Environmental and Ecological Impact of Soil Microorganisms in Plant Sciences
The field of environmental and microbial ecology is undergoing a revolution, having ramifications in microbiology, ecology, and ecosystem research. The tremendous accumulation of molecular data is revealing a vast diversity of microbial groups, as well as unique microbial functions. Plants are part of a varied ecosystem in their natural habitat, which includes numerous and different microorganisms in the soil. Some of these microbes, such as mycorrhizal fungi and nitrogen-fixing symbiotic bacteria, have long been known to improve plant performance through enhancing mineral nutrition. A substantial quantity of data on the structure and dynamics of plant microbiota, as well as the functional capacities of isolated community members, is currently available. The adoption of appropriate delivery techniques and formulations, as well as smart, knowledge-driven microbe selection, is required. Plant-microbe interactions may now be studied in greater depth thanks to modern biotechnology. In this review, we look at the interaction between the soil, the host, and the microbial population, as well as their involvement in plant sciences, to see what we can learn about the ecosystem function of plants and symbiotic microorganisms, which will help us to improve agricultural techniques.
Introduction
Several studies have shown that soil microorganisms and plant–microbe interactions can improve environmental quality and reduce global warming. Soil microbial community composition changes between compartments and levels, posing a serious problem in soil ecology. Microbes, especially plant growth-promoting microorganisms, are vital in agriculture (PGPMs). The following three mechanisms contribute to plant growth - (i) PGPMs that operate as biofertilizers (like nitrogen-fixing bacteria and phosphate- solubilizing bacteria) help plants absorb nutrients [1], (ii) Phytostimulators (microbes that produce phytohormones, including Azospirillum) can directly stimulate plant development [2, 3], (iii) Plants are protected from phytopathogens by biological control agents (Trichoderma, Pseudomonas, and Bacillus) [4–6].
However, as a metaorganism, plants have only lately been identified as having their own specific and intertwined microbiome [7]. A plant’s ecology is influenced by interactions with microbes that live in or on the plant. In ecosystem functioning, the roles of both plants and their accompanying microorganisms have been recognized, but the specific mechanisms are still unclear. Being unable to move, plants have co-evolved with microorganisms, which have resulted in a variety of processes that influence the outcome of their interactions [8]. In Ciccazzo’s work [9], a certain bacterial population is selected by a plant species. The long-term effects of soil microbes on a plant species’ coexistence are defined by changes in the plant’s relative performance in response to changes in the microbial composition [10, 11]. A variety of volatile compounds can be produced by plant growth-promoting rhizobacteria (PGPR), and these compounds are specific to bacterial species and other closely related species [12, 13, 14]. Some of these bacterial volatiles can boost plant development, decrease disease- stimulating induced systemic resistance (ISR), or antagonize phytopathogens [15], nematodes, or insects [16, 17]. In this review, we will explore understanding the environmental and ecological impacts of soil microorganisms which will help farmers improve their methods while also helping us to appreciate the significance of microbial communities in plant biology.
Environmental Impact of Soil Microorganisms
Some soil microorganisms known as mycorrhiza, as well as plant-associated bacteria (rhizospheric or endophytic), can help their plant hosts with nutrient supply, which is vital not only for optimal plant development in normal conditions, but also for plant survival in hostile settings [18]. Endophytic bacteria can support plant growth through a variety of mechanisms, including biological nitrogen fixation, tolerance, and the existence of pollutant degradation pathways. The biomass obtained from those plants and soil microbes can be a valuable resource for producing bio-energy as alternative fuels such as biodiesel, bioethanol, or biogas, which are thought to slow down global warming. As a result, it’s possible that this occurred as part of the compatible reaction during mycorrhization. During times of world change, mycorrhiza may be useful in overcoming xenobiotic or environmental stress [18, 19].
Furthermore, knowing changes in ecosystem environmental characteristics is important on a worldwide basis. On the one hand, more precise understanding would aid in improved environmental management and the protection of environmental structures, reducing damages such as soil contamination and erosion in ecosystems subjected to pollution, drought, or other environmental stressors. Locally, sustainable usage of environmental species is required for a balanced economic and ecological growth, especially in light of global warming. In recent years, subsidies for research in both ecosystems have been substantially decreased, exacerbating the problems of providing incentives to conserve environmental quality and assure functional microbial behaviour [18, 19]. With such a powerful impact on the environment, it is critical to better understand the mechanisms involved in order to control it. The impact of environmental management on soil microorganisms is critical for improving our understanding of how to maintain such ecosystems’ quality. Moreover, soil structure is a part of the environment’s ecology, and it can be changed by the following factors:
- Plant community structure and vegetation cover are readily damaged and difficult to recover.
- Humus and/or nutrients, particularly P (Phosphorus), are frequently low in soils.
- Nutrients and organic matter in the soil are frequently depleted and quickly lost.
- There are temperature extremes that are seasonal and/or diurnal.
- Precipitation is usually strong and erosive, and it falls in short bursts.
- There is a chance of strong winds and bushfires, among other things.
