Glycoprotein Producing AM Fungi lifecycle and Potential Role in Agricultural Plant Lifespan and Global Environmental Changes for Sustainable Green Technology
The fungi forming arbuscular mycorrhizal fungi (AM fungi) are obligate symbionts microbe that predominates with inside the roots and soil of land flora. AM fungi are monophyletic group of phylum Glomeromycota. The AM fungi now no longer simplest arise in roots however additionally produce hung networks of extraradical mycelium (ERM) indeterminate length, spreading from host roots into the encompassing soil and establishing belowground interconnections amongst plants belonging to the identical or to unique taxa, in which they form a dense mycelial network (MNW). Reproduction in AM fungi takes place via asexual spores, which might be produced at the hyphae tips. These thick-walled spores stay with inside the soil for prolonged periods of time. AM fungi facilitate host plants to develop vigorously beneath stressful situations with the aid of using mediating a chain of complicated conversation occasions among the plant and the AM fungus leading to enhanced photosynthetic rate and other gas exchange in addition to improved water. AM fungi provide key nutrients to host plant lifespan. AM fungi transfer phosphorus (P), other macro and micro nutrients, along with water to the hosts of grassland and tropical forest plants in exchange for sugars. AM fungi form a network of filaments that associate with plant roots and draw nutrients from the soil that the root system would not be able to access otherwise. AM fungus plant alliance stimulates plant growth and accelerates root improvement in nearly all crops. The AM fungal extraradical hyphae (ERH) alongside glomalin connected soil protein (GCSP) extensively influence the soil carbon dynamics (SCD) and enhance soil and plant health. They additionally increase the resistance in plants against plant pathogens and surface area of root system for higher absorption of nutrients from soil. Therefore, they can be used as biofertilizer in addition to biocontrol agent. Present manuscript delineated the prospective of AM fungi as a biostimulants can potentially strengthen plants' adaptability to converting Agriculture system for green technology.
Introduction
Fungus is a microscopic microorganism of the group eukaryotes that consists of yeasts, moulds and mushrooms.
These organisms are very small requiring a microscope for thorough observation. They are globally plentiful and found in a huge variety of habitats. Numerous useful fungal species to be had in hemisphere that helped shape civilisation and fungi have had a considerable impact on human and plant lifespan. Plants began out setting up throughout terrestrial environments more than 430 million years ago; they had been observed via way of means of a symbiotic fungus, mycorrhizae. AM fungus (Endomycorrhiza), are grouped in a monophyletic group, the phylum Glomeromycota consists of all known AM fungi has coevolved with the majority of plant ever since. Considering the lengthy evolutionary records of mycorrhizae, it is not surprising that the mycorrhizal association is present in more than 95% of all vascular plant, as AM fungi apparently lacks host specificity. AM fungi are a root endosymbiosis among plants and glycoprotein producing fungi (GPPF). It is the greatest widespread terrestrial plant symbionts, enhancing water and mineral nutrients uptake to the plant. Mycorrhizae display many characteristics of primitive fungi: they produce spores, display little diversification, does now no longer have a sexual reproductive cycle, and cannot alive without living host. There are various types of mycorrhizae found in hemisphere. Ectomycorrhiza rely on fungi enveloping the roots in a sheath (mantle) and a Hartig net of hyphae that extends into the roots among cells in forest plants. The fungal partner can belong to the Ascomycota, Basidiomycota, or Zygomycota. In a second type, the Glomeromycota fungi form vesicular arbuscular interactions with AM fungi. AM fungi form arbuscules that penetrate root cells and are the site of the metabolic exchanges among the fungus and the host plant. The arbuscules (pint-sized trees like structure) have a shrub like appearance. Orchids rely on a third type of mycorrhiza. Orchids are epiphytes that form small seeds without much storage to sustain germination and growth. Their seeds are not germinated without a mycorrhizal partner (Basidiomycete). After nutrients in the seed are depleted, fungal symbionts support the growth of the orchid by providing necessary carbohydrates and minerals. A few orchids continue to be mycorrhizal association throughout their lifespan. AM fungi are obligate biotrophs feeding simplest on the products of photosynthesis of their living hosts. Generally, the fungi are not specialized to their potential hosts, although some plant species more favour the development of these fungi than others [1, 2, 3, 4, 5, 6]. The fungi belong to the most commonly occurring soil microorganisms of the world and are associated with plants of the earth together with angiosperms, gymnosperms and pteridophytes having roots, in addition to the gametophytes of a few mosses, lycopods, and Psilotalus, which do now no longer have real roots [2].
Various literature imply that AM fungi increase the root absorptive area and subsequently the plant nutrition’s [7, 8, 9, 10, 11], influence succession of plant communities [12], their competitiveness [13, 14] and phenology [13], equalize the level of nutrition of co-existing plants by formation of hyphal bridges transferring nutrients among them [15], and increase soil structure through binding sand grains into aggregates by ERH [16, 17, 18, 19]. AM fungi increased the tolerance of plants to heavy metals [20, 21, 22, 23, 24]; water stresses [25], in addition pathogenic fungi and nematodes [26, 27, 28, 29]. The requirement of AM fungi for as much as 20% of host photosynthate for establishment and maintenance is nicely accepted [30, 31, 32]. Keeping in view, the importance of AM fungi and additionally the contemporary studies improvements related to their applications in agriculture and forestry, this manuscript focuses on the lifespan and potential role of AM fungi as biofertilizer within the regulation of plant growth, development, with improved nutrient uptake beneath nerve-racking environments, overall crop improvement and converting universal sustainable Agriculture for green technology.
Background of AM Fungi
AM fungi are soilborne mutual symbiotic fungi that could extensively enhance plant nutrients uptake and resis- tance to numerous abiotic stresses. AM fungi include of intr- aradical hyphae (IRH) in the roots and extraradical hyphal (ERH) structures outside the roots. The IRH structures are related to arbuscules, vesicles, and IRH. The ERH structures are spores, and auxiliary cells are formed members of the genera Gigaspora, Pacispora, and Scutellospora. Haustori- ums like arbuscules are the main sites of nutrient exchange among a host plant and a fungus [6, 18, 20, 24, 32]. They are formed inside the cells of the internal root cortex (IRC) [1, 4, 5, 6, 33, 34] and are indicators of active lively mycorrhizae. Ar- buscules vary in morphology, relying at the commonplace association of the arbuscular fungal species [35]. Fungi of the genera Acaulospora, Archaeospora, Ambispora, Diversis- pora, Entrophospora, Glomus, Intrapora, Kuklospora, Pacis- pora, and Paraglomus produce arbuscules with cylindrical or slightly flared, slender trunks, whose branches steadily taper in width towards tips. Arbuscules of members of the gen- era Gigaspora and Scutellospora usually have swollen trunks with branches tapering abruptly at tips. Globose or ovoid, thin-walled vesicles are storage organs filled with lipids and glycolipids [36]. AM fungal vesicles are origin an intercalary swelling in the root’s tips of IRH. Vesicles can be inter or in- tracellular and can be found in both the internal and the outer layers of the cortical parenchyma. In Glomus species vesicles are mostly ellipsoid, while the ones of Acaulospora, Entro- phospora, and Kuklospora highly vary in shape and often have knobs and concavities on their surface [35]. Vesicles are nev- er produced by members of the genera Gigaspora and Scutel- lospora. Members of the genera Archaeospora, Intraspora, and Paraglomus not often produce vesicles. Intercellular hy- phae (ICH) in roots additionally comprise storage materials and take part in transportation of the substances absorbed by ERH from the soil to arbuscules or directly to root cells of the host plant [1, 4, 6, 7]. IRH can be straight or with H or Y shaped branches. They might also form coils, whose fre- quency of incidence relies upon on their position in a root and the generic affiliation of the arbuscular fungal species [35]. Generally, coils extra abundantly arise at entry points. IRH of Glomus species are infrequently coiled in the other regions of an AM fungal root. In contrast, coils produced by species of the other genera of AM fungi usually are abundant and evenly distributed along mycorrhizal roots. AM fungi additionally vary in the degree of evenness of distribution along roots and the intensity of staining. The distribution of AM fungal structures of members of the genera Ambispora, Archaeospora, Acaulospora, Diversispora, Entrophospora, In- traspora, Kuklospora, and Paraglomus is patchy, while that of mycorrhizae of the genera Gigaspora, Glomus, Pacispora, and Scutellospora commonly is continuous. The strength of staining of AM fungi of the genera Ambispora, Archaeos- pora, Diversispora, Intraspora, and Paraglomus could be very faint to faint, the ones of Acaulospora, Entrophospora, and Kuklospora faint to moderate, those of Glomus dark, and those of Gigaspora, Pacispora and Scutellospora very dark [37, 38]. A majority of the species of AM fungi belong to the sub-phylum Glomeromycotina, of the phylum Mucoro- mycota [39]. Four orders of AM fungi, namely, Glomerales, Archaeosporales, Paraglomerales, and Diversisporales, had been recognized on this sub-phylum that still consist of 25 genera [40]. They are obligate biotrophs and ingest plant photosynthetic products [41] and lipids to perform their life- cycle [42]. AM fungi mediated growth promotion is not only by enhancing water and mineral nutrients uptake from the adjoining soil however additionally via way of means of safe- guarding the plants from fungal pathogens [27, 29, 43, 44]. Therefore, AM fungi are critical endosymbionts playing an effective role in plant productivity and the functioning of the ecosystem for sustainable crop improvement.
Paleobiology of AM fungi
Paleobiological and molecular evidence imply that AM fungi are an ancient symbiosis that originated at more than 460 million years ago. AM fungal symbiosis is ubiquitous among terrestrial plants, which indicates that mycorrhizas were present in the early ancestors of extant global plants. This positive association with plants can also additionally have facilitated the development of terrestrial plants. The Rhynie chert of the lower Devonian has yielded fossils of the earliest land plants where in AM fungi have been observed [45]. The Early Devonian noticed the development of terrestrial flora. Plants of the Rhynie chert from the Lower Devonian have been determined to contain structures resembling vesicles and spores of present Glomus species. Colonized fossil roots had been observed in Aglaophyton foremost and Rhynia, which can be ancient plants possessing characteristics of vascular plants and bryophytes with primitive protostelic rhizomes [46]. IRH was observed in root intracellular spaces, and arbuscules were observed with inside the layer thin wall cells similar to palisade parenchyma. The fossil arbuscules appear very much similar to those of existing AM fungi [46]. The cells containing arbuscules have thickened walls, which can be additionally observed in extant colonized cells. Mycorrhizas from the Miocene show a vesicular morphology closely resembling that of present Glomerales. This conserved morphology can also additionally reflect the ready availability of nutrients provided by the plant hosts in each modern and Miocene mutualism [45]. However, it could be argued that the efficacy of signalling approaches is probable to have evolved since the Miocene, and this can’t be detected in the fossil record. A fine-tuning of the signalling processes could enhance coordination and nutrient exchange among symbionts at the same time as growing the fitness of each the fungi and the plant symbionts.
