyrate and f TMA, which are not produced normally during life, but their production started to appear after 24 hours PM and 48 hours PM, respectively [9]. Some brain metabolites were higher in humans than in any other animal species, such as NAA in the premortem period [10] and free glycerol in the postmortem period. Free glycerol showed a clear significant signal in the resonance spectrum of the human brain at 48 hours PM [18].
In sheep, the highest concentration of all brain metabolites was recorded for acetate, which reached > 200 mmol/kg on the 15th day PM [9]. Studies have shown that acetate initially originates from the degradation of NAA and later from bacterial decomposition. The concentration increased gradually after death until reaching a peak on the 9th day PM and then stayed stationary until the 17th day PM [1].
Interestingly, a high correlation coefficient (r) was reported between the predicted time estimated by H1-MRS and the real postmortem time in five sheep brain metabolites, including acetate, alanine, free trimethylamine, propionate, and butyrate, as (r = 0.97) until 250 hours PM and (r = 0.93) until 300 hours PM [1, 3, 4, 10]. In addition, many metabolites produced in sheep brains were detected in human brains, and their concentration changes formed a similar resonance spectrum, which indicated that sheep could be a valid model for humans in that aspect [1].
Despite all previous studies and their promising results, investigators faced many limitations. First, an inadequate number of available subjects due to related ethical aspects reduced the investigation of diversified causes of death and estimation of their relative PMI. Second, the absence of a magnetic resonance system as a research tool in research centers led to difficult feasibility, as MRI and MRS have been used as diagnostic tools in hospitals for living people. Third, the formation of gas bubbles in brain tissue complicated the selection of voxels with homogenous brain tissue [14, 15, 16] and deteriorated the measurement of some metabolites, leading to the inability to follow the changes in brain metabolite concentrations for a long time postmortem.
Therefore, there are important precautions that should be considered in further work to ensure accurate and reliable results. Examples include using a H1-MRS system specialised for research to increase the time estimated after death without intervals and positioning the volume of interest in the cortical pyramidal cells of the parietal lobe instead of the thalamus of the brain, as the pyramidal cells are more durable [10]. It should be mentioned that the concentration of the same metabolite varies among different brain parts, as NAA and creatine concentrations varied in a prominent way among the cortex, cerebellum, and white matter of the rabbit brain [27]. In addition, cadavers already found dead are preferable to those slaughtered or killed by researchers to avoid the risk of bias caused by knowing the real-time of death and the overestimation of spectroscopy accuracy. Additionally, using fresh experimental cadavers without preservation in a refrigerator would lead to realistic data at room temperature. Cooling of whole cadavers or isolated heads of animals slows brain tissue decomposition and changes the lifespan of each postmortem metabolite [1]. The declaration of the causes of death, especially in human cases, and the preparation of a considered number of subjects with the same cause of death are essential for providing concise and evidence-based results [1]. On another note, treatment of animals with brain-targeted drugs before the beginning of experiments, such as narcotics, sedatives…etc. might alter the relative metabolite concentrations and increase the possibility of false results [28].
Additionally, the measurement of brain metabolite concentrations in a whole intact body is preferable to using isolated heads [16]. As in sheep, there were more metabolites in isolated head brains than in intact body brains, including propionate, butyrate, and acetate. However, investigators could find fTMA and lactate in intact bodies’ brains at late PMI. This change in metabolites and their relative concentrations might result from the migration of gut bacteria to the brain of intact cadavers to play a role in decomposition after death [16]. Gut bacteria have a significant role in the determination of PMI, as shown by Bonte, et al. [29, 30] several years ago. Recently, it was supposed that they contribute to the determination of types of brain metabolites through hormonal transmitters in humans and other animals, but this issue is still under study [31, 32].
Estimation of PMI with an accurate, rapid, and feasible method is one of the major calls in the world of forensic medicine, and developing more trials by using H1-MRS will help in the achievement of this goal through simple inverse mathematical functions allowing the determination of PMI by a single MRS measurement.
Conclusion
Proton-magnetic resonance spectroscopy (H1-MRS) could identify the post-mortem interval (PMI) of many different species accurately, and beyond the achievable range by the known conventional methods.