Scientists conducted two long-term trials in a Mediterranean habitat, demonstrating that inoculation with indigenous soil microorganisms improved not only plant establishment but also soil fertility and environmental quality [20]. Soil nitrogen (N) content, organic matter, and hydrostable soil aggregates have all grown as a result of this symbiosis, as has nutrient transfer among plant species associated with natural succession. The introduction of target indigenous plant species in conjunction with a managed community of microbial systems was found to be a successful approach for assisting the recovery of low vegetative habitats. The diversity of soil microbe species relevant to the setting of the current project is first discussed in this chapter, followed by a review of the proposed mechanisms of soil structure formation [21]. Microbial symbioses provide a biofertilizing and environmental stability role because colonized plants are better able to receive nutrients and tolerate environmental challenges. In the ecosystem, colonized plants’ greater uptake of soil minerals means that fertilizer treatments can be reduced significantly while crop yields remain comparable or even higher.
Ecological Impact of Potential Soil Microorganisms
Plants have evolved to include a diverse range of microorganisms that play critical functions in plant growth and health. A substantial quantity of data on the structure and dynamics of plant microbiota, as well as the functional capacities of isolated community members, is currently available (Figure 1). There is a pressing need to put microbial innovations into practice, both because of the intriguing functional potential of plant microbiota and because of existing crop production issues [22]. Plants actively recruit microorganisms from the soil/rhizosphere, the phyllosphere, the anthosphere (the external environment of flowers), the spermosphere (the exterior of germinated seed), and the carposphere (the external environment of fruit) [23]. The majority of root microbiota is horizontally transported, meaning it comes from the soil environment, which comprises a varied range of microorganisms dominated by Acidobacteria, Verrucomicrobia, Bacteroidetes, Proteobacteria, Planctomycetes, and Actinobacteria. Bacteria can also be spread vertically via seeds. Seeds are also a major source of microorganisms, which grow in the developing plant’s roots [24, 25]. Plants provide unique ecological niches for soil microbes, which invade the rhizosphere, roots, and to a degree above ground sections [26]. Similarly, Kawasaki, et al. [27] reported that the Brachypodium distachyon (a model for wheat) rhizosphere was dominated by Burkholderiales, Sphingobacteriales and Xanthomonadales, while the bulk soil was dominated by the order Bacillales. Organic acids, amino acids, fatty acids, phenolics, plant growth regulators, nucleotides, sugars, putrescine, sterols, and vitamins, as well as other root exudates, have been shown to impact microbial composition near roots [28].
However, above-ground plant tissues such as vegetative foliar parts, leaves, and floral parts provide distinct settings for endophyte and epiphyte diversities, although the ecology of endosphere and phyllosphere bacteria differs significantly [29]. As a result, different microorganisms are found in the endosphere and phyllosphere at the genus and species level, such as Pseudomonas, Sphingomonas, Frigoribacterium, Curtobacterium, Bacillus, Enterobacter, Acinetobacter, Erwinia, Citrobacter, Pantoea, and Methylobacterium as predominant genera [30, 31]. Wallace, et al. [32] investigated the maize leaf microbiome across 300 different maize lines and discovered that sphingomonads and methylobacteria were the most common species. They also discovered that environmental conditions influenced the phyllosphere’s microbial composition. Pseudomonas and Enterobacteriaceae were found to be the most common taxa in apple flowers, according to Steven, et al. [33]. Pseudomonas was also reported to be the most prevalent genus in various investigations on apple, almond, grapefruit, tobacco, and pumpkin blossoms [34]. Seed-associated bacteria have only lately been studied, with Proteobacteria, Actinobacteria, Bacteroidetes, and Firmicutes being the most common [35, 36]. Seed microbiotas have a connection to soil microbiota, as well as those of flowers and fruits [37, 38].
Functions and Factors Affecting Microorganisms
The term “core plant microbiome” refers to microorganisms that are intimately connected with a particular plant species or genotype regardless of soil or environmental circumstances [39]. A variety of biotic and abiotic variables determine the microbial composition of any plant organ. External environmental conditions such as climate, pathogen presence, and human practices [40] influence microbiota of above- and below-ground plant parts, whereas soil pH, salinity, soil type, soil structure, soil moisture, and soil organic matter and exudates [41] are most relevant for belowground plant parts (Table 1). Beneficial, neutral, and harmful microbes make up the plant microbiome. Plant growth-promoting bacteria (PGPB) can enhance plant growth in a number of ways, both directly and indirectly. Auxin, cytokinin, and gibberellin are phytohormones produced by certain PGPB that affect plant growth by modifying endogenous hormone levels in interaction with a plant. Furthermore, some PGPB can secrete an enzyme called 1-aminocyclopro pane-1-carboxylate (ACC) deaminase, which lowers ethylene levels in the plant. Pseudomonas spp., Arthrobacter spp., Bacillus spp., and other bacteria have been found to boost plant development by producing ACC deaminase [42] (Table 1).
Phytotoxic chemicals, proteins, and phytohormones produced by some bacteria can induce illness symptoms. Pseudomonas syringae, for example, is a well-known plant pathogen with a wide host range that includes tomato, tobacco, olive, and green bean. Erwinia amylovora is another well- known pathogenic bacterium that causes fire blight in fruit trees and ornamental plants. Many significant crop diseases, such as potato and banana, are linked to Xanthomonas species, Ralstonia solanacearum, and Xylella fastidiosa [43]. In terms of above- and below-ground ecological diversity in a terrestrial ecosystem, global temperature change can have an impact on plant ecology. The terrestrial plant community affects the microbial ecology of the soil, which can lead to changes in ecosystem function. Plant species cohabitation can be governed by resource partitioning and sharing of soil resources. Root symbionts have been discovered to improve nutrient uptake efficiency and let the host to persist in low- nutrient environments, thereby contributing directly to the competitive exclusion of other plants [44].