Molecular Evidence of AM Fungi
Increased interest in AM fungal symbiosis and the improvement of sophisticated molecular techniques has led to the rapid improvement of genetic evidence. Wang, et al. [47] investigated plant genes inclusive of DMI1, DMI3; IPD3 involved in communication with order Glomales fungal partners. These three genes can be sequenced from all major clades of modern land plants, including liverworts, the maximum basal group, and phylogeny of the three genes proved to agree with then current land plant phylogenies. This means that mycorrhizal genes ought to be present with inside the common ancestor of land plants, and they ought to be vertically inherited considering plants colonized land [47]. Distribution and Habitat of AM Fungi AM fungi are found in all soils as fungal spores, and lack any intrinsic movement capabilities. Spores are typically present in the rhizosphere soil region which is usually the middle region of soil, in the immediate area of a plant root system and soil associated microorganisms. However, once in the vicinity of a nearby host plant’s root system and under the correct conditions spore germination can initiate. This result in the growth of hyphae the ones branch out to close by host plant root systems. Whereby, a mutual symbiotic relationship establishes among the two organisms.
Reproduction and Lifecycle of AM Fungi
Reproduction in AM fungi happens via spores that are produced at the hyphal tips. These thick-walled spores stay with inside the soil for extended periods of time. AM fungal spores had been capable of germinate and bring produce hyphae with living host.
Presymbiosis of AM Fungi
The improvement of AM fungi earlier than root colonization (RC), referred to as presymbiosis, comprises three stages including as spore germination, hyphal growth and host recognition and appressorium formation.
AM Fungal Spore Germination
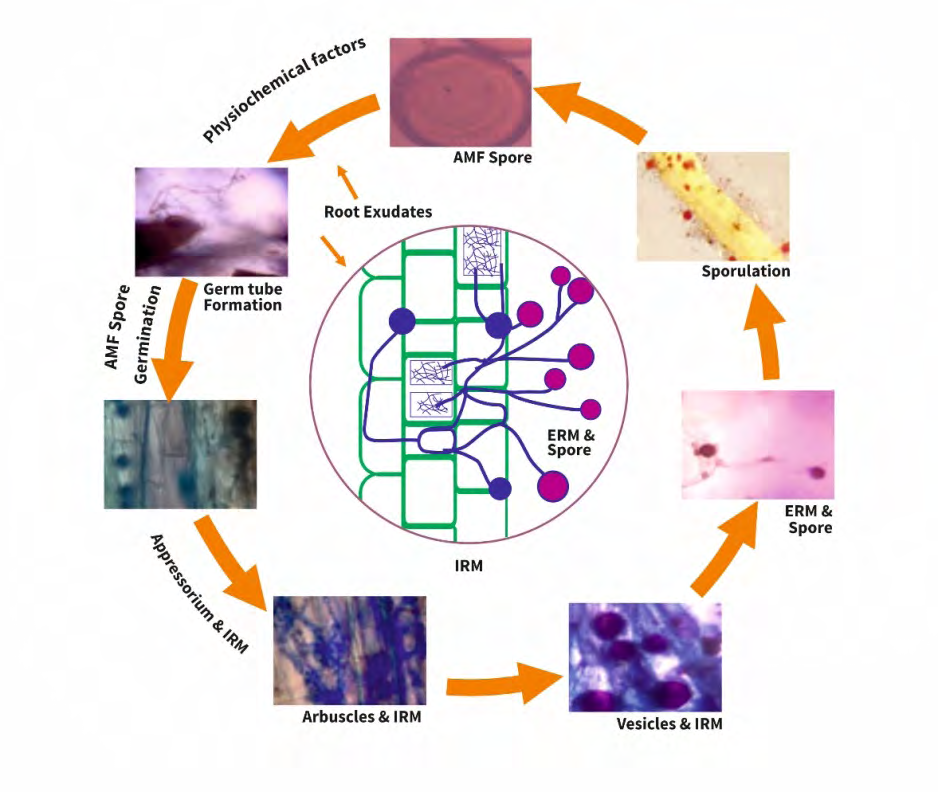
Generally, AM fungal spore reproduction happening with the aid of using asexual spores (Figure 1). AM fungal spores are thick-walled, multinucleate resting structures especially produced on the tip of sporogenous hyphae continuous with mycorrhizal ERH. The germination of the spore does not depend on the plant, as spores have been germinated under experimental conditions in the absence of plants both in vitro (modified living roots) and in vivo condition. However, the speed of germination may be hyperbolic wit aid of using host root exudates (Figure 2). AM fungal spores germinate given suitable conditions of the soil matrix, temperature, Co2 concentration, pH, and PC.
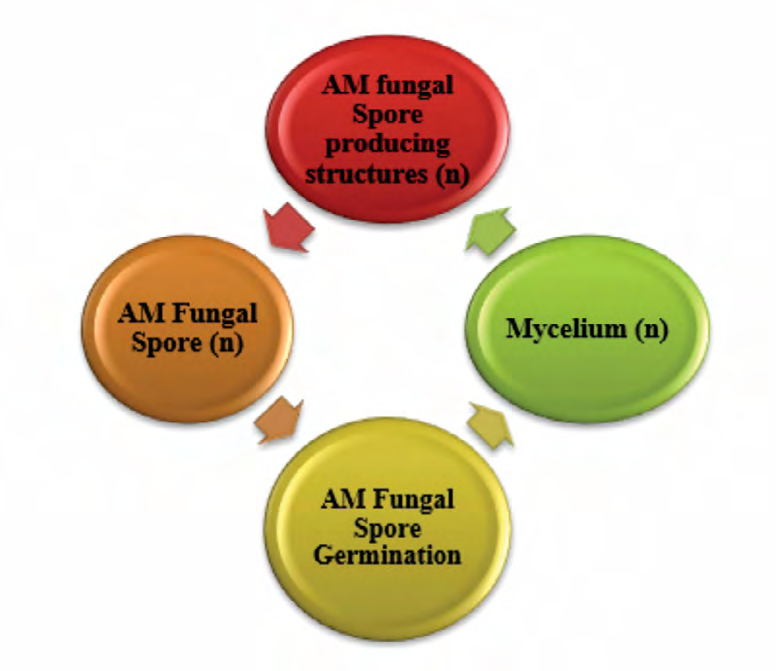
Figure1: Graphical overview of asexual life cycle of AM fungi.
Hyphal growth of AM Fungi
The growth of AM fungal hyphae through the soil is controlled by host root exudates referred to as strigolactones, and consequently the soil PC (Figure 2). Low PC within the soil increase hyphal growth (HG) and branching similarly as induce plant exudation of compounds that manipulate hyphal branching intensity [48, 49]. The branching of AM fungal hyphae grownup in P media of 1 mM is extensively decreased; however, the length of the germ tube and total hyphal growth weren’t affected. A level of 10 mM P pent- up each hyphal growth and branching of AM fungi. This PC occurs in natural soil conditions and might consequently make a contribution to decreased AM fungal colonization [48].
Host Recognition of AM Fungi
Root exudates (RE) from AM fungal host plants grown all through a liquid medium with and without P are proven to have an impact on hyphal growth (Figure 2). Spores of Gigaspora margarita and Glomus intraradices had been grown in host plant exudates. Hyphae of AM fungi grown withinside the exudates from roots starved of P grew plenty of and created tertiary branches as compared to the ones grown in exudates from plants given adequate P. Once the expansion promoting root exudates had been superimposed in low concentration, the AM fungi created scattered long branches. Because the concentration of exudates was enlarged, the fungi created a lot of tightly clustered branches. At the highest concentration arbuscules, the AM fungal structures of P exchange had been formed [48]. Further evidence that AM fungi exhibit host- specific chemotaxis, that allow hyphal growth (HG) closer to the roots of a potential host plant: Spores of Glomus mosseae had been separated from the roots of a host plant, no host plants, and dead host plant through a membrane permeable most effective to hyphae. In the treatment with the host plant, the fungi crossed the membrane and continually emerged within 800 µm of the root, however now no longer within side the remedies with nonhost plants and dead plants [50].
Appressorium/Infection Structure Formation
AM fungal hyphae encounter the root foundation of a host plant, an appressorium or infection structure forms on the root cuticle (Figure 2). From this structure hyphae can penetrate into the host’s parenchyma cortex (Figure 2). AM fungi would really like no chemical signals from the plant to make the appressoria. AM fungi may form appressoria on the cell walls of “ghost” cells in the course of which the protoplast have been eliminated to eliminate signaling among the fungi and additionally the plant host (Figure 2). However, the hyphae didn’t similarly penetrate the cells and develop in close to the root cortex, which suggests that signalling among symbionts is needed for similarly increase growth as soon as appressoria are fashioned [49].
AM Fungal Cell Structure, Metabolism and Lifespans
AM fungi are an obligate biotroph, it requires a living photoautotropic host to complete their lifecycle and produce the next generation of spores (Figures 1 & 2). AM fungal species are totally asexual spore (Figure 1) produced on the pointer of the hyphae. AM fungal spores are producing outside or inside the host root. AM fungal spores are capable to germinate without a host plant in vitro condition with contact of modified living roots. In absence of living root, spores germinate and formed a germination tube which grows via the soil until it finds a host root. AM fungal spores penetrate the root and develop among root cells or it penetrates the cell wall and grown inside root cells (Figure 2). Once spores penetrate the root cells, arbuscules are formed (Figure 2). The arbuscules are tree shaped subcellular for nutrient exchange among AM fungi and symbiotic plant partners. Soil might also additionally include over hundred meters of hyphae per cubic centimetres [24, 51]. This network of hyphae is designed to increase the plants uptake of important macro and micro nutrients which include N, P, K, Zn, Fe, S, Mn, Mg, Cu and water.