Acknowledgements
We would like to thank Prof. Burkhard Madea, Institute of Forensic Medicine, University of Bonn, Germany, for providing us with his valuable recommendations. We also thank Prof. Maha Ghanem, Forensic Medicine Department, Faculty of Medicine, University of Alexandria, Egypt, for guiding us with her advice.
Funding
No funding was received for conducting this study.
Disclosure of Potential Conflicts of Interest
The authors declare that they have no conflicts of interest.
Research Involving Human Participants and/or Animals
The study is a systematic literature review, so this section is not applicable.
Informed Consent
This section is not applicable in this study.
Ethical Approval
The authors used PRISMA checklist and QUADAS-2 tool to conduct a high-quality study.
Data Availability Statements
All data generated or analysed during this study are included in the published article.
References
-
Scheurer E, Ith M, Dietrich D, Kreis R, Hüsler J, et al. (2005) Statistical evaluation of time‐dependent metabolite concentrations: estimation of post‐mortem intervals based on in situ1H‐MRS of the brain. NMR in Biomedicine 18(3): 163-172.
-
Campobasso CP, Vella GD, Introna F (2001) Factors affecting decomposition and Diptera colonization. Forensic science international 120(1-2): 18-27.
-
Madea B (2005) Is there recent progress in the estimation of the postmortem interval by means of thanatochemistry?. Forensic science international 151(2-3): 139-149.
-
Madea B (2016) Methods for determining time of death. Forensic Science Medicine and Pathology 12(4): 451- 485.
-
Ith M, Bigler P, Scheurer E, Kreis R, Hofmann L, et al. (2002) Identification of metabolites emerging during autolysis and bacterial heterolysis of decomposing brain tissue by 1H-MRS in situ and in vitro. Proc Int Soc Magn Reson Med.
-
Daldrup T (1978) The significance of the formation of α-and γ-aminobutters during the composition of any human organic material. A Possibility of Deadline Determination. 57. Tgg Dtsch Ges Rechtsmed, Düsseldorf.
-
Daldrup T (1981) Towards Kinetics of the Postmortem Bacterial Glutamine Metabolism of the glutamic acid in brain. Z Rechtsmed 86(3): 195-203.
-
Daldrup T (1983) Practical experience with determination of leachate by analysis of bacteria and metabolic products. Journal of Legal Medicine 90: 19-25.
-
Ith M, Bigler P, Scheurer E, Kreis R, Hofmann L, et al. (2002) Observation and identification of metabolites emerging during postmortem decomposition of brain tissue by means of in situ 1H‐magnetic resonance spectroscopy. Magnetic resonance in medicine 48(5): 915-920.
-
Scheurer E, Ith M, Dietrich R, Kreis R, Hüsler J, et al. (2003) Statistical evaluation of 1H-MR spectra of the brain in situ for quantitative determination of postmortem intervals (PMI). Proc Int Soc Magn Reson Med 11: 569.
-
Choe BY, Gil HJ, Suh TS, Shinn KS (1995) Postmortem metabolic and morphologic alterations of the dog brain thalamus with use of in vivo 1H magnetic resonance spectroscopy and electron microscopy. Investigative radiology 30(5): 269-274.
-
Michaelis T, Helms G, Frahm J (1996) Metabolic alterations in brain autopsies: proton NMR identification of free glycerol. NMR in biomedicine 9(3): 121-124.
-
Banaschak S, Rzanny R, Reichenbach J, Kaiser W, Klein A (2005) Estimation of postmortem metabolic changes in porcine brain tissue using 1H-MR spectroscopy— preliminary results. International Journal of legal medicine 119(2): 77-79.
-
Yang TT, Li ZW, Liu L, Zheng N (2007) Estimation of postmortem interval with multivoxel-voxel proton 1H- MR spectroscopy. Fa Yi Xue Za Zhi 23(6): 405-408.
-
Yang TT, Li ZW, Liu L, Zhen N (2008) Estimation of postmortem interval with single-voxel proton 1H-MR spectroscopy at different temperature. Fa Yi Xue Za Zhi 24(2): 85-89.
-
Ith M, Kreis R, Scheurer E, Dirnhofer R, Boesch C (2001) Using 1H-MR Spectroscopy in Forensic Medicine to Estimate the Post-Mortem Interval: A Pilot Study in an Animal Model and its Application to Human Brain. Proc Intl Soc Mag Reson Med 9: 388.