Soil is a complex ecosystem that hosts bacteria, fungus, protists, and animals, despite the fact that plant physiologists sometimes see it as only a source of nutrients for plants [45]. Plants engage in a wide range of interactions with these soil- dwelling creatures, spanning the entire ecological spectrum (competitive, exploitative, neutral, commensal, mutualistic) (Figure 1). Most interaction investigations in modern plant science have focused on reducing pathogenic impacts such herbivory and infection [46] or reducing abiotic stress conditions [47, 48].
| Sl. No. | Organisms involved | Type of interaction | Compounds/ mechanisms involved | Findings | References |
|---|---|---|---|---|---|
| 1 | Moniliophthora roreri and Trichoderma harzianum | Phytopathogen– endophyte | T39 butenolide, harzianolide, sorbicillinol | Compounds dependent on the phytopathogen presence and were spatially localized in the interaction zone. | [49] |
| 2 | Trichoderma atroviride and Arabidopsis sp. | Endophyte– plant | Indole acetic acid- related indoles | Plant root colonization promotes growth and enhances systemic disease resistance in the plant by endophytes. | [50] |
| 3 | Xylella fastidiosa and Methylobacterium mesophilicum | Phytopathogen– endophyte | Hydroxamate type | Genes related to energy production, stress, transport, and motility were upregulated in the phytopathogen, but genes related to growth were downregulated. | [51] |
| 4 | Burkholderia gladioli, B. seminalis, and orchid | Phytopathogen– endophyte– plant | Extracellular polysaccharides; altering hormone metabolism | By using extracellular polysaccharides and by altering hormone metabolism, the endophyte strain probably interacts with the plant, as was suggested by genomic analysis. | [52] |
| 5 | Bradyrhizobium diazoefficiens and Aeschynomene afraspera | Symbiont–plant | C35 hopanoids | C35 hopanoids are essential for symbiosis and are related to evasion of plant defense, utilization of host photosynthates, and nitrogen fixation. | [53] |
| 6 | Stachybotrys elegans and Rhizoctonia solani | Mycoparasite– host | Trichothecenes and atranones | Mycoparasite-induced alterations in Rhizoctonia solani metabolism, growth, and development by the production of mycotoxins. The biosynthesis of many antimicrobial compounds by R. solani was downregulated. | [54] |
| 7 | Candida albicans and Pseudomonas aeruginosa | Microbial community | Quorum sensing | The Pseudomonas aeruginosa QS system may block the yeast-to-hypha transition or activate the hypha-to-yeast reversion of Candida albicans. Farnesol produced by C. albicans downregulates the QS system of P. aeruginosa. | [55] |
| 8 | Vibrio fischeri and fishes or squids | Symbiont–fish | Quorum sensing | In symbiotic association with fishes and squids, the auto-inducer molecule reaches a threshold and luminescence genes are activated. | [56] |
| 9 | Rhizobium leguminosarum and plants | Symbiont–fish | Quorum sensing | The quorum-sensing system in these bacteria is related to different functions: nodulation efficiency, growth inhibition, nitrogen fixation, and plasmid transfer. | [57] |
| 10 | Xanthomonas or Xylella and grapevines or citrus | Pathogen–host | Quorum sensing | Quorum-sensing signaling molecules control the expression of virulence factor as well a s biofilm formation. | [42] |
| 11 | Pantoea stewartii and Zea mays | Pathogen–host | Quorum sensing | Quorum-sensing mutants of Pantoea stewartii were not able to disperse and migrate in the vasculature, consequently decreasing the disease. | [58] |
| 12 | Pseudomonas syringae and tabacco and bean | Phytopathogen– plant | Quorum sensing | Quorum-sensing system allows this bacterium to control motility and exopolysaccharide synthesis essential on biofilm formation and leaves colonization. | [59] |
| 13 | Streptomyces coelicolor and other Actinomycetes spp. | Microbial community | Prodiginines, ctinorhodins, coelichelins, acyl- desferrioxamines, and other compounds | The 227 compounds differentially produced in the interactions were unique. | [60] |
| 14 | Aspergillus nidulans and Streptomyces rapamycinicus | Microbial community | Aromatic polyketides | Activation of fungal secondary metabolite genes that were otherwise silent led physical interaction between the microorganisms. The actinomycete triggered alterations in fungal histone acetylation. | [61] |
| 15 | Pseudomonas sp. | Microbial community | Pyoverdines (siderophore) | Pyoverdines act as signaling molecules, activating a cascade that results in the production of several virulence factors. It is essential to infection and biofilm formation. | [62] |
| 16 | Burkholderia sp., Rhizopus sp., and rice | Symbiont phytopathogen plant | Rhizoxin, bongkrekic acid, enacyloxins | In the absence of the endosymbiont the fungus does not form spores. The phytotoxin rhizoxin is the causal agent of rice seedling blight produced by the endosymbiont; fungus induces the growth of the endosymbiont. | [63] |
| 17 | Vibrio sp. and diverse marine bacteria strains | Microbial community | N,N-bis-(2,3- Dihydroxybenzoyl) -Oserylserine:exogenous siderophore | Siderophores and iron-regulated outer membrane proteins produced by marine bacteria and other species only in the presence of exogenous siderophores. | [64] |
Table 1: Recent Studies of Microbial Interaction.