Symbiosis of AM fungi
AM fungus forms extremely branched structures in parenchyma for nutrient exchange for the plant referred arbuscules (Figure 2) [1, 4, 6, 52]. These are the distinguishing specific structures of AM fungus. Arbuscules are the sites of exchange for P, C, water, and alternative nutrients [1, 4, 6, 24, 28, 53, 54]. There are two forms: Paris sort is characterised by the expansion of hyphae from one cell to the next; and Arum sort is characterised by the expansion of hyphae within the house between plant cells (55). The selection between Paris sort and Arum sort is primarily determined by the host family, though some families or species are capable of either sort [55]. The host plant exerts an effect over the ERH proliferation and arbuscule formation [1]. There’s a decondensation of the body substance plant’s chromatin that indicates inflated transcription of the plant’s deoxyribonucleic acid (DNA) in arbuscule containing cells [52]. Major modifications are needed within the plant host cell to accommodate the arbuscules. The vacuoles shrink and different cellular organelles proliferate. The plant cell cytoskeleton structure is organized around the arbuscules. There are two different sorts of hyphae that originate from the colonised host plant root. Once colonization has occurred, transient runner hyphae grow from the plant root into the soil. These are the ERH that take up P and other nutrients that are given to the plant. AM fungal hyphae have a high surface-to-volume quantitative relation, creating their assimilative ability larger than that of plant roots [56]. AM fungal hyphae also are finer than roots and may enter into pores of the soil that are inaccessible to roots [1, 57]. The fourth kind of AM fungal hyphae is morphologically distinct and grows from the roots and colonizes different host plant roots [53].
Diversity of AM Fungi
A general 336 species of AM fungi, inside which half dozen dominant genera inclusive of Acaulospora, Glomus, Gigaspora, Scutellospora and Enterophospora are superabundant in cultivated than in non-cultivated lands (AM fungi species listen on Gogle.com). Genus Glomus is dominant genera available in worldwide in land and it’s multiplied for biostimulants.
Characteristics of AM Fungi
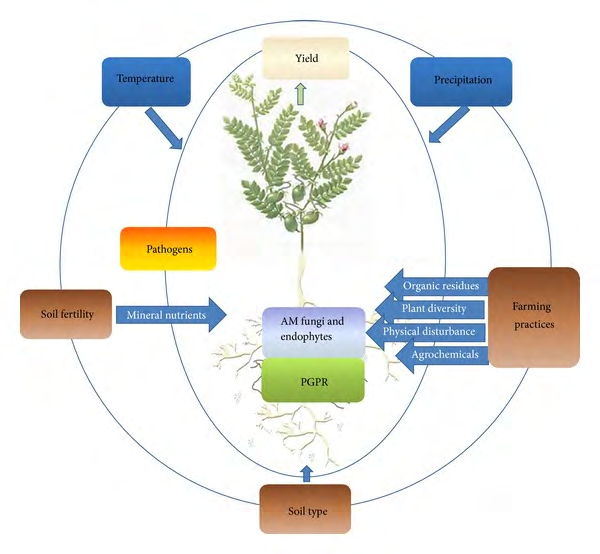
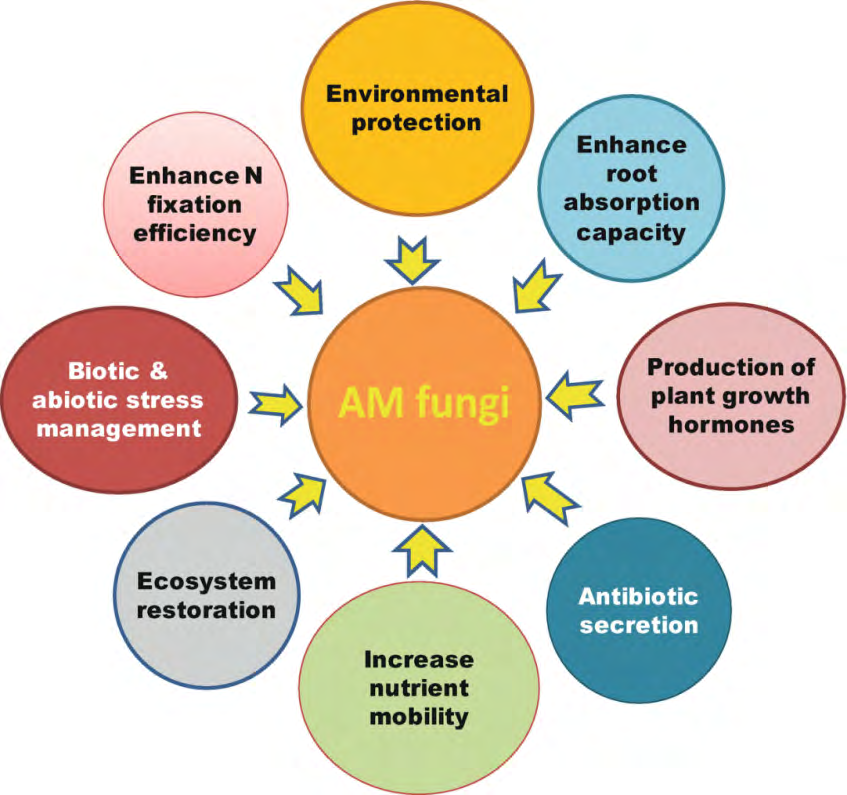
The symbiotic association of AM fungi is a traditional instance of mutualistic relationship that could regulate the growth and improvement of plants (Figures 3,5). The mycelial network of fungi extends underneath the roots of the plant and promotes nutrient uptake (NU) that is otherwise not available. The AM fungal mycelium colonizes roots of many plants belong to different species, resulting into a common mycorrhizal network (CMN). CMN is taken in to the consideration as a primary component of the terrestrial ecosystem (TC) with its extensive consequences on distinct plant communities, specifically on invasive plants [1, 24, 32, 58] and the fungal mediated transport of phosphorus (P) and nitrogen (N) to plants [11, 17, 43, 59, 60]. Moreover, communal nutrients additionally relocate from fungi to the plant, together with different related effects and enhance plant tolerance to biotic and abiotic factors (Figure 3- 5). They have the capacity to enhance characteristics of soil and therefore inspire plant improvement in normal in addition to in stressful circumstances [61, 62, 63]. AM fungi colonization improves tolerance of plants to stressful cues with the aid of using bringing approximately numerous modifications of their morpho-physiological traits [62, 63, 64]. AM fungi are considered as natural growth regulators of a majority of terrestrial flora. AM fungi are used as bioinoculants, and researchers inspire their use as prominent biofertilizers in sustainable crop productivity. Furthermore, AM fungi inoculated soil forms more constant masses and significantly higher extraradical hyphal mycelium [1,6,24,32,65). Glomalin related soil protein (GRSP) is thought to maintain water content in soils exposed to different abiotic stresses [66], which afterward regulates water frequencies among soil and plants, automatically triggering plant improvement. Glomalin contains 30–40% C and its associated compounds that safeguard soil from desiccation with the aid of using improving the soil water holding capacity [67]. Growth associated functions, which includes stomatal conductance, leaf water potential (LWP), relative water content (RWC), PSII efficiency, and CO2 assimilation are affected by AM fungi inoculation [24, 68, 69]. AM fungi additionally help enhance water stress tolerance by physiological alteration of the above ground organs and tissues [70]. AM fungi enhance the accumulation of dry matter and improve water moisture uptake, therefore enhancing plant tolerance against stresses. Exploitation of AM fungi for plant growth in various biological ecosystems can make a contribution significantly to organic culturing for growth promotion and yield maximization.
Environmental Significance of AM Fungi
AM fungi are extraordinary useful to the environment, as they dramatically help in enhancing soil and plant fitness and maximise the absorption of macro and micro nutrients (Figures 3-4). This symbiotic relationship among fungus and plant spanning millions of years and feature allowed for plant to colonise areas that might had been too harsh to withstand. However, their presence in soil makes them liable to erosion, and tilling. Tilling reduces the inoculation of the soil and the efficacy of the fungi by disrupting the hyphal network (Figure 3).
Application of AM Fungi as a Bio-fertilizer
AM fungi produce glomalin protein in soil environment and will increases aggregation of soil particles. It is likewise enhancing soil oxygen and carbon content for plant and soil health (Figure 3). AM fungal mediated plant increased many folds root growth and helps quicker plant establishment. It is increase absorption range through as plenty as 50 times additional and increase overall root biomass and root yield in cereals, pulses, vegetables, spices, fruits crops. Biofertilizers (as AM fungi) are a mixture of naturally occurring substances (spores, root bit, hypha, mycelium and substrate) that are used to improve soil fertility, productivity and they’re their critical significance in agro ecosystems described in figure 5. AM fungi are very beneficial for soil health (SH) in addition to for PG and development. Different research studies carried out on AM fungi during the past three decades have highlighted their uncountable advantages on soil and PH and crop productiveness (CP). Therefore, its miles broadly believed that AM fungi could be considered as a replacement of chemical fertilizers (CF), because of the fact AM fungi utility can effectively reduce the quantitative use of CF input [1, 6, 11, 24]. Continuous use of lifeless CF, herbicides, and fungicides has prompted divers troubles to soil, plants, and human health (HH), through their negative effect on the quality of food products, SH, air and water systems [11, 60, 71]. It is thought that AM fungi can possibly lower down the use of CF up to 50% for best agricultural production; however, this estimate relies upon at the form of plant species and the prevalent stressful regimes.
Nutrient uptake and exchange Efficiency of AM Fungi
AM fungi are favourable mutual symbiotic relationship with host. Bioactive phytochemicals involved in interactions between plants and soil microorganisms along with AM fungi. They have restrained saprobic ability and depend on the host plant for their carbon nutrition for food. AM fungi take up the products of the plant host›s photosynthesis as hexoses. Carbon switch from plant to fungi might also additionally arise through the arbuscules or IRH [1, 4, 5, 6, 32, 72]. Secondary synthesis from the hexoses via way of means of AM fungi takes place withinside the intraradical mycelium (IRM). Inside the mycelium, hexose is transformed to trehalose and glycogen. Trehalose and glycogen are carbon storage forms that can be rapidly synthesized and degraded and might buffer the intracellular sugar concentrations. The intraradical hexose enters the oxidative pentose phosphate pathway, which produces pentose for nucleic acids. Lipid biosynthesis additionally takes place with inside the IRM. Lipids are then stored or exported to ERH wherein they’ll store or metabolized. The breakdown of lipids into hexoses, referred as gluconeogenesis, occurs in the ERM [72]. Approximately
25% of the carbon translocated from the plant to the fungi is stored in the ERH [73]. Nearly 20% of the host plant’s carbon can be transferred to the AM fungi [72]. This represents the host plant’s considerable carbon investment in mycorrhizal network (MNW) and contribution to the below-ground organic carbon pool (OCP).
Increasing the plant’s carbon supply to the AM fungi, AM fungi are escalations uptake and transfer of P and different macro and micro nutrients from to host plant. Likewise, nutrients uptake and transfer are lowered when the photosynthate supplied to the fungi is decreased. Species of AM fungi differ in their capabilities to supply the plant with nutrients. In a few cases, AM fungi are poor symbionts, offering little P even as taking exceedingly excessive quantities of carbon [74]. The foremost gain of AM fungi to plants has been attributed to extended uptake of nutrients, mainly P. AM fungi can be much more efficient than plant roots at taking up P. P and different minerals travels to the root via diffusion and hyphae reduce the distance required for diffusion, thus increasing uptake.