-
Musshoff F, Klotzbach H, Block W, Traeber F, Schild H, et al. (2011) Comparison of post-mortem metabolic changes in sheep brain tissue in isolated heads and whole animals using 1H-MR spectroscopy—preliminary results. International Journal of legal medicine 125: 741- 744.
-
Listos P, Gryzinska M, Piorkowski J, Teresinski G, Buszewicz G, et al. (2016) Post-Mortem Estimation of Time of Death of Dogs Based on Measurements of Kidney Temperature in Comparison with Rectal Temperature. Acta Veterinaria 66(1): 76-88.
-
Proctor KW, Kelch W J, New JC (2009) Estimating the time of death in domestic canines. J Forensic Sci 54(6): 1433-1437.
-
Erlandsson M, Munro R (2007) Estimation of the post- mortem interval in beagle dogs. Sci Justice 47(4): 150- 154.
-
Keough N, Myburgh J, Steyn M (2017) Scoring of Decomposition: A Proposed Amendment to the Method When Using a Pig Model for Human Studies. Journal of Forensic Sciences 62(4): 986-993.
-
Wang Y, Ma MY, Jiang XY, Wang J, Li L, et al. (2017) Insect succession on remains of human and animals in Shenzhen, China. Forensic Sci Int 271: 75-86.
-
Aly M, Osman K, Galal F, Hassan G (2017) Comparative Study on Outdoor and Indoor Forensic Insects Encountered on Rabbit Corpses in Upper Egypt. Journal of Pharmacy and Biological Sciences 12(3): 41-54.
-
Boulkenafet F, Berchi S, Lambiase S (2015) Preliminary study of necrophagous Diptera succession on a dog carrion in Skikda, North-east of Algeria. Journal of Entomology and Zoology Studies 3(5): 364-369.
-
Sharma M, Singh D (2015) Historical approach of Forensic Entomology: A review. International Journal of Advanced Research in Science, Engineering and Technology 2(9): 838-845.
-
Bansode S (2017) Effect of seasonal variations on the life cycle of Lucilia cuprina (Wiedemann, 1830) (Diptera: Calliphoridae). Journal of Entomology and Zoology Studies 5(3): 1518-1522.
-
Petroff OA, Ogino T, Alger JR (1988) High‐resolution proton magnetic resonance spectroscopy of rabbit brain: regional metabolite levels and postmortem changes. Journal of Neurochemistry 51(1): 163-171.
-
McGowan JC, Hill C, Mastrodonato A, LaGamma CT, Denny CA, et al. (2018) Prophylactic ketamine alters nucleotide and neurotransmitter metabolism in brain and plasma following stress. Neuropsychopharmacology 43: 1813- 1821.
-
Bonte W (1978) Postmortem Protein Catabolism: Experimental Investigations into Problems of Forensic Leichenzeit Determination. Germany.
-
Daldrup T, Huckenbeck W (1984) Bedeutung des FäulnisbakteriumsClostridium sordellii für die Leichenaltersbestimmung. Z eitschrift für Rechtsmedizin 92: 121-125.
-
Nithianantharajah J, Balasuriya GK, Franks AE, Hill- Yardin EL (2017) Using Animal Models to Study the Role of the Gut–Brain Axis in Autism. Current Developmental Disorders Reports 4(2): 28-36.
-
Swann J, Garcia-Perez I, Braniste V, Wilson ID, Sidaway JE, et al. (2017) Application of 1H NMR Spectroscopy to the Metabolic Phenotyping of Rodent Brain Extracts: A Metabonomic Study of Gut Microbial Influence on Host Brain Metabolism. Journal of Pharmaceutical and Biomedical Analysis 143: 141-146.
- Forensic Implications of Adverse Drug Reactions in Schizophrenia A Case Series
- Narcotics and Digital Forensics: Bridging Crimes in the Digital Age
- Ethics in Forensic Psychiatry: Principles, Dilemmas, and Human Rights
- Impact of Acute Stress on Attentional Orienting to Social Cues
- Head Injury and Intracranial Hemorrhage in Western Region of Libya
- A Forensic Study on Handedness: Examination of Handwriting Features in Right and Left Handed Writers