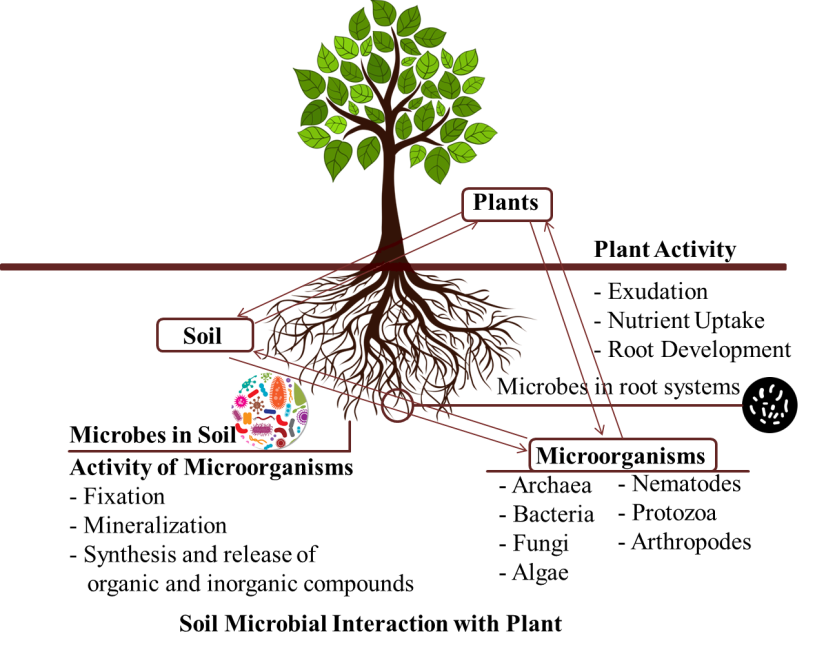
Soil Improvement for the Plant with Microorganisms
To be genuinely effective in an agricultural setting, proposed growth-promoting strains must be shown to be able to be re-inoculated onto plants, colonize the rhizospheric niche successfully, and then mediate nutrient mobilization that improves plant development. Plant–microbe interaction assays can be used to examine prospective strains for their capacity to stimulate plant growth and nutrient absorption (Figure 1) [65]. Plant growth stimulation by microbial mobilization of alternative nitrogen sources has also been reported, as evidenced by greater yield in plants treated with bacterial strains [66, 67]. Sustainable agriculture has grown in popularity in recent years, necessitating new technology advancements to limit the usage of environmentally hazardous chemical fertilizers and pesticides. Several research have focused on plant probiotics as an alternative soil fertilization source; their application in agriculture enhances nutrient availability and conserves field management while causing no harmful consequences [68] (Table 2).
Many biogeochemical processes are mediated by soil microflora, with some species influencing organic matter and soil pollutant biodegradation (rhizoremediators) as well as abiotic stress tolerance [69, 70]. Environmental pollution levels have continuously increased in tandem with industrial growth. Efforts to restore polluted soil have been made, however traditional hazardous chemical treatment approaches are both excessively expensive and damaging to soil microbiota [71]. Furthermore, Cadmium, chromium, copper, lead, mercury, nickel, zinc, and other heavy metals are the most common inorganic pollutants [72]. To rehabilitate polluted soil, however, an environmentally beneficial and low-cost method has been used. Plant species capable of collecting heavy metals are used in phytoremediation for area recovery. The plant’s ability to tolerate huge concentrations of pollutants without compromising its biomass is a limiting factor for this technique’s success [73].
| Sl. No. | Microbial Agents | Nutrient Cycling | Soil Structure |
| 1 | Microflora (fungi, bacteria, actinomycetes) | Catabolize organic matter; mineralize and immobilize nutrients. | Produce organic compounds that bind aggregates; hyphae entangle particles onto aggregates. |
| 2 | Microfauna (Acarina, Collembola) | Regulate bacterial and fungal populations; alter nutrient turnover. | May affect aggregate structure through interactions with microflora |
| 3 | Mesofauna (Acarina, Collembola, enchytraeids) | Regulate fungal and microfaunal populations; alter nutrient turnover; fragment plant residues. | Produce fecal pellets; create biopores; promote humification. |
| 4 | Macrofauna (isopods, centipedes, millipedes, earthworms, etc.) | Fragment plant residues; stimulate microbial activity. | Mix organic and mineral particles; redistribute organic matter and micro- organisms; create biopores; promote humification; produce fecal pellets. |
Table 2: Influences of soil biota on soil processes in ecosystems [70].
Also Metal-tolerant microorganisms, particularly PGPR, have been examined for their ability to bioaccumulate metals in contaminated settings, which improves metal uptake and stimulates plant development. The major mechanism of bioremediation is microbe-plant association, which leads to soil purification and plant development [74].