The rate of P flowing into AM fungi can be up to six times that of the root hairs [75]. In few cases, the role of P and different nutrients uptake can be completely taken over by the MNW, and all of the plant’s P can be of hyphal origin [74]. AM fungi are maximum frequent in plants growing on mineral soils, and are of severe significance for plants growing in nutrient-deficient substrates consisting of in volcanic soil and sand dune environments. The populations of AM fungi are best in plant groups with excessive rang consisting of tropical rainforests and temperate grasslands in which they’ve many ability host plants and might take advantage of their ability to colonize a broad host range. There is a decrease incidence of mycorrhizal colonization (MC) in very arid or nutrient-rich soils. Mycorrhizas had been found in aquatic habitats; however, waterlogged soils had been proven to lower MC in a few species [76].
Mineral Nutrition Efficiency of AM Fungi
Excessive land use can also additionally have a drastic effect on the general biodiversity, which in turn can also additionally have an effect on the ecosystem function as publicised through numerous reports. AM fungi enhance water and nutrients uptake because of AM fungal ERH accomplishing out of the depletion region and extract mineral and nutrients are offer to host. AM fungi are plenty beneficial to increase bioavailability of nutrients and allowing for reduced irrigation and higher fertilizer efficiency (Figure 4). An outstanding role of such symbiotic relationship is to transfer nutrients which include organic carbon (OC), with inside the form of lipids and sugars [77]. MC is broadly believed to stimulate nutrient uptake in plants. It is obvious that inoculation of AM fungi can enhance the concentration of numerous macro and micro nutrients significantly, which results in accelerated photosynthate manufacturing and for this reason accelerated biomass accumulation [6, 11, 18, 59, 78]. AM fungi have the functionality to reinforce the uptake of inorganic nutrients in nearly all plants, specifically of P [1, 11, 18, 59]. AM fungi also are very powerful in assisting plants to take up nutrients from the nutrient-deficient soils. Apart from the macronutrients, AM fungi affiliation has been reported to increase the phyto availability of micronutrients such as Zn, Fe and Cu [6, 24]. AM fungi enhance the surface absorbing functionality of host roots. Experimental trials on tomato plants inoculated with AM fungi have proven accelerated leaf area, and N, K, calcium, and P contents, reflecting superior plant growth [79]. AM fungi develop symbiosis with roots to acquire crucial nutrients from the host plant and therefore offer mineral nutrients which include N, P, K, Ca, Zn, and S in return. Thus, AM fungi offer nutritional support to the plants even beneath irrelevant situations in the root cells. AM fungi produce arbuscules fungal structures (AFS), which help in exchange of inorganic minerals and the compounds of C and P, ultimately imparting a considerable vigor to host plants [4, 8, 79, 80, 81, 82]. Therefore, they are able to substantially enhance the P concentration in each root and shoot systems. Under P restricted situation, AM fungal affiliation improves P supply to the infected roots of host plants. Increased photosynthetic activities and different leaf functions are directly related to improved growth frequency of AM fungi inoculation that is directly linked to the uptake of N, P, and C, which move towards roots and promote the improvement of crops. It has been determined that AM fungi maintain P and N uptake ultimately helping in plant improvement at better and decrease P ranges.
AM fungal symbiosis undoubtedly accelerated the concentrations of N, P, and Fe in Pelargonium graveolens L. under drought stress [85]. Gomez-Bellot et al. [86] stated improved ranges of P, Ca, and K in Euonymus japonica under salinity stress because of immediate fungus attachment. It is thought that AM fungi enhance the uptake of almost all essential nutrients and contrarily decrease the uptake of Na and Cl, leading to growth stimulation [87, 88]. Many scientists have stated the role of AM fungi in uptake of soil nutrients, specifically of N and P that could efficiently promote the growth of host plants. Several research studies have defined that AM fungi have the capacity to absorb and transfer N to the close by plants or host plants. AM fungi symbiosis produce tremendous underground ERM starting from the roots as much as the encircling rhizosphere, thereby assisting in enhancing the uptake of nutrients. AM fungi immunization improves C and N accumulation and N assimilation under ambient and elevated CO2 concentrations [89]. AM fungi had been stated to enhance growth, accumulation of micro and macro-nutrients, and their allocation with inside the plantlets grown under accelerated increased ranges of Mn [11, 79, 90]. Enhancement of plant nutrition and upkeep of Ca2+ and Na+ ratio are the substantial dynamic attributes that assist enhance useful components of AM fungi colonization on universal plant performance. Improved growth and levels of protein, Fe, and Zn had been discovered in mycorrhizal chickpea [91]. Moreover, different reports have proven superior activity of a K+ transporter withinside the mycorrhizal roots of Lotus japonicus [92]. Moreover, meta- analysis reports confirmed the role of mycorrhizal symbiosis to numerous micro-nutrients in crops. Several researches carried out at same stage in the previous couple of years have proven that AM fungi (Glomus mosseae and Rhizophagus irregularis) exhibited improved heavy metal translocation with inside the shoot [93]. Micronutrients (Zn and Cu) being diffusion restricted in soils are absorbed through plants with the help of AM fungal hyphae.
Plant Productivity Efficacy of AM fungi
AM fungi now no longer most effective can enhance the nutrient status of crops, however can also enhance the quality and quantity of crops. AM fungi colonized strawberry exhibited increased levels of secondary metabolites resulting in improved antioxidant property [94]. AM fungi can enhance the dietary quality of crops by affecting and manufacturing of carotenoids and certain volatile compounds [95]. Prasad [79] located useful consequences of AM fungi on the quality and productiveness of solanaceous crops (tomatoes, potato, and brinjal). Zeng et al. [96] have mentioned improved contents of sugars, organic acids, vitamin C, flavonoids, and minerals because of Glomus versiforme resulting in enhanced citrus fruit quality. AM fungal symbiosis induces more suitable accumulation of anthocyanins, chlorophyll, carotenoids, overall soluble phenolics, tocopherols, and numerous mineral nutrients [97, 98, 99, 100, 101, 102]. AM fungi had been employed in a large-scale field production of maize, [103], yam [104], potato [105], soybean [102, 106, 107, 108] Allium [11] confirming that AM fungi own a substantial ability for boosting crop yield. AM fungi also can improve the biosynthesis of valuable phytochemicals in edible plants and lead them to healthy food manufacturing chain [109].
AM Fungi Increase Production of Growth Hormones
Plants with AM fungi exhibit better content of growth regulators such as cytokinin’s and auxins as comparison to non-mycorrhizal ones. AM fungi colonised roots display adjustments in root morphology through acquiring plentiful thicker and deliver fewer root hairs. Hormone’s accumulation in the host tissue is affected by MC with adjustments in the levels of cytokinin’s, abscisic acid, gibberellins like substances. The effect of AM fungi on photosynthesis and host morphology might also be hormonal.
Abiotic Stresses efficacy of AM Fungi
Drought tolerance Activity of AM Fungi
AM fungi inoculated plants are successful to tolerance of drought; heat and toxic level of minerals because of the fact those fungi useful for take-up toxic minerals and enhance general plant and soil health [6, 11, 24]. AM fungi assist plants to uptake soil nutrients in exchange for sugars produced by the plants. In forests, ectomycorrhizas form long strands known as hyphae that run among trees, acting as connectors. This massive underground transportation network is known as the CMN. The CMN makes use of chemical communication to exchange nutrients among trees on an as need basis. Besides nutrients, the CMN additionally facilitate trees get water that their personal roots might now no longer be capable of reach. Once deep soils are wet and shallow soils are dry, trees pull water upward through their tap roots, which might be deep withinside the soil, as much as the shallow roots. Evaporation is through the leaves and from the soil surface act like a drinking straw, pulling water through the plant and soil. AM fungi can clutch that water coming up from the deep tap root of one tree, transfer it alongside the highways of the CMN, and deposit it in the roots of a distant tree. AM fungi treated plants are increased tolerance to drought and survival in excessive soil temperature, soil toxins and extreme soil pH.
Salinity tolerance Efficacy of AM Fungi
It is widely recognised that the soil salinization is an increasing environmental hassle posing an excessive risk to global food security. Salinity stress is thought to suppress growth of plants with the aid of using affecting the vegetative improvement and net assimilation rate resulting in decreased yield. It is likewise promoting the immoderate era of reactive oxygen species [110, 111]. Salt affected soils, really appropriate use of AM fungi for mitigating the salinity caused detrimental results on plants [112]. Several research studies have stated the performance of AM fungi to impart growth and yield enhancement in plants under salinity stress [113]. El-Nashar [114] reported that AM fungi enhanced growth rate, leaf water potential (LWP), and water use efficiency (WUE) of the Antirrhinum majus plants. Ait-El- Mokhtar et al. [115] have reported the useful effects of AM fungi symbiosis on physiological parameters together with photosynthetic rate, stomatal conductance, and leaf water relations under saline regimes. AM fungi inoculated Allium sativum plants confirmed improved growth which including leaf area index, fresh and dry biomass under saline conditions [11, 116]. Concentrations of total P, Ca2+, N, Mg2+, and K+ have been better withinside the AM fungal treated Cucumis sativus plants as compared with uninoculated plants under salt stress conditions [117]. AM fungal inoculation to Capsicum annuum exhibited improved chlorophyll contents, and Mg2+ and N uptake coupled with decreased Na+ transport under saline conditions [118]. AM fungal inoculation can effectively regulate the levels of key growth regulators. AM fungi colonized plants have the capability to decrease oxidative stress by suppressing lipid membrane peroxidation under salinity stress [113, 119]. AM fungi have been additionally to improve the accumulation of numerous organic acids ensuring in up regulation of the osmoregulation process in plants grown under saline stress.
Heavy Metals Tolerance Efficacy of AM Fungi
AM fungi are extensively believed to support plant establishment in soils contaminated with heavy metals, due to their potential to reinforce protection system of the AM fungal mediated plants to promote growth and expansion. Heavy metals might also additionally acquire in food crops, fruits, vegetables, and soils, inflicting numerous health hazards. AM fungi affiliation with wheat positively improved nutrient uptake under aluminium stress [120]. Heavy metals can be immobilized in the fungal hyphae of internal and external origin [121] which have the capability to fix heavy metals in the cell wall and store them in the vacuole or might also additionally chelate with a few different substances in the cytoplasm [122] and subsequently reduce metal toxicity withinside the plants. The sturdy outcomes of AM fungi on plant improvement and growth under severe stressful situations are most customarily because of the capability of these fungi in improving morphological and physiological processes that boom plant biomass and therefore uptake of essential immovable nutrients such as Cu, Zn, and P and accordingly decreased metal toxicity in the host plants [24, 32, 123, 124]. It is likewise believed that improved growth or chelation in the rhizospheric soil can cause metal dilution in plant tissues [125]. AM fungi reportedly bind Cd and Zn in the cell wall of mantle hyphae and cortical cells, thereby restricting their uptake and ensuing in better growth, yield, and nutrient status [32]. AM fungal mycelia have a high cation exchange potential and absorption of metals [126]. It is pertinent that low Cd mobility and toxicity also can be addressed with AM fungi through increasing soil pH [127], restoring Cd in the ERM [128], and binding Cd to glomalin, a glycoprotein. AM fungi had been very effective in decreasing the levels of Cd in each the vacuoles and cell wall, which added approximately Cd detoxification in rice [129]. Various procedures that occur through the AM fungi are immobilization/restriction of metal compounds, precipitation of polyphosphate granules in the soil, adsorption to fungal cell wall chitin, and heavy metal chelation inside the fungus (Figure 5).
High and Low Temperature Tolerance Efficacy of AM fungi
As soil temperatures increase, plant community reactions might be relying on AM fungal interactions for sustainable yield. Heat stress notably influences PG and development with the resources of the usage of imparting, loss of plant vigor and inhibition of seed germination, retarded growth rate, decreased biomass production, wilting and burning of leaves and reproductive organs, abscission and senescence of leaves, damage further to discoloration of fruit, reduction in yield and cell death and greater appropriate oxidative stress. Generally, AM fungi treated plants spectacle better growth under heat stress. Maya and Matsubara [130] have stated the association of Glomus fasciculatum with PG and development most important to positive changes in growth under the conditions of high temperature. AM fungi can increase plant tolerance to cold stress. Moreover, a majority of information’s state that several plants inoculated with AM fungi at low temperature grow broaden better than without AM fungal inoculated plants [131, 132]. AM fungi support plants in preventing cold stress and eventually improve plant development [133]. AM fungi also can maintain moisture in the host plant increase plant secondary metabolites leading to strengthen plant immune system, and increase protein content for supporting the plants to combat cold stress conditions [134]. Symbiotic AM fungal relationship improves water and plant relationships and increases gas exchange potential and osmotic adjustment [135]. AM fungi increase the synthesis of chlorophyll leading to a noteworthy perfection in the concentrations of numerous metabolites in plants subjected to cold stress conditions [24, 134, 136].
Environments, Human and AM Fungi
The complete ability that AM fungi can play in agriculture, phytoremediation habitat loss and whilst complete to attain more healthier lawns are currently being investigated. The ability for restoring regions affected with industrial processes ought to enhance the effective value of the area by promoting the revegetation of the area and rebuild the soil of the disturbed area. Similarly, the ability to enhance agricultural yields; if now no longer enhance plant survival, through inoculating the soil with AM fungi ought to show to be a beneficial strategy in the future to ensure food security.
Seed Production and Offspring Fertility Efficacy of AM Fungi
AM fungi are improved vigor in offspring generations and greater fertility and survival of seeds from AM fungal mediated plants
AM Fungi Increased Tolerances to Disease and Pest
AM fungi increase tolerance pests to root and soilborne pathogens and increase tolerance to foliar pathogens via Mycorrhiza-induced resistance in addition to escalation secondary metabolite production [1, 27, 29].
Carbon Cycling Efficacy of AM Fungi
Significant quantity of carbon flows via AM fungal mycelia to absolutely extraordinary additives of soils. Production of glycoproteins inclusive of glomalin which might be involved inside the formation and strength of soil aggregates ought to have additionally an essential influence on extraordinary microorganisms associated with the AM fungal mycelium [23, 54, 137].
Phytoremediation Efficacy of AM Fungi
Disturbance of native plant groups in desertification- threatened regions is frequently accompanied through degradation of physical and biological soil properties, soil structure, nutrient availability, and organic matter. When restoring disturbed land, it is crucial to replace not only the above ground vegetation but also biological and physical soil properties [138]. A particularly new technique to restoring land is to inoculate soil with AM fungi whilst reintroducing vegetation in ecological restoration. It has enabled host plants to establish themselves on degraded soil and enhance soil quality and health [24, 32, 139]. Soil’s quality parameters had been considerably improved long-term when a mixture of indigenous AM fungal species was introduced compared to non-nodulated soil and soil inoculated with a single exotic species of AM fungi [138]. The benefits had been elevated PG, enhanced P uptake [140] and soil N content, higher soil organic matter, soil aggregation, attributed to higher legume nodulation in the presence of AM fungi, higher water infiltration, and soil aeration because of soil aggregation [24, 138]. Native strains of AM fungi improve the extraction of heavy metal(s) from the polluted soils and make the soil healthy and appropriate for crop production [141].
Inferior World without AM Fungi
The vital and useful symbiotic relationship among plants and AM fungi cannot be taken for granted. Life absent AM fungi might be extensively different, because the beneficial relationship among AM fungi and plant are of utter significance for their survival. Their role in the ecosystem in addition to agriculture might be impossible to replace.
Extortions to AM Fungi
Climate change poses a great threat to AM fungi, because of irreparable damage to diverse ecosystems, in addition to the increasing rate of habitat loss because of human activities. Therefore, successive steps must be carried out to mitigate next screw ups which could stand up from those concerns.
Global Climate Change Efficacy of AM fungi
Global climate change is affecting AM fungal populations and interactions among AM fungi and their plant hosts. It is usually accepted that interactions among organisms can affect their response to global climate change. In latest meta- analyses, AM fungi have been discovered to improve plant biomass under drought conditions and reduce plant biomass under simulated nitrogen deposition studies [24, 142, 143]. AM fungi themselves had been proven to increase their biomass in response to accelerated atmospheric CO2 [144]. Climate change is offering us with demanding situations associated with water, food, and nutrient availability. Some places might also additionally have shortages, even as others have excess. The relationship among forest trees and the AM fungi that assist them percentage sources primarily based totally on need would possibly assist us solve those problems.
Insects/pests Resistance Efficiency of AM fungi
AM fungi can usage these underground connections to produce and receive warning signals [145] specifically, once a host plant is attacked by an aphid, the plant signals close connected plants of its condition. The
host plant releases volatile organic compounds that attract the insect’s predators. The plants connected by AM fungi also are prompted to provide identical volatile organic compounds that defense the uninfected plants from being targeted by the insect pest [145]. In addition, this assists the AM fungi by preventing the plant’s carbon relocation that negatively affects the fungi’s growth and happens once the plant is attacked by herbivores [145].
Conclusion
Various research studies have already recognized the useful role of AM fungi in enhancing PG below stressful environments. Therefore, on this manuscript, the existing evidence associated with the AM fungal lifecycle and its widespread position has been combined in a coherent manner for know-how of AM fungal symbiotic courting with a variety of plants below stress environments. Previously, the AM fungi had been particulars mentioned as useful entities for nutrient uptake from soil; however, recently, it’s been honestly depicted that plants inoculated with AM fungi can efficaciously fight divers environmental cues, such as salinity, drought, nutrient stress, alkali stress, cold stress, and excessive temperatures, and therefore enables boosting yield and productiveness of a massive wide variety of agricultural crops, pulses, oilseeds, fruit, fibre crops, vegetables, forest in addition to nursery development. Encouragement of AM fungi utilization is of enormous significance for modern global agricultural systems for their consistent sustainability. Undoubtedly, exploitation of AM fungi for agricultural development can substantially lessen the usage of artificial (lifeless) fertilizers and different chemicals, thereby endorsing the bio healthy agriculture. AM fungi inoculation, growth and productiveness enhancement in crop plants might be useful to triumph over the consumption requirement of increasing population throughout the globe. In addition, environment friendly technologies shall be exceptionally advocated because of their widespread use. The number one cognizance of destiny research studies must be at the identity of genes and gene products controlling the AM fungi mediated growth and improvement regulation below stressful cues. Understanding the AM fungi brought about modulations in the tolerance mechanisms and additionally the crosstalk triggered to control plant overall performance can facilitate enhance crop productivity. AM fungi must be explored in any respect ranges to extra inspect their role in nature as a biofertilizer for sustainable agricultural production for green technology.
Conflict of Interest Statements
There is no conflict of interest.
References
-
Prasad K (1993) Studies on ecological factors affecting vesicular arbuscular mycorrhizal infection in Sugarcane. Muzaffarpur, Bihar, India: B.R. Ambedkar Bihar University, India.
-
Smith SE, Read DJ (1997) Mycorrhizal symbiosis. Academic Press. Harcourt Brace and Company, Publishers. San Diego, London, New York, Boston, Sydney, Tokyo, Toronto.
-
Prasad K (2000) Occurrence of vesicular arbuscular mycorrhizal fungi in some cultivated crop plants. In: Maheshwari DK, Dubey RC, Prasad G, Navneet (Eds.), Microbes: Agriculture, Industry and Environment, Bishen Singh Mahendra Pal Singh Publishers, Dehra Dun, pp: 65- 69.
-
Prasad K (2015) Biofertilizers: A new dimension for agriculture and environmental development to improve production in sustainable manner. Journal of Basic and Applied Mycology 11(2): 5-13.
-
Prasad K (2013) Arbuscular mycorrhizal fungus plays a major role in agriculture and natural ecosystems to improve production in sustainable manner. In: Jamaluddin, Singh AK (Eds.), Microbes and Sustainable Plant Productivity. Scientific Publication (India) Jodhpur; India, pp: 13-138.
-
Prasad K (2017) Biology, Diversity and Promising Role of Mycorrhizal Entophytes for Green Technology. In: Maheshwari DK (Ed.), Endophytes: Biology and Biotechnology, Series Sustainable Development and Biodiversity 15. Springer International Publishing AG, Switzerland, pp: 257-301.
-
Bisleski RL (1973) Phosphate pools, phosphate transport, and phosphate availability. Annual Review of Plant Physiology 24: 225-252.
-
Prasad K, Kaushik S (2004) Ecology, Physiology, Biochemistry and Taxonomy of Arbuscular Mycorrhizal fungi. In: Gautam SP, Bansal YK, Pandey AK (Eds.), Biological Diversity: Current Trends, Prof. R.C. Rajak festschrift volume. Shree Publication and Distributors, New Delhi, India, pp: 134-141.
-
Prasad K, Meghavanshi MK (2005) Interaction between indigenous Glomus fasciculatum (AM fungus) and Rhizobium and their stimulatory effect on growth, nutrient uptake and nodulation in Acacia nilotica (L.) Del. Flora and Fauna 11(1): 51-56.