Future Development of Application in Plant Microbe Interaction
Understanding the connections between plants and microbes, as well as microbes and microbes, will be useful in the future as a regulating microbiome for reducing disease incidence and increasing gross plant yield. In the future, a well-studied plant-microbe alliance could assist enhance crop output at a low cost, potentially leading to another “Green Revolution.” Since people began to farm and were no longer nomads, plants have been an important element of our diet. Since then, the globe has been faced with the constant issue of feeding an ever-increasing population. Microbes’ role in plant-microbe interactions has been extensively researched during the last decade [75, 76]. Extensive research suggests that utilizing beneficial bacteria is a superior long-term strategy for increasing crop productivity, which will play a key role in disease transmission and management [77]. Transcriptomics, proteomics, and metabolomics, as well as bioinformatics, have all been used extensively in plant abiotic stress research [78]. The proteomics approach has been widely used to analyze plant protein profiles in response to abiotic challenges, which could lead to the development of new stress tolerance strategies [79]. Another technique for studying the collection of metabolites found in microbial communities is microbial metabolomics [80]. Infections and their emergence have posed a serious danger to food security, agricultural practices, and the conservation of food species, and understanding the emergence of new pathogens and their significance has become a key job [81].
Conclusion
Plant microbiota and their interactions are extremely complex, and a variety of factors influence how communities form and function. Plant-associated microbes have a big impact on the health and performance of their hosts. Attempts to use beneficial bacteria in the field, on the other hand, have failed to reliably boost crops. Plant-symbiotic microbial community interactions, ecological repercussions of plant- associated microorganisms, and plant-microbial metabolic dynamics are all poorly understood at the moment. More light will be shed on the intricacy of the metabolic potentials of soil microbial communities and their value to the soil ecosystem as DNA-sequencing tools evolve and become more accessible to various working groups. The introduction of genomic techniques has greatly aided our understanding of plant-bacterial interactions, yet genomic approaches are still insufficient to fully explain plant-pathogen interactions. The effect of these alterations on soil biology is still unknown and needs to be investigated further. The fundamental question is still how the impact on taxonomic groups relates to implications on soil microbial communities’ functional capacities.
Moreover, Plants rely on the metabolic activities of soil microbes to gain access to refractory soil-borne nutrients. Given the environmental harm caused by existing fertilization processes, optimizing plant–microbe nutritional interactions is a current research goal for more sustainable agriculture systems. The mechanisms driving the formation of the plant microbiome and its modification in response to plant nutritional status, on the other hand, are exceedingly complicated and difficult to predict. This understanding could help to guide the extremely promising techniques to using microorganisms for more sustainable plant nutrition in a mechanistic way. However, there is still a huge gap in understanding when it comes to a sound theory of plant– microbe interactions.
Acknowledgements
We would like to express our gratitude to the management of MEDWIN PUBLISHERS for inviting us to write the Mini Review and providing the support for the publications.
Conflict of Interest
The authors declare no conflict of interest.
References
-
Kennedy IR, Islam N (2001) The Current and Potential Contribution of Asymbiotic Nitrogen Fixation to Nitrogen Requirements on Farms: A Review. Australian Journal of Experimental Agriculture 41(3): 447-457.
-
Spaepen S, Vanderleyden J, Remans R (2007) Indole- 3-Acetic Acid in Microbial and Microorganism-Plant Signalling. FEMS Microbiol Rev 31(4): 425-448.
-
Glick BR, Todorovic B, Czarny J, Cheng Z, Duan J, et al. (2007) Promotion of Plant Growth by Bacterial ACC Deaminase. Critical Reviews in Plant Sciences 26(5-6): 227-242.
-
Mohiddin FA, Khan MR, Khan SM (2010) Why Trichoderma is Considered Super Hero (Super Fungus) Against the Evil Parasites? Plant Pathology Journal 9(3): 1-11.
-
Peighami-Ashnaei S, Sharifi-Tehrani A, Ahmadzadeh M, Behboudi K (2009) Interaction of Different Media on Production and Biocontrol Efficacy of Pseudomonas Fluorescens P-35 and Bacillus Subtilis B-3 Against Grey Mould of Apple. Journal of Plant Pathology 91(1): 65-70.
-
Dawar S, Wahab S, Tariq M, Zaki MJ (2010) Application of Bacillus Species in the Control of Root Rot Diseases of Crop Plants. Archives Phytopathology Plant Protection 43(4): 412-418.
-
Mendes R, Garbeva P, Raaijmakers JM (2013) The Rhizosphere Microbiome: Significance of Plant Beneficial, Plant Pathogenic and Human Pathogenic Microorganisms. FEMS Microbiol Rev 37(5): 634-663.
-
Oldroyd GE (2013) Speak, Friend and Enter: Signalling Systems that Promote Beneficial Symbiotic Associations in Plants. Nat Rev Microbiol 11(4): 252-263.
-
Ciccazzo S, Esposito A, Rolli E, Zerbe S, Daffonchio D, et al. (2014) Different Pioneer Plant Species Select Specific Rhizosphere Bacterial Communities in a High Mountain Environment. Springer Plus 3: 391.
-
Bever JD (2003) Soil Community Feedback and the Coexistence of Competitors: Conceptual Frameworks and Empirical Tests. New Phytol 157(3): 465-473.
-
Bever JD, Westover KM, Antonovics J (1997) Incorporating the Soil Community into Plant Population Dynamics: The Utilityof the Feedback Approach. J Ecol 85(5): 561-573.