-
Prasad K, Pandey AK (2012) Mycorrhizal symbiosis: A new dimension for agriculture and environmental development to improve production in sustainable manner In: Bagyaraj DJ, Tilak KVBR, Kehri HK (Eds.), Microbial Diversity and Function, New Delhi, New India Publishing Agency, New Delhi, India, pp: 389-402.
-
Prasad K (2021) Impact of Biological Fertilizer Arbuscular Mycorrhizal Fungi and Conventional Fertilizers Mobilization on Growth, Yield, Nutrient’s uptake, Quercetin and Allin Contents in Allium Crops Cultivation under Field Conditions in Semi-Arid Region of India. South Asia Journal of Experimental Biology 11(1): 15-26.
-
Janos DP (1980) Mycorrhizae influence tropical succession. Biotropica 12(2): 56-64.
-
Allen EB, Allen MF (1986) Water relations of xeric grasses in the field: interactions of mycorrhizae and competition. New Phytologist 104(4): 559-571.
-
Fitter AH (1977) Influence of mycorrhizal infection on competition for phosphorus and potassium by two grasses. New Phytologist 79(1): 119-125.
-
Newman EI (1988) Mycorrhizal links between plants: their functioning and ecological significance. Advances in Ecological Research 18: 243-270.
-
Koske RE, Sutton JC, Sheppard BR (1975) Ecology of Endogone in Lake Huron sand dunes. Canadian Journal of Botany 53(2): 87-93.
-
Prasad K, Deploey JJ (1999) Incidence of arbuscular mycorrhizae and their effect on certain species of trees. Journal of the Pennsylvania Academy of Science 73(3): 117-122.
-
Prasad K (2021) Diversification of Glomermycota form arbuscular mycorrhizal fungi associated with vegetable crops cultivated underneath natural ecosystems in arid region of Rajasthan, India. Current Investigations in Agriculture and Current Research 9(2): 1205-1212.
-
Sutton JC, Sheppard BR (1976) Aggregation of sand- dune soil by endomycorrhizal fungi. Canadian Journal of Botany 54(4): 326-333.
-
Dehn B, Schuepp H (1989) Influence of VA mycorrhizae on the uptake and distribution of heavy metals in plants. Agriculture and Environment 29: 79-83.
-
Griffioen WAJ, Ernst WHO (1989) The role of VA mycorrhiza in the heavy metal tolerance of Agrostis capillaris L. Agriculture, Ecosystems and Environment 29(4): 173-177.
-
Prasad K, Rajak RC (2000) Biotechnological application of mycorrhizae in reclamation of mined dumps. In: Bagyaraj DJ, Tilak KVBR, Kehri HK, (Eds.), Integrated Management of Plant Resources. Scientific Publishers (India): Jodhpur, pp: 283-292.
-
Prasad K, Rajak RC (2001) Microbes and wasteland management: Challenge ahead. In: Roy AK, Varma SK (Eds.), Wasteland Management and Environment. Scientific Publishers (India): Jodhpur, pp: 27-39.
-
Prasad K (2021) Arbuscular mycorrhizal fungi and plant collaborations influences ecology and environmental changes for global sustainable development. Journal of Ecology and Natural Resources 5(1): 1-16.
-
Stahl PO, Smith WK (1984) Effects of different geographic isolates of Glomus on the water relations of Agropyron smithii. Mycologia 76(2): 261-267.
-
Schonbeck F (1978) Einfluss der endotrophen Mykorrhiza auf die Krankheitsresistenz hoherer Pflanzen. Z. PflKrank. PflSchut 85: 191-196.
-
Prasad K (1998) Biological control of rhizospheric microflora of Saccharum officinarum L. plant through vesicular arbuscular mycorrhizal (Glomus fasciculatum) fungi. Biome 8(1-2): 131-136.
-
Prasad K, Warke RV, Khadke K (2019) Management of soilborne pathogens to improve production of pulses using organic Technologies for sustainable agriculture. International Journal of Research and Analytical Reviews 6(2): 82-101.
-
Prasad K (2011) Interaction between arbuscular Mycorrhizal fungus (Glomus fasciculatum) and rhizospheric fungi in Saccharum officinarum L. Amravati University Research Journal 5: 53-65.
-
Graham JH (2000) Assessing costs of arbuscular mycorrhizal symbiosis in agro ecosystems. In: Podila GK, Douds DD Jr (Eds.), Current Advances in Mycorrhizal Research. St. Paul, MN: APS Press, pp: 127-140.
-
Jacobsen I, Rosendahl L (1990) Carbon flow into soil and external hyphae from roots of mycorrhizal cucumber plants. New Phytologist 115(1): 77-83.
-
Prasad K (2020) Positive Importance of Arbuscular Mycorrhizal Fungi for Global Sustainable Agriculture and Environment Management for Green Technology. Current Investigations in Agriculture and Current Research 9(2): 1182-1185.
-
Mosse B (1973) Plant growth response to vesicular- arbuscular mycorrhizae. X. response of Stylosanthes and maize to inoculation in unsterile soils. New Phytologist 78(2): 277-288.
-
Morton JB (1993) Problems and solutions for the integration of glomalin taxonomy, systematic biology, and the study of endomycorrhizal phenomena. Mycorrhiza 2: 97-109.
-
Morton JB (2000) Evolution of endophytism in arbuscular mycorrhizal fungi of Glomales. In: Bacon CW, White JH (Eds.), Microbial endophytes. Marcel Dekker Inc, New York, pp: 121-140.
-
Mosse B (1981) Vesicular-Arbuscular Mycorrhiza Research for Tropical Agriculture. Hawaii Institute of Tropical Agriculture and Human resources, University of Hawaii.
-
Morton JB, Redecker D (2001) Two families of Glomales, Archaeosporaceae and Paraglomaceae, with two new genera Archaeospora and Paraglomus, based on concordant molecular and morphological characters. Mycologia 93(1): 181-195.
-
Sieverding E, Oehl F (2006) Revision of Entrophospora and description of Kuklospora and Intraspora, two new genera in the arbuscular mycorrhizal Glomeromycetes. Journal of Applied Botany and Food Quality 80(1): 69- 81.
-
Spatafora JW, Chang Y, Benny GL, Lazarus K, Smith ME, et al. (2016) A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 108(5): 1028-1046.
-
Redecker D, Schussler A, Stockinger H, Sturmer SL, Morton JB, et al. (2013) An evidence-based consensus for the classification of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza 23(7): 515-531.
-
Bago B, Pfeffer PE, Shachar-Hill Y (2000) Carbon metabolism and transport in arbuscular mycorrhizas. Plant Physiology 124(3): 949-958.
-
Jiang YN, Wang WX, Xie QJ, Liu N, Liu LX, et al. (2017) Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 356(6343): 1172-1175.
-
Smith S, Read D (2008) Mycorrhiza symbiosis. 3rd (Edn.), San Diego, CA: Academic Press.
-
Jung SC, Martinez-Medina A, Lopez-Raez JA, Pozo MJ (2012) Mycorrhiza-induced resistance and priming of plant defences. Journal of Chemical Ecology 38: 651-664.
-
Kar RK, Mandaokar BD, Kar R (2005) Mycorrhizal fossil fungi from the Miocene sediments of Mirozam, Northeast India. Current Science 89: 257-259.
-
Remy W, Taylor T, Hass H, Kerp H (1994) Four hundred- million-year-old vesicular arbuscular mycorrhizae. Proceedings of the National Academy of Sciences of the United States of America 91(25): 11841-11843.
-
Wang B, Yeun LH, Xue Y, Liu Y, Ane JM, et al. (2010) Presence of three mycorrhizal genes in the common ancestor of land plants suggests a key role of mycorrhizas in the colonization of land by plants. New Phytologist 186(2): 514-525.
-
Nagahashi G, Douds DD, Abney GD (1996) Phosphorus amendment inhibits hyphal branching of VAM fungus Gigaspora margarita directly and indirectly through its effect on root exudation. Mycorrhiza 6(5): 403-408.
-
Douds DD, Nagahashi G (2000) Signalling and Recognition Events Prior to Colonisation of Roots by Arbuscular Mycorrhizal Fungi. In: Podila GK, Douds DD (Eds.), Current Advances in Mycorrhizae Research. Minnesota: APS Press, pp: 11-18.
-
Sbrana C, Giovannetti M (2005) Chemotropism in the arbuscular mycorrhizal fungus Glomus mosseae. Mycorrhiza 15(7): 539-545.
-
Parniske M (2008) Arbuscular mycorrhiza: the mother of plant root endosymbiosis. Nature Reviews Microbiology 6(10): 763-775.
-
Gianinazzi S, Gianinazzi-Pearson V (1986) Progress and headaches in endomycorrhiza biotechnology. Symbiosis 2: 139-149.
-
Wright SF (2005) Management of Arbuscular Mycorrhizal Fungi. In: Zobel RW, Wright SF (Eds.), Roots and Soil Management: Interactions between roots and the soil. USA: American Society of Agronomy, pp: 183- 197.
-
Prasad K, Warke RV (2018) Biology, diversity and promising role of microbial inoculations specially mycorrhizae in agriculture and natural ecosystems to improve productivity for sustainable development and environmental sustainability, National conference, GKV, Hardwar, India, pp: 7-8.
-
Armstrong L, Peterson RL, Armstrong L, Peterson RL (2002) The Interface between the Arbuscular Mycorrhizal Fungus Glomus intraradices and Root Cells of Panax quinquefolius: A Paris-Type Mycorrhizal Association. Mycologia 94(4): 587-595.
-
Turrini A, Bedini A, Loor MB, Santini G, Sbrana C, et al. (2018) Local diversity of native arbuscular mycorrhizal symbionts differentially affects growth and nutrition of three crop plant species. Biology and Fertility of Soils 54: 203-217.
-
Bolan NS (1991) A critical review of the role of mycorrhizal fungi in the uptake of phosphorus by plants. Plant and Soil 134(2): 189-207.
-
Pringle A, Bever JD, Gardes M, Parrent JL, Rillig MC, Klironomos JN (2009) Mycorrhizal symbioses and plant invasions. Annual Review of Ecology, Evolution, and Systematics 40: 699-715.
-
Prasad K (2021) Effect of Dual Inoculation of Arbuscular Mycorrhiza Fungus and Cultivar Specific Bradyrhizobium Japonnicum on the Growth, Yield, Chlorophyll, Nitrogen and Phosphorus Contents of Soybean (Glycine Max (L.) Merrill.) grown on Alluvial Soil. Journal of Innovation in Applied Research 4(1): 1-12.