-
Schumpp O, Deakin WJ (2010) How Inefficient Rhizobia Prolong their Existence within Nodules. Trends Plant Sci 15(4): 189-195.
-
Lerner A, Herschkovitz Y, Baudoin E, Nazaret S, Moenne- Loccoz Y, et al. (2006) Effect of Azospirillum Brasilense Inoculation on Rhizobacterial Communities Analyzed by Denaturing Gradient Gel Electrophoresis and Automated Ribosomal Intergenic Spacer Analysis. Soil Biology and Biochemistry 38(6): 1212-1218.
-
Trabelsi D, Ben Ammar H, Mengoni A, Mhamdi R (2012) Appraisal of the Crop-Rotation Effect of Rhizobial Inoculation on Potato Cropping Systems in Relation to Soil Bacterial Communities. Soil Biology and Biochemistry 54: 1-6.
-
Babi´c KH, Schauss K, Hai B (2008) Influence of Different Sinorhizobium Meliloti Inocula on Abundance of Genes Involved in Nitrogen Transformations in the Rhizosphere of Alfalfa (Medicago Sativa L.). Environ Microbiol 10(11): 2922-2930.
-
Naiman AD, Latr´onico A, Garc´ıa de Salamone IE (2009) Inoculation of Wheat with Azospirillum Brasilense and Pseudomonas Fluorescens: Impact on the Production and Culturable Rhizosphere Microflora. European Journal of Soil Biology 45(1): 44-51.
-
De Salamone IEG, Di Salvo LP, Ortega JSE, Sorte PMFB, Urquiaga S, et al. (2010) Field Response of Rice Paddy Crop to Azospirillum Inoculation: Physiology of Rhizosphere Bacterial Communities and the Genetic Diversity of Endophytic Bacteria in Different Parts of the Plants. Plant and Soil 336: 351-362.
-
Khalvati M, Dincer I (2013) Environmental Impact of Soil Microorganisms on Global Change. Springer Science pp: 233-250.
-
Khalvati M, Bartha B, Dupigny A, Schroder P (2010) Arbuscular Mycorrhizal Association is Beneficial for Growth and Detoxification of Xenobiotics of Barley Under Drought Stress. Journal of Soils and Sediments 10: 54-64.
-
Requena N, Perez Solis E, Azcon Aguilar C, Jeffries P, Barea JM (2001) Management of Indigenous Plant-Microbe Symbioses Aids Restoration of Desertified Ecosystems. Appl Environ Microbol 67(2): 495-498.
-
Rillig MC (2004) Arbuscular mycorrhizae, glomalin and soil aggregation quality. Can J Soil Sci 84: 355-363.
-
Compant S, Samad A, Faist H, Sessitsch A (2019) A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. Journal of Advanced Research 19: 29-37.
-
Hardoim PR, van Overbeek LS, Berg G, Pirttilä AM, Compant S, et al. (2015) The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Mol Biol Rev 7993): 293-320.
-
Liu Y, Zuo S, Xu L, Zou Y, Song W (2012) Study on diversity of endophytic bacterial communities in seeds of hybrid maize and their parental lines. Arch Microbiol 194(12): 1001-1012.
-
Hardoim PR, Hardoim CC, van Overbeek LS, van Elsas JD (2012) Dynamics of seedborne rice endophytes on early plant growth stages. PLoS ONE 7(2): e30438.
-
Hartmann A, Schmid M, van Tuinen D, Berg G (2009) Plant-driven selection of microbes. Plant and Soil 321(1- 2): 235-257.
-
Kawasaki A, Donn S, Ryan PR, Mathesius U, Devilla R, et al. (2016) Microbiome and exudates of the root and rhizosphere of Brachypodium distachyon, a model for wheat. PLoS ONE 11(10): e0164533.
-
Hartmann A, Rothballer M, Schmid M, Hiltner L (2008) A pioneer in rhizosphere microbial ecology and soil bacteriology research. Plant and Soil 312(1-2): 7-14.
-
Compant S, Mitter B, Colli Mull JG, Gangl H, Sessitsch A (2011) Endophytes of grapevine flowers, berries, and seeds: identification of cultivable bacteria, comparison with other plant parts, and visualization of niches of colonization. Microb Ecol 62(1): 188-197.
-
Zarraonaindia I, Owens SM, Weisenhorn P, West K, Hampton-Marcell J, et al. (2015) The soil microbiome influences grapevine-associated microbiota. mBio 6(2): e02527-e2614.
-
Kecskeméti E, Berkelmann Löhnertz B, Reineke A, Cantu D (2016) Are epiphytic microbial communities in the carposphere of ripening grape clusters (Vitis vinifera L.) different between conventional, organic, and biodynamic grapes? PLoS ONE 11(18): e0160852.
-
Wallace J, Kremling KA, Kovar LL, Buckler ES (2018) Quantitative genetics of the maize leaf microbiome. Phytobiomes Journal 2(4): 208-224.
-
Steven B, Huntley RB, Zeng Q (2018) The influence of flower anatomy and apple cultivar on the apple flower phytobiome. Phytobiomes 2(3): 171-179.
-
Aleklett K, Hart M, Shade A (2014) The microbial ecology of flowers: an emerging frontier in phyllosphere research. Botany 92: 253-266.
-
Barret M, Briand M, Bonneau S, Préveaux A, Valière S, et al. (2015) Emergence shapes the structure of the seed microbiota. Appl Environ Microbiol 81(4): 1257-1266.
-
Johnston-Monje D, Raizada MN (2011) Conservation and diversity of seed associated endophytes in Zea across boundaries of evolution, ethnography and ecology. PLoS ONE 6(6): e20396.
-
Glassner H, Zchori-Fein E, Yaron S, Sessitsch A, Sauer U, et al. (2018) Bacterial niches inside seeds of Cucumis melo L. Plant and Soil 422(1-2): 101-113.
-
Mitter B, Pfaffenbichler N, Flavell R, Compant S, Antonielli L, et al. (2017) A new approach to modify plant microbiomes and traits by introducing beneficial bacteria at flowering into progeny seeds. Front Microbiol 8(11): 1-10.
-
Toju H, Peay KG, Yamamichi M, Narisawa K, Hiruma K, et al. (2018) Core microbiomes for sustainable agroecosystems. Nature Plants 4: 247-257.
-
Fierer N (2017) Embracing the unknown: disentangling the complexities of the soil microbiome. Nature Reviews Microbiology 15: 579-590.
-
Ladygina N, Hedlund K (2010) Plant species influence microbial diversity and carbon allocation in the rhizosphere. Soil Biol Biochem 42(2): 162-168.
-
Mansfield J, Genin S, Magori S, Citovsky V, Sriariyanum M, et al. (2012) Top 10 plant pathogenic bacteria in molecular plant pathology. Mol Plant Pathol 13(6): 614- 629.
-
Braga RM, Dourado MN, Araújo WL (2016) Microbial interactions: ecology in a molecular perspective. Braz J Microbiol 47(S1): 86-98.
-
Salas-Marina MA, Silva-Flores MA, Uresti-Rivera EE, Castro-Longoria E, Herrera-Estrella A, et al. (2011) Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acid/ethylene and salicylic acid pathways. Eur J Plant Pathol 131: 15-26.
-
Lacava PT, Araújo WL, Marcon J, Maccheroni W, Azevedo JL (2004) Interaction between endophytic bacteria from citrus plants and the phytopathogenic bacteria Xylella fastidiosa, causal agent of citrus-variegated chlorosis. Lett Appl Microbiol 39(1): 55-59.
-
Araújo WL, Creason AL, Mano ET, Camargo-Neves AA, Minami SN, Chang JH, Loper JE (2016) Genome sequencing and transposon mutagenesis of Burkholderia seminalis TC3.4.2 R3 identify genes contributing to suppression of orchid necrosis caused by B. gladioli. Mol Plant Microbe Interact 29(6): 435-446.
-
Barrière Q, Guefrachi I, Gully D, Lamouche F, Pierre O, et al. (2017) Integrated roles of BclA and DD carboxypeptidase 1 in Bradyrhizobium differentiation within NCR-producing and NCR-lacking root nodules. Sci Rep 7: 1-13.
-
Chamoun R, Aliferis KA, Jabaji S (2015) Identification of signatory secondary metabolites during mycoparasitism of Rhizoctonia solani by Stachybotrys elegans. Front Microbiol 6: 1-11.
-
Polke M, Sprenger M, Scherlach K, Albán-Proaño MC, Martin R, et al. (2017) A functional link between hyphal maintenance and quorum sensing in Candida albicans. Mol Microbiol 103(4): 595-617.
-
Fuqua WC, Winans SC, Greenberg EP (1994) Quorum sensing in bacteria: the LuxR-LuxI family of cell density- responsive transcriptional regulators. J Bacteriol 176(2): 269-275.
-
Gonzalez JE, Marketon MM (2003) Quorum sensing in nitrogen-fixing rhizobia. Microbiol Mol Biol Rev 67(4): 574-592.
-
Koutsoudis MD, Tsaltas D, Minogue TD, von Bodman SB (2006) Quorum-sensing regulation governs bacterial adhesion, biofilm development, and host colonization in Pantoea stewartii subspecies stewartii. Proc Natl Acad Sci USA 103(15): 5983-5988.
-
Quiñones B, Dulla G, Lindow SE (2005) Quorum sensing regulates exopolysaccharide production, motility, and virulence in Pseudomonas syringae. Mol Interact 18(7): 682-693.
-
Bentley SD, Chater KF, Cerdeño-Tárraga AM, Challis GL, Thomson NR, et al. (2002) Complete genome sequence of the model actinomycete Streptomyces coelicolor A3 (2). Nature 417: 141-147.
-
Bertrand S, Bohni N, Schnee S, Schumpp O, Gindro K, et al. (2014) Metabolite induction via microorganism co- culture: a potential way to enhance chemical diversity for drug discovery. Biotechnol Adv 32(6): 1180-1204.
-
Jimenez PN, Koch G, Thompson JA, Xavier KB, Cool RH, et al. (2012) The multiple signaling systems regulating virulence in Pseudomonas aeruginosa. Microbiol Mol Biol Rev 76(1): 46-65.
-
Depoorter E, Bull MJ, Peeters C, Coenye T, Vandamme P, et al. (2016) Burkholderia: an update on taxonomy and biotechnological potential as antibiotic producers. Appl Microbiol Biotechnol 100(12): 5215-5229.