-
Prasad K (2021) Advantages and nutritional importance of organic agriculture produces food on human, soil and environmental health in modern lifestyle for sustainable development. Aditum Journal of Clinical and Biomedical Research 5(2): 1-7.
-
Navarro JM, Perez-Tornero O, Morte A (2014) Alleviation of salt stress in citrus seedlings inoculated with arbuscular mycorrhizal fungi depends on the root stock salt tolerance. Journal of Plant Physiology 171(1): 76-85.
-
Alqarawi AA, Abd-Allah EF, Hashem A (2014) Alleviation of salt-induced adverse impact via mycorrhizal fungi in Ephedra aphylla Forssk. Journal of Plant Interactions 9(1): 802-810.
-
Alqarawi AA, Hashem A, Abd_Allah EF, Alshahrani TS, Huqail AA (2014) Effect of salinity on moisture content, pigment system, and lipid composition in Ephedra alata Decne. Acta biologica Hungarica 65 (1): 61-71.
-
Hashem A, Abd_Allah EF, Alqarawi AA, Aldubise A, Egamberdieva D (2015) Arbuscular mycorrhizal fungi enhance salinity tolerance of Panicum turgidum Forssk by altering photosynthetic and antioxidant pathways. Journal of Plant Interactions 10(1): 230-242.
-
Syamsiyah J, Herawati A, Mujiyo (2018) The potential of arbuscular mycorrhizal fungi application on aggregate stability in Alfisol soil. IOP Conf. Series: Earth Environmental Science 142: 012045.
-
Wu Z, McGrouther K, Huang J, Wu P, Wu W, et al. (2014) Decomposition and the contribution of glomalin-related soil protein (GRSP) in heavy metal sequestration: field experiment. Soil Biology and Biochemistry 68: 283-290.
-
Sharma S, Prasad R, Varma A, Sharma AK (2017) Glycoprotein associated with Funneliformis coronatum, Gigaspora margarita and Acaulospora scrobiculata suppress the plant pathogens in vitro. Asian Journal of Plant Pathology 11(4): 192-202.
-
He F, Sheng M, Tang M (2017) Effects of Rhizophagus irregularis on photosynthesis and antioxidative enzymatic system in Robinia pseudoacacia L. under drought Stress. Frontiers in Plant Science 8: 183-186.
-
Chandrasekaran M, Chanratana M, Kim K, Seshadri S, Sa T (2019) Impact of arbuscular mycorrhizal fungi on photosynthesis, water status, and gas exchange of plants under salt stress—a meta-analysis. Frontiers in Plant Science 10: 457-460.
-
Barzana G, Aroca R, Paz JA, Chaumont F, Martinez-Ballesta MC, et al. (2012) Arbuscular mycorrhizal symbiosis increases relative apoplastic water flow in roots of the host plant under both well-watered and drought stress conditions. Annals of Botany 109(5): 1009-1017.
-
Yang S, Li F, Malhi SS, Wang P, Dongrang S, et al. (2004) Long term fertilization effects on crop yield and nitrate nitrogen accumulation in soil in North-western China. Agronomy Journal 96(4): 1039-1049.
-
Pfeffer P, Douds D, Becard G, Shachar-Hill Y (1999) Carbon Uptake and the Metabolism and Transport of Lipids in an Arbuscular Mycorrhiza. Plant Physiology 120(2): 587- 598.
-
Hamel C (2004) Impact of arbuscular mycorrhiza fungi on N and P cycling in the root zone. Canadian Journal of Soil Science 84(4): 383-395.
-
Smith S, Smith A, Jakobsen I (2003) Mycorrhizal Fungi Can Dominate Phosphate Supply to Plants Irrespective of Growth Responses. Plant Physiology 133(1): 16-20.
-
Smith SE, Read DJ (2002) Mycorrhizal Symbiosis. Academic Press: London.
-
Luginbuehl LH, Menard GN, Kurup S, Van Erp H, Radhakrishnan GV, et al. (2017) Fatty acids in arbuscular mycorrhizal fungi are synthesized by the host plant. Science 356(6343): 1175-1178.
-
Mitra D, Navendra U, Panneerselvam U, Ansuman S, Ganeshamurthy AN, et al. (2019) Role of mycorrhiza and its associated bacteria on plant growth promotion and nutrient management in sustainable agriculture. International Journal of Life Sciences and Applied Science 1(1): 1-10.
-
Prasad K (2021) Influence of Arbuscular Mycorrhizal Fungal Biostimulants ad Conventional Fertilizers on Some Solanaceous Crops for Growth, Productivity and Nutrient Stoichiometry Under Field Conditions in Semi- Arid Region of Maharashtra, India. Journal of Experimental Biology and Agricultural Sciences 9(1): 75-86.
-
Li X, Zeng R, Liao H (2016) Improving crop nutrient efficiency through root architecture modifications. Journal of Integrative Plant Biology 58(3): 193-202.
-
Prasad K (1997) Effect of vesicular arbuscular mycorrhizal fungi, particularly Glomus fasciculatum and phosphates on chlorophyll content of Saccharum officinarum L. Indian Journal of Applied and Pure Biology 12(2): 91-95.
-
Prasad K, Gautam SP (2005) Effect of inoculation of arbuscular mycorrhizal fungus (Glomus macrocarpum) on the growth and nutrients uptake of Dendrocalamus strictus (Roxb.) Ness under field conditions. Anusandhan 1: 53-62.
-
Liu LZ, Gong ZQ, Zhang YL, Li PJ (2014) Growth, cadmium uptake and accumulation of maize Zea mays L. under the effects of arbuscular mycorrhizal fungi. Ecotoxicology 23(10): 1979-1986.
-
Liu C, Ravnskov S, Liu F, Rubaek GH, Andersen MN (2018) Arbuscular mycorrhizal fungi alleviate abiotic stresses in potato plants caused by low phosphorus and deficit irrigation/partial root-zone drying. Journal of Agricultural Science 156(1): 46-58.
-
Amiri R, Ali N, Nematollah E, Mohammad RS (2017) Nutritional status, essential oil changes and water-use efficiency of rose geranium in response to arbuscular mycorrhizal fungi and water deficiency stress. Symbiosis 73: 15-25.
-
Gomez-Bellot MJ, Ortuno MF, Nortes PA, Vicente-Sanchez J, Banon S, et al. (2015) Mycorrhizal euonymus plants and reclaimed water: biomass, water status and nutritional responses. Scientia Horticulturae 186: 61-69.
-
Evelin H, Giri B, Kapoor R (2012) Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-raecum. Mycorrhiza 22(3): 203-217.
-
Gautam SP, Prasad K (2001) VA mycorrhiza-Importance and biotechnological application. In: Maheshwari DK, Dubey RC, (Eds.), Innovative Approaches in Microbiology. Bishon Singh Mahendra Pal Singh, Dehradun, India, pp: 83-114.
-
Zhu XC, Song FB, Liu SQ, Liu FL (2016) Arbuscular mycorrhiza improves growth, nitrogen uptake, and nitrogen use efficiency in wheat grown under elevated CO2. Mycorrhiza 26(2): 133-140.
-
Bati CB, Santilli E, Lombardo L (2015) Effect of arbuscular mycorrhizal fungi on growth and on micronutrient and macronutrient uptake and allocation in olive plantlets growing under high total Mn levels. Mycorrhiza 25(2): 97-108.
-
Pellegrino E, Bedini S (2014) Enhancing ecosystem services in sustainable agriculture: Biofertilization and biofortification of chickpea (Cicer arietinum L.) by arbuscular mycorrhizal fungi. Soil Biology and Biochemistry 68: 429-439.
-
Battini F, Grønlund M, Agnolucci M, Giovannetti M, Jakobsen I (2017) Facilitation of phosphorus uptake in maize plants by mycorrhizosphere bacteria. Scientific Reports 7: 4686-4689.
-
Ali N, Masood S, Mukhtar T, Kamran MA, Rafique M, et al. (2015) Differential effects of cadmium and chromium on growth, photosynthetic activity, and metal uptake of Linum usitatissimum in association with Glomus intraradices. Environmental Monitoring and Assessment 187(6): 311-314.
-
Castellanos-Morales V, Villegas J, Wendelin S, Vierheiling H, Eder R, et al. (2010) Root colonization by the arbuscular mycorrhizal fungus Glomus intraradices alters the quality of strawberry fruit (Fragaria ananassa Duch.) at different nitrogen levels. Journal of the Science of Food and Agriculture 90(11): 1774-1782.
-
Hart M, Ehret DL, Krumbein A, Leung C, Murch S, et al. (2015) Inoculation with arbuscular mycorrhizal fungi improves the nutritional value of tomatoes. Mycorrhiza 25(5): 359-376.
-
Zeng L, Jian Fu L, Jian Fu L, Ming Yuan W (2014) Effects of arbuscular mycorrhizal (AM) fungi on citrus quality under nature conditions. Southwest China Journal of agricultural science 27(5): 2101-2105.
-
Baslam M, Garmendia I, Goicoechea N (2011) Arbuscular mycorrhizal fungi (AMF) improved growth and nutritional quality of greenhouse grown lettuce. Journal of Agricultural and Food Chemistry 59(10): 5504-5515.
-
Prasad K (1997) Improvement of biomass yields of Terminalia Arjuna L. through vesicular arbuscular mycorrhizal fungi (Glomus fasciculatum) under nursery and field condition. Journal of Phytological Research 10(2): 39-41.
-
Prasad K (1998) Effect of Glomus fasciculatum VAM form and Rhizobium on biomass yield and nutrient uptake of Dalbergia sissoo L. Journal of Tropical Forestry 14(111): 143-148.
-
Prasad K (2002) Interaction between Glomus fasciculatum AMF and Rhizobium and their effect of Prosopis juliflora in nursery conditions. Journal of Basic Applied Mycology 1(1): 130-143.
-
Prasad K (2010) Responses of dual inoculation of arbuscular mycorrhizal fungi on the biomass production, phosphate, roots and shoots phenol concentrations of Terminali arjuna under field conditions. Mycorrhiza News 22(2): 12-27.
-
Prasad K (2011) Effect of Glomus intraradices AM fungi on the shoot dry matter, seed dry yield, and nitrogen and phosphorus uptake of soybean (Glycine max (L.) Merrill) inoculated with cultivar-specific Bradyrhizobium japonicum. Mycorrhiza News 23(3): 13-18.
-
Sabia E, Claps S, Morone G, Bruno A, Sepe L, Aleandri R (2015) Field inoculation of arbuscular mycorrhiza on maize (Zea mays L.) under low inputs: preliminary study on quantitative and qualitative aspects. Italian Journal of Agronomy 10(1): 30-33.
-
Lu F, Lee C, Wang C (2015) The influence of arbuscular mycorrhizal fungi inoculation on yam (Dioscorea spp.) tuber weights and secondary metabolite content. Peer Journal 3: 1266.
-
Hijri M (2016) Analysis of a large dataset form field mycorrhizal inoculation trials on potato showed highly significant increase in yield. Mycorrhiza 26(3): 209-214.
-
Meghavanshi MK, Prasad K, Harwani D, Mahna SK (2008) Response of soybean cultivars towards inoculation with three Arbuscular Mycorrhizal Fungi (AMF) and Bradyrhizobium japonicum in alluvial soil. European Journal of Soil Biology 44(3): 316-323.
-
Meghavanshi MK, Prasad K, Mahna SK (2010) Symbiotic potential, competitiveness and compatibility of indigenous Bradyrhizobium japonicum isolates to three soybean genotypes of two distinct agro-climatic regions of Rajasthan, India. Saudi Journal of Biological Sciences 17(4): 303-310.
-
Meghvanshi MK, Prasad K, Mahna SK (2005) Identification of pH tolerant Bradyrhizobium japonicum strains and their symbiotic effectiveness in soybean (Glycine max (L.) Merr.) in low nutrient soil. African Journal of Biotechnology 4(7): 663-666.
-
Rouphael Y, Franken P, Schneider C, Schwarz D, Giovannetti M, et al. (2015) Arbuscular mycorrhizal fungi act as bio-stimulants in horticultural crops. Scientia Horticulturae 196: 91-108.
-
Ahanger MA, Tomar NS, Tittal M, Argal S, Agarwal RM (2017) Plant growth under water/ salt stress: ROS production; antioxidants and significance of added potassium under such conditions. Physiology and Molecular Biology of Plants 23(4): 731-744.
-
Ahanger MA, Alyemeni MN, Wijaya L, Alamri SA, Alam P, et al. (2018) Potential of exogenously sourced kinetin in protecting Solanum lycopersicum from NaCl- induced oxidative stress through up-regulation of the antioxidant system, ascorbate–glutathione cycle and glyoxalase system. PLoS One 13 (9): e0202-e0175.
-
Santander C, Sanhueza M, Olave J, Borie F, Valentine C, et al. (2019) Arbuscular mycorrhizal colonization promotes the tolerance to salt stress in lettuce plants through an efficient modification of ionic balance. Journal of Soil Science and Plant Nutrition 19(2): 321-331.
-
Latef AAHA, Chaoxing H (2014) Does Inoculation with Glomus mosseae Improve Salt Tolerance in Pepper Plants? Journal of Plant Growth Regulation 33: 644-653.
-
Nashar YIE (2017) Response of snapdragon (Antirrhinum majus L.) to blended water irrigation and arbuscular mycorrhizal fungi inoculation: uptake of minerals and leaf water relations. Photosynthetica 55(2): 201-209.
-
Mokhtar MAE, Laouane RB, Anli M, Boutasknit A, Wahbi S, et al. (2019) Use of mycorrhizal fungi in improving tolerance of the date palm (Phoenix dactylifera L.) seedlings to salt stress. Science Horticulturae 253(8): 429-438.
-
Borde M, Dudhane M, Jite PK (2010) AM fungi influences the photosynthetic activity, growth and antioxidant enzymes in Allium sativum L. under salinity condition. Notulae Scientia Biologicae 2(4): 64-71.
-
Hashem A, Alqarawi AA, Radhakrishnan R, Arjani ABFA, Aldehaish HA, et al. (2018) Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi Journal of Biological Sciences 25(6): 1102-1114.
-
Cekic FO, Unyayar S, Ortas I (2012) Effects of arbuscular mycorrhizal inoculation on biochemical parameters in capsicum annuum grown under long term salt stress. Turk J Bot 36: 63-72.
-
Talaat NB, Shawky BT (2014) Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity. Environmental and Experimental Botany 98: 20-31.
-
Aguilera P, Cornejo P, Borie F, Barea JM, Baer EV, et al. (2014) Diversity of arbuscular mycorrhizal fungi associated with Triticum aestivum L. plants growing in an and sol with high aluminum level. Agriculture, Ecosystems and Environment 186: 178-184.
-
Ouziad F, Hildebrandt U, Schmelzer E, Bothe H (2005) Differential gene expressions in arbuscular mycorrhizal-colonized tomato grown under heavy metal stress. J Plant Physiol 162: 634-649.
-
Punamiya P, Datta R, Sarkar D, Barber S, Patel M, et al. (2010) Symbiotic role of Glomus mosseae in phytoextraction of lead in vetiver grass Chrysopogon zizanioides L. J Hazard Mater 177(1-3): 465-474.
-
Kanwal S, Bano A, Malik RN (2015) Effects of arbuscular mycorrhizal fungi on metals uptake, physiological and biochemical response of Medicago sativa L. with increasing Zn and Cd concentrations in soil. American Journal of Plant Sciences 6(18): 2906- 2923.
-
Miransari M (2017) Arbuscular mycorrhizal fungi and heavy metal tolerance in plants. In: Wu QS (Ed.), Arbuscular mycorrhizas and stress tolerance of plants. Singapore: Springer Nature, pp: 174-161.
-
Audet P (2014) Arbuscular mycorrhizal fungi and metal phytoremediation: ecophysiological complementarity in relation to environmental stress. In: Ahmad P, Rasool S (Eds.), Emerging technologies and management of crop stress tolerance. San Diego: Academic Press, pp: 133-160.
-
Takacs T, Voros I (2003) Effect of metal non-adapted arbuscular mycorrhizal fungi on Cd, Ni and Zn uptake by ryegrass. Acta Agronomica Hungarica Hungary 513: 347-354.
-
Shen H, Christie P, Li X (2006) Uptake of zinc, cadmium and phosphorus by arbuscular mycorrhizal maize (Zea mays L.) from a low available phosphorus calcareous soil spiked with zinc and cadmium. Environ Geochem Health 28(1-2): 111-115.
-
Janouskova, M., Pavlikova D (2010) Cadmium immobilization in the rhizosphere of arbuscular mycorrhizal plants by the fungal extraradical mycelium. Plant and Soil 332: 511-520.
-
Li H, Luo N, Zhan LJ, Zhao HM, Li YW, et al. (2016) Do arbuscular mycorrhizal fungi affect cadmium uptake kinetics, subcellular distribution and chemical forms in rice? Sci Total Environ 571: 1183-1190.
-
Maya MA, Matsubara YI (2013) Influence of arbuscular mycorrhiza on the growth and antioxidative activity in Cyclamen under heat stress. Mycorrhiza 23(5): 381-390.
-
Chen S, Jin W, Liu A, Zhang S, Liu D, et al. (2013) Arbuscular mycorrhizal fungi (AMF) increase growth and secondary metabolism in cucumber subjected to low temperature stress. Scientia Horticulturae 160: 222- 229.
-
Liu X, Song Q, Tang Y, Li W, Xu J, et al. (2013) Human health risk assessment of heavy metals in soil-vegetable system: a multi-medium analysis. Sci Total Environ 463- 464: 530-540.
-
Birhane E, Sterck FJ, Fetene M, Bongers F, Kuyper TW (2012) Arbuscular mycorrhizal fungi enhance photosynthesis, water use efficiency, and growth of frankincense seedlings under pulsed water availability conditions. Oecologia 169(4): 895-904.
-
Latef AAHA, Chaoxing H (2011) Arbuscular mycorrhizal influence on growth, photosynthetic pigments, osmotic adjustment and oxidative stress in tomato plants subjected to low temperature stress. Acta physiologiae Plantarum 33: 1217-1225.
-
Zhu XC, Song FB, Liu SQ, Liu TD, Zhou X (2012) Arbuscular mycorrhizae improve photosynthesis and water status of Zea mays L. under drought stress. Plant, Soil and Environment 58: 186-191.
-
Zhu XC, Song FB, Xu HW (2010) Arbuscular mycorrhizae improve low temperature stress in maize via alterations in host water status and photosynthesis. Plant and Soil 331(1-2): 129-137.
-
Ali N, Masood S, Mukhtar T, Kamran MA, Rafique M, et al. (2015) Differential effects of cadmium and chromium on growth, photosynthetic activity, and metal uptake of Linum usitatissimum in association with Glomus intraradices. Environ Monit Assess 187(6): 311.
-
Jeffries P, Gianinazzi S, Perotto S, Turnau K, Barea JM (2003) The Contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biology and Fertility of Soils 37: 1-16.
-
Akhtar O, Mishra R, Kehri HK (2017) Arbuscular Mycorrhizal Association Contributes to Cr Accumulation and Tolerance in Plants Growing on Cr Contaminated Soils. Proceedings of the National Academy of Sciences India. Section B, Biological Sciences 89(1): 63-70.
-
Fillion M, Brisson J, Guidi W, Labrecque M (2011) Increasing phosphorus removal in willow and poplar vegetation filters using arbuscular mycorrhizal fungi. Ecological Engineering 37(2): 199-205.
-
Akhtar O, Kehri HK, Zoomi I (2020) Arbuscular mycorrhiza and Aspergillus terreus inoculation along with compost amendment enhance the phytoremediation of Cr-rich technosol by Solanum lycopersicum under field conditions. Ecotoxicology and Environmental Safety 201: 110869.
-
Kivlin SN, Emery SM, Rudgers JA (2013) Fungal symbionts alter plant response to global change. American AM J Bot 100(7): 1445-1457.
-
Worchel ER, Giauque HE, Kivlin SN (2013) Fungal symbionts alter plant drought response. Microb Ecol 65(3): 671-678.
-
Treseder KK (2004) A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytologist 164(2): 347-355.
-
Babikov Z, Lucy G, Bruce Toby JA, Birkett M, Caulfield JC, et al. (2013) Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack. Ecology Letters 16(7): 835-843.
-
Johnson D, Gilbert L (2015) Interplant signalling through hyphal networks. New Phytol 205(4): 1448- 1453.
- Lessons to Learn: Trees are More than the Lungs of the World
- Community Forestry Enterprises as a Model for Sustainable Forest Development: The Case Of The "Baja Tarahumara" in Chihuahua, Mexico
- Ecological and Socio-Economic Impacts of Chromolaena odorata and Mesosphaerum suaveolens, Two Invasive Alien Species in Central and Southern Benin, West Africa
- Epigenetic Sustainability: Modeling the Human Factor as a Natural Resource through Science 4.0 and the NR3C1 Biological Pilot
- Growth-at-Risk: A Framework for Assessing Economic Vulnerability
- The Rural Territory as a Socioecological System for the Management of Public Policy for Sustainable Rural Development