-
Kanoh K, Kamino K (2001) Effect of exogenous siderophores on iron uptake activity of marine bacteria under iron-limited conditions. Appl Environ Microl 67(4): 1710-1717.
-
Rudrappa T, Czymmek KJ, Paré PW, Bais HP (2008) Root- secreted malic acid recruits beneficial soil bacteria. Plant Physiol 148(3): 1547-1556.
-
Bonkowski M, Villenave C, Griffiths B (2009) Rhizosphere fauna: the functional and structural diversity of intimate interactions of soil fauna with plant roots. Plant Soil 321: 213-233.
-
Zhang Y, Lubberstedt T, Xu ML (2013) The genetic and molecular basis of plant resistance to pathogens. J Genet Genomics 40(1): 23-35.
-
Muller DB, Vogel C, Bai Y, Vorholt JA (2016) The plant microbiota: systems-level insights and perspectives. Annu Rev Genet 50: 211-234.
-
Meena KK, Sorty AM, Bitla UM, Choudhary K, Gupta P, et al. (2017) Abiotic stress responses and microbe- mediated mitigation in plants: the omics strategies. Front Plant Sci 8: 172.
-
Oconnor PJ, Smith SE, Smith FA (2002) Arbuscular mycorrhizas influence plant diversity and community structure in a semiarid herbland. New Phytologist 154(1): 209-218.
-
Kozdroj J, Trevors JT, Elsas JDV (2004) Influence of introduced potential biocontrol agents on maize seedling growth and bacterial community structure in the rhizosphere. Soil Biology and Biochemistry 36(11): 1775-1784.
-
Ahemad M, Kibret M (2014) Mechanisms and applications of plant growth promoting rhizobacteria: current perspective. J. King Saud Univ Sci 26(1): 1-20.
-
Gao G, Yin D, Chen S (2012) Effect of biocontrol agent Pseudomonas fluorescens 2P24 on soil fungal community in cucumber rhizosphere using T-RFLP and DGGE. PLoS ONE 7(2): e31806.
-
Shaharoona B, Naveed M, Arshad M, Zahir ZA (2008) Fertilizer-dependent efficiency of Pseudomonas for improving growth, yield, and nutrient use efficiency of wheat (Triticum aestivum L.). Appl Microbiol Biotechnol 79(1): 147-155.
-
Adesemoye AO, Torbert HA, Kloepper JW (2009) Plant growth promoting rhizobacteria allow reduced application rates of chemical fertilizers. Microb Ecol 58(4): 921-929.
-
Bhardwaj D, Ansari MW, Sahoo K, Tuteja N (2014) Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb Cell Fact 13: 1-10.
-
Rajkumar M, Sandhya S, Prasad MNV, Freitas H (2012) Perspectives of plant-associated microbes in heavy metal phytoremediation. Biotechnol Adv 30(6): 1562- 1574.
-
Zhuang X, Chen J, Shim H (2007) New advances in plant growth-promotion rhizobacteria for bioremediation. Environ Int 33(3): 406-413.
-
Hendrix PF, Crossley DA, Blair JM, Coleman DC (1990) Soil biota as components of sustainable agroecosystems. In: Edwards CA, et al. (Eds.), 1st (Edn.), Sustainable Agricultural Systems. Soil and Water Conservation Society, IA, pp: 637-654.
-
Ma Y, Prasad MNV, Rajkumar M, Freitas H (2010) Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils. Biotechnol Adv 29(2): 248-258.
-
Vorholt JA (2012) Microbial life in the phyllosphere. Nature Reviews Microbiology 10(12):828-840.
-
Bulgarelli D, Schlaeppi K, Spaepen S, Themaat EVLV, Lefert PS (2013) Structure and functions of the bacterial microbiota of plants. Annual Review of Plant Biology 64: 807-838.
-
Reid A (2011) Microbes helping to improve crop productivity. Microbe 6(10): 435.
-
Soda N, Wallace S, Karan R (2015) Omics study for abiotic stress responses in plants. Advances in Plants & Agriculture Research 2(1): 00037.
-
Gupta B, Saha J, Sengupta A, Gupta K (2013) Plant Abiotic Stress: ‘Omics’ Approach. J Plant Biochem Physiol 1: 100e108.
-
Barkal LJ, Theberge AB, Guo CJ, Spraker J, Rappert L, et al. (2016) Microbial metabolomics in open microscale platforms. Nature Communications 7: 10610.
-
Thynne E, McDonald MC, Solomon PS (2015) Phytopathogen emergence in the genomics era. Trends in Plant Science 20(4): 246-255.
- Lessons to Learn: Trees are More than the Lungs of the World
- Community Forestry Enterprises as a Model for Sustainable Forest Development: The Case Of The "Baja Tarahumara" in Chihuahua, Mexico
- Ecological and Socio-Economic Impacts of Chromolaena odorata and Mesosphaerum suaveolens, Two Invasive Alien Species in Central and Southern Benin, West Africa
- Epigenetic Sustainability: Modeling the Human Factor as a Natural Resource through Science 4.0 and the NR3C1 Biological Pilot
- Growth-at-Risk: A Framework for Assessing Economic Vulnerability
- The Rural Territory as a Socioecological System for the Management of Public Policy for Sustainable Rural